Monuments Unveiled: Genetic Characterization of Large Old Chestnut (Castanea sativa Mill.) Trees Using Comparative Nuclear and Chloroplast DNA Analysis

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Extraction
2.2. Microsatellite Analysis
2.3. Chloroplast DNA Analysis
2.4. Data Analysis
2.4.1. Nuclar Microsatellite Diversity
2.4.2. Estimation of Nuclear Gene Pools
2.4.3. Chloroplast DNA Sequence Analyses
3. Results
3.1. Genetic Diversity
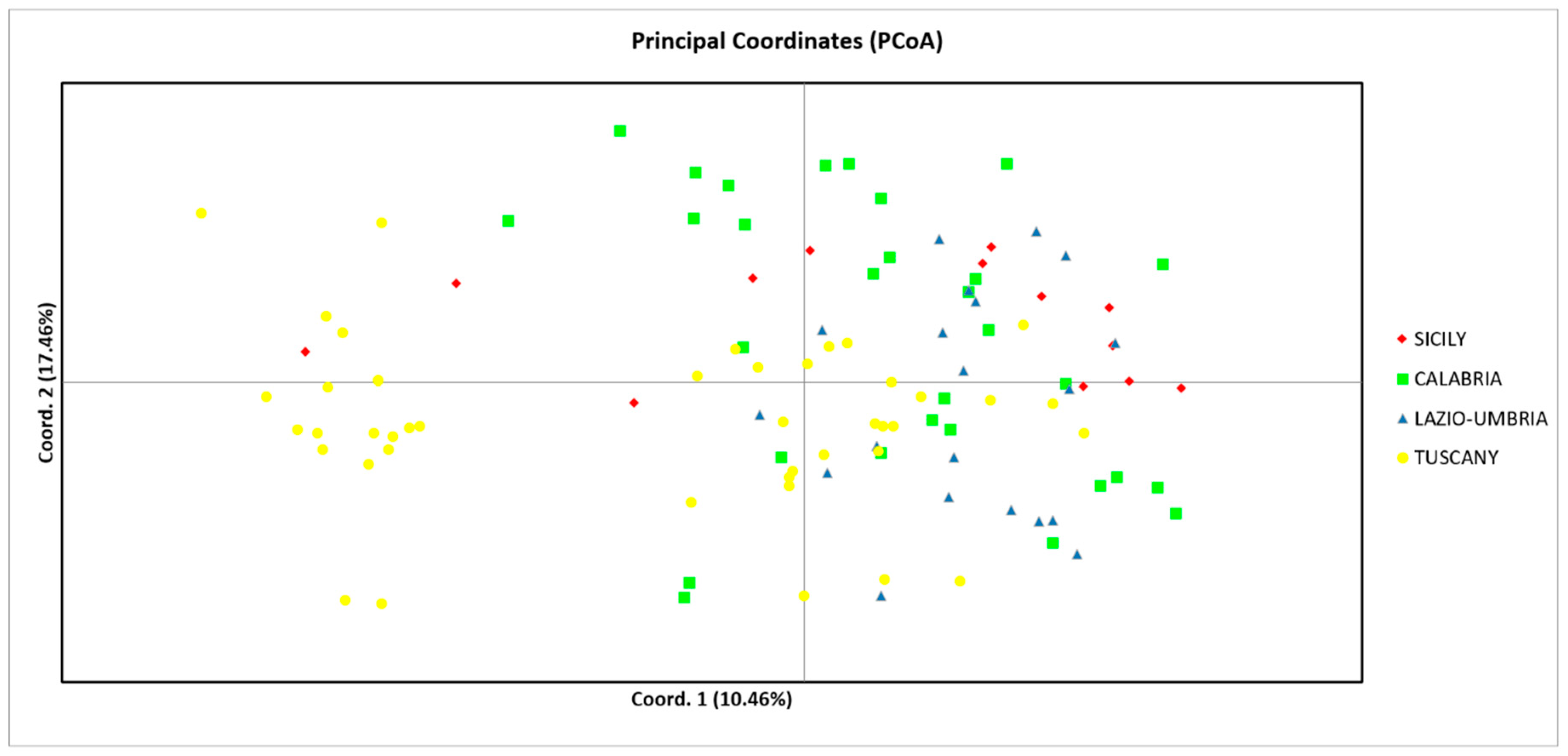
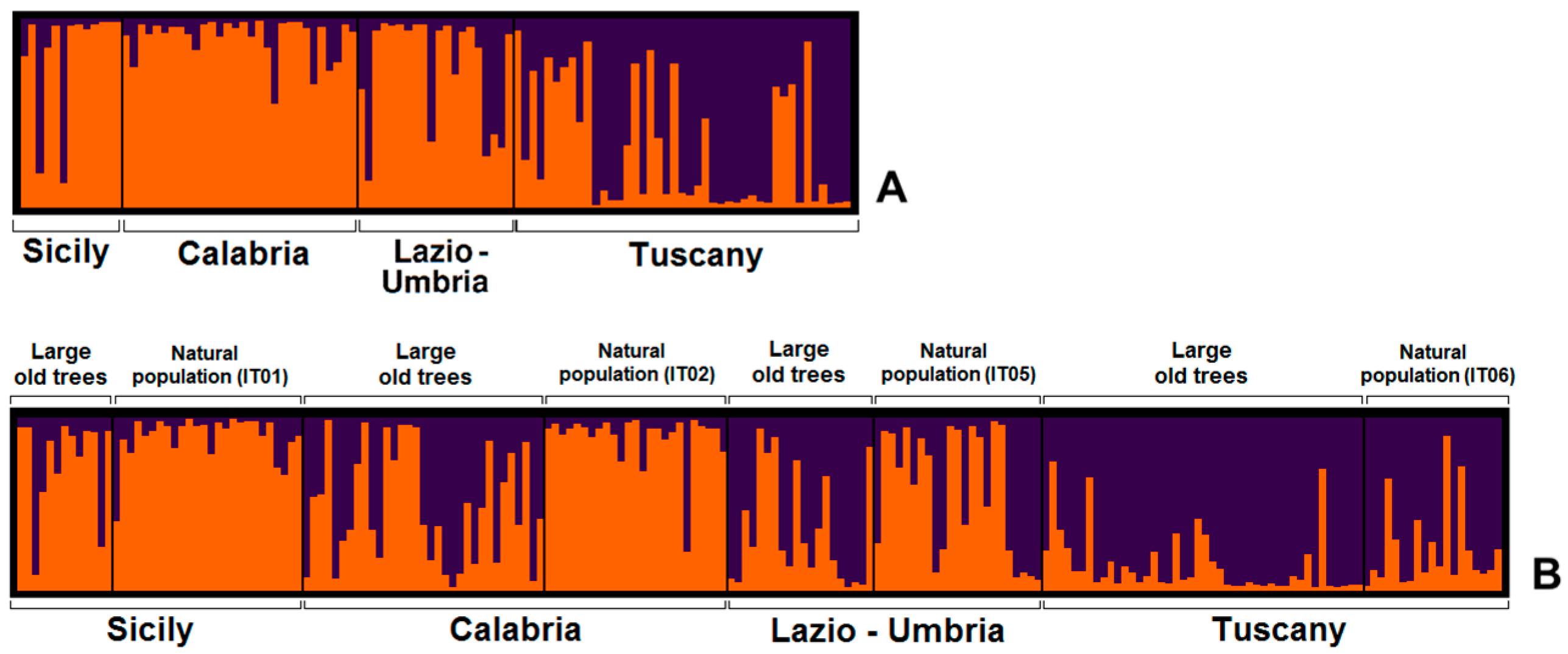
3.2. Nuclear DNA Variation
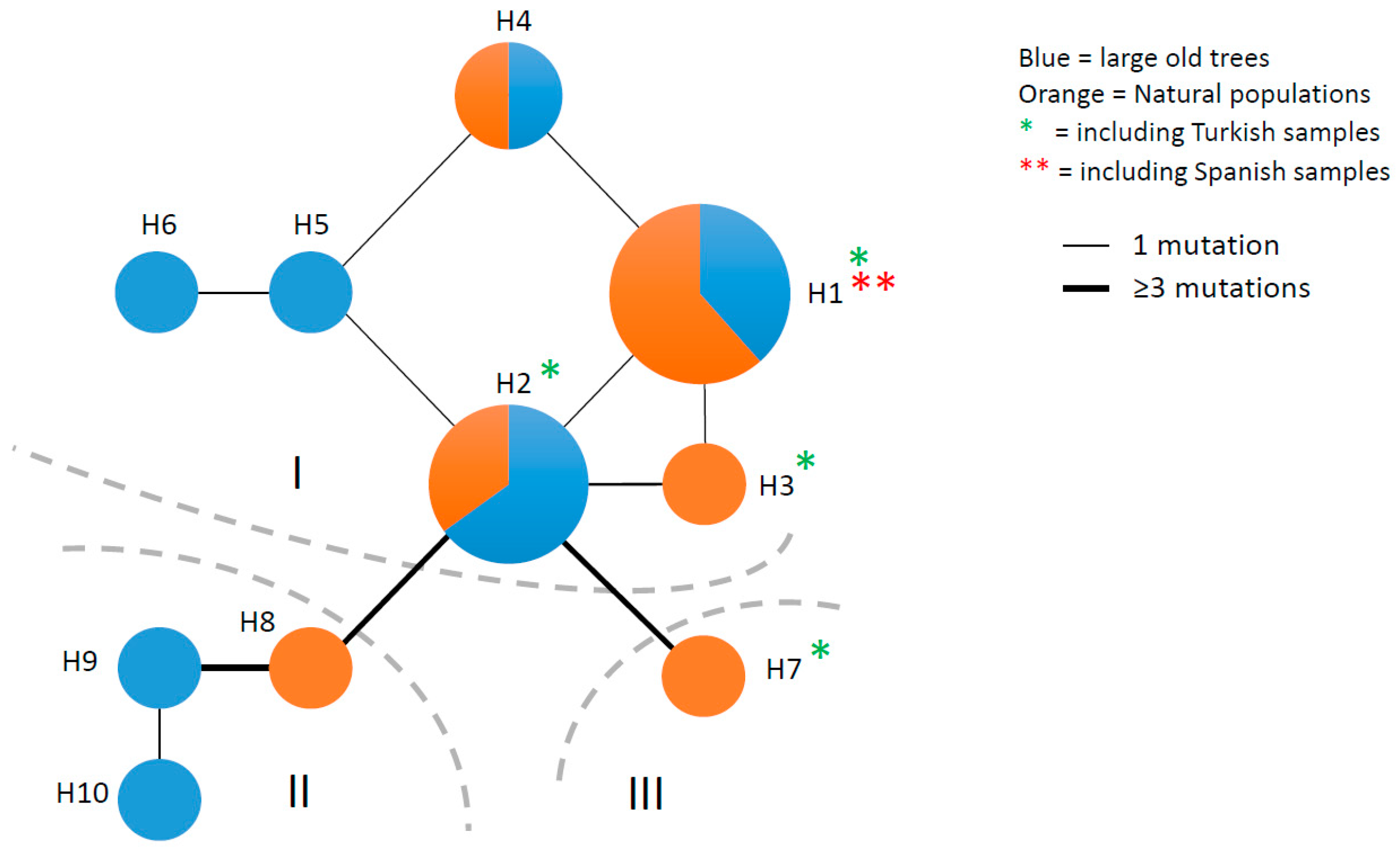
3.3. Chloroplast DNA Variation
4. Discussion
4.1. Central and South Italy as Reservoir of Giant Chestnut Trees
4.2. Genetic Identity of Large Old Trees
4.3. Ancient Phylogeographic Signatures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petit, R.J.; Hampe, A. Some Evolutionary Consequences of Being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.S.; Bell, D.M.; Hersh, M.H.; Nichols, L. Climate change vulnerability of Forest biodiversity: Climate and competition tracking of demographic rates. Glob. Chang. Biol. 2011, 17, 1834–1849. [Google Scholar] [CrossRef]
- Kremer, A.; Potts, B.M.; Delzon, S. Genetic divergence in forest trees: Understanding the consequences of climate change. Funct. Ecol. 2014, 28, 22–36. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Ann. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene Flow and Local Adaptation in Trees. Ann. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Florian, J.A.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninenk, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change—Evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar]
- Pritchard, H.W.; Moat, J.F.; Ferraz, J.B.S.; Marks, T.R.; Camargo, J.L.C.; Nadarajan, J.; Ferraz, I.D.K. Innovative approaches to the preservation of forest trees. For. Ecol. Manag. 2014, 333, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Lindenmayer, D.B.; Laurance, W.F. The Unique Challenges of Conserving Large Old Trees. Trends Ecol. Evol. 2016, 31, 416–418. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F. The ecology, distribution, conservation and management of large old trees. Biol. Rev. 2017, 92, 1434–1458. [Google Scholar] [CrossRef]
- Blicharska, M.; Mikusiński, G. Incorporating social and cultural significance of large old trees in conservation policy. Conserv. Biol. 2014, 28, 1558–1567. [Google Scholar] [CrossRef]
- Lutz, J.A.; Furniss, T.J.; Johnson, D.J.; Davies, S.J.; Allen, D.; Alonso, A.; Anderson-Teixeira, K.J.; Andrade, A.; Baltzer, J.; Becker, K.M.L.; et al. Global importance of large diameter trees. Glob. Ecol. Biogeogr. 2018, 27, 849–864. [Google Scholar] [CrossRef] [Green Version]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.A.; Abarca, D.; et al. Epigenetic regulation of adaptive responses of forest tree species to the environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef]
- Beghè, D.; Ganino, T.; Dall’Asta, C.; Silvanini, A.; Cirlini, M.; Fabbri, A. Identification and characterization of ancient Italian chestnut using nuclear microsatellite markers. Sci. Hortic. 2013, 164, 50–57. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Cannizzaro, S.; Corinto, G.L. The role of monumental trees in defining local identity and in tourism. A case study in the Marches Region. Geoprog. J. (S Humanit. 1) 2014, 1, 29–48. [Google Scholar]
- Ciaffi, M.; Alicandri, E.; Vettraino, A.M.; Paolacci, A.R.; Tamantini, M.; Tomao, A.; Agrimi, M.; Kuzminsky, E. Conservation of veteran trees within historical gardens (COVE): A case study applied to Platanus orientalis L. in central Italy. Urban For. Urban Green. 2018, 34, 336–347. [Google Scholar] [CrossRef]
- Díez, C.; Trujillo, I.; Barrio, E.; Belaj, A.; Diego, B.; Rallo, L. Centennial olive trees as a reservoir of genetic diversity. Ann. Bot. 2011, 108, 797–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barazani, O.; Westberg, E.; Hanin, N.; Dag, A.; Kerem, Z.; Tugendhaft, Y.; Hmidat, M.; Hijawi, T.; Kadereit, J.V. A comparative analysis of genetic variation in rootstocks and scions of old olive trees—A window into the history of olive cultivation practices and past genetic variation. BMC Plant Biol. 2014, 14, 146. [Google Scholar] [CrossRef] [Green Version]
- Anestiadou, K.; Nikoloudakis, N.; Hagidimitriou, M.; Katsiotis, A. Monumental olive trees of Cyprus contributed to the establishment of the contemporary olive germplasm. PLoS ONE 2017, 12, e0187697. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Barreneche, T.; Mattioni, C.; Villani, F.; Díaz-Hernández, B.; Martín, L.M.; Robles-Loma, A.; Cáceres, Y.; Martín, A. Instant domestication process of European chestnut cultivars. Ann. App. Biol. 2018, 174, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Mattioni, C.; Martin, M.A.; Pollegioni, P.; Cherubini, M.; Villani, F. Microsatellite markers reveal a strong geographical structure in European populations of Castanea sativa (Fagaceae): Evidence for multiple glacial refugia. Am. J. Bot. 2013, 100, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Krebs, P.; Pezzatti, G.B.; Beffa, G.; Tinner, W.; Conedera, M. Revising the sweet chestnut (Castanea sativa Mill.) refugia history of the last glacial period with extended pollen and macrofossil evidence. Quat. Sci. Rev. 2019, 206, 111–128. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Jiménez-Alfaro, B.; Chytrý, M.; Díaz-Varela, E.R.; Álvarez-Álvarez, P. Glacial refugia and mid-Holocene expansion delineate the current distribution of Castanea sativa in Europe. Palaeogeogr. Palaeoclimatol. Palaeocl. 2018, 491, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobot. 2004, 13, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Gross, B.L.; Henk, A.D.; Richards, C.M.; Fazio, G.; Volk, G.M. Genetic diversity in Malus X domestica (Rosaceae) through time in response to domestication. Am. J. Bot. 2014, 101, 1770–1779. [Google Scholar] [CrossRef] [PubMed]
- Mellano, M.G.; Beccaro, G.L.; Donno, D.; Torello Marinoni, D.; Boccacci, P.; Canterino, S.; Cerutti, A.K.; Bounous, G. Castanea spp. biodiversity conservation: Collection and characterization of the genetic diversity of an endangered species. Gen. Res. Crop Evol. 2012, 59, 1727–1741. [Google Scholar] [CrossRef]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2010, 177, 309–334. [Google Scholar] [CrossRef]
- Mason, A.S. SSR genotyping. In Plant Genotyping; Methods in Molecular Biology Book Series; Humana Press: New York, NY, USA, 2015; Volume 1245, pp. 77–89. [Google Scholar] [CrossRef]
- Rojaora Om, P.; Mosseler, A. Challenges and opportunities for conservation of forest genetic resources. Euphytica 2001, 118, 197–212. [Google Scholar] [CrossRef]
- Holderegger, R.; Kamm, U.; Gugerli, F. Adaptive vs. neutral genetic diversity: Implications for landscape genetics. Landsc. Ecol. 2006, 21, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Dumolin-Lapegue, S.; Demesure, B.; Fineschi, S.; Le, C.V.; Petit, R.J. Phylogeographic structure of white oaks throughout the European continent. Genetics 1997, 146, 1475–1487. [Google Scholar]
- Simeone, M.C.; Grimm, G.W.; Papini, A.; Vessella, F.; Cardoni, S.; Tordoni, E.; Piredda, R.; Franc, A.; Denk, T. Plastome data reveal multiple geographic origins of Quercus Group Ilex. PeerJ 2016, 4, e1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewingtown, A.; Parker, E. Ancient Trees: Trees that Live for 1000 Years; Collins and Brown Eds: London, UK, 2012; ISBN 9781849940580. [Google Scholar]
- Buck, E.J.; Hadonou, M.; James, C.J.; Blakesley, D.; Russell, K. Isolation and characterization of polymorphic microsatellites in European chestnut (Castanea sativa Mill.). Mol. Ecol. Notes 2003, 3, 239–241. [Google Scholar] [CrossRef]
- Marinoni, D.; Akkak, A.; Bounous, G.; Edwards, K.J.; Botta, R. Development and characterization of microsatellite markers in Castanea sativa (Mill.). Mol. Breed. 2003, 11, 127–136. [Google Scholar] [CrossRef]
- Steinkellner, H.; Lexer, C.; Turetschek, E.; Glössl, J. Conservation of (GA) n microsatellite loci between Quercus species. Molr Ecol. 1997, 6, 1189–1194. [Google Scholar] [CrossRef]
- Shaw, J.; Craddock, J.H.; Binkley, M.A. Phylogeny and Phylogeography of North American Castanea Mill. (Fagaceae) Using cpDNA Suggests Gene Sharing in the Southern Appalachians (Castanea Mill., Fagaceae). Castanea 2012, 77, 186–211. [Google Scholar] [CrossRef]
- Piredda, R.; Simeone, M.C.; Attimonelli, M.; Bellarosa, R.; Schirone, B. Prospects of barcoding the Italian wild dendroflora: Oaks reveal severe limitations to tracking species identity. Mol. Ecol. Resour. 2011, 11, 72–83. [Google Scholar] [CrossRef]
- Chapuis, M.P.; Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Meirmans, P.; Tienderen, P. GENOTYPE and GENODIVE: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2008, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Kalinowski, S.T. HP-RARE 1.0: A computer program for performing rarefaction on measures of allelic richness. Mol. Ecol. Notes 2005, 5, 187–189. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statisticsfor the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [PubMed]
- Exoffier, L.; Laval, G.; Schneider, S. Arlequin version (3.0): An integrated software package for population genetics data analysis. Evol. Bioinfor. 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Wen, W. Documentation for STRUCTURE Software Version 2.3.4; Department of Human Genetics, University of Chicago: Chicago, II, USA, 2009. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Mattioni, C.; Martín, A.; Chiocchini, F.; Cherubini, M.; Gaudet, M.; Pollegioni, P.; Velichkov, I.; Jarman, R.; Chambers, F.; Paule, L.; et al. Landscape genetics structure of European sweet chestnut (Castanea sativa Mill): Indications for conservation priorities. Tree Genet. Genomes 2017, 13, 39–52. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Crandall, K.A. Selecting the Best-Fit Model of Nucleotide Substitution. Syst. Biol. 2011, 50, 580–601. [Google Scholar] [CrossRef]
- Zapponi, L.; Mazza, G.; Farina, A.; Fedrigoli, L.; Mazzocchi, F.; Roversi, P.F.; Peverieri, G.S.; Mason, F. The role of monumental trees for the preservation of saproxylic biodiversity: Re-thinking their management in cultural landscapes. Nat. Conserv. 2017, 19, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Gellini, R.; Grossoni, P. Botanica Forestale; CEDAM: Padova, Italy, 1997; ISBN 8813202601. [Google Scholar]
- Larson, D.W.; Matthes, U.; Gerrath, J.A.; Larson, N.W.K.; Gerrath, J.M.; Nekola, J.C.; Walker, G.L.; Porembski, S.; Charlto, A. Evidence for the widespread occurrence of ancient forests on cliffs. J. Biogeogr. 2001, 27, 319–321. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F.; Baliva, M.; Dinella, A.; Di Fiore, L.; Marchiano, V.; Presutti, S.E.; de Vivo, G.; Schettino, A.; di Filippo, A. Tree growth patterns associated with extreme longevity: Implication for ecology and conservation of primeval trees in Mediterranean mountains. Anthropocene 2019, 26, 100199. [Google Scholar] [CrossRef]
- Douaihy, B.; Saliba, C.; Stefan, J.; Simeoni, M.C.; Cardoni, S.; Frahat, P.; Bou Dagher Kharrat, M. Traking diversity and evolutionary pathways of Lebanese oak taxa through plastome analyses. Bot. Lett. 2020, 167, 315–330. [Google Scholar] [CrossRef]
- Poli, I. Alberi Monumentali Della Garfagnana; Storia e Mitologia; Pacini Fazzi, M., Ed.; Pacini Fazzi: Lucca, Italy, 2014; ISBN 978-88-6550378-2. [Google Scholar]
- Muir, G.; Lowe, A.J.; Fleming, C.C.; Vogl, C. High nuclear genetic diversity, high levels of outcrossing and low differentiation among remnant populations of Quercus petraea at the margin of its range in Ireland. Ann. Bot. 2004, 93, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Dane, F. Comparative chloroplast and nuclear DNA analysis of Castanea species in the southern region of the USA. Tree Genet. Genomes 2013, 9, 107–116. [Google Scholar] [CrossRef]
- Gismondi, A.; di Marco, G.; Delorenzo, M.; Canini, A. Upgrade of Castanea sativa (Mill.) genetic resources by sequencing of barcode markers. J. Genet. 2015, 94, 519–524. [Google Scholar] [CrossRef]
- Lahaye, R.; van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newmaster, S.G.; Fazekas, A.J.; Steeves, R.A.; Janovec, J. Testing candidate plant barcode regions in the Myristicaceae. Mol. Ecol. Resour. 2008, 8, 480–490. [Google Scholar] [CrossRef]
- Fineschi, S.; Taurchini, D.; Villani, F.; Vendramin, G.G. Chloroplast DNA polymorphism reveals little geographical structure in Castanea sativa Mill. (Fagaceae) throughout southern European countries. Mol. Ecol. 2000, 9, 1495–1503. [Google Scholar] [CrossRef]
- Krebs, P.; Conedera, M.; Pradella, M.; Torriani, D.; Felber, M.; Tinner, W. Quaternary refugia of the sweet chestnut (Castanea sativa Mill.): An extended palynological approach. Veg. Hist. Archaeobot. 2004, 13, 145–160. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | Traditional Name * | Region | Geographical Area | Collected Samples | Latitude | Longitude | Trunk Circumference (in Meters) |
|---|---|---|---|---|---|---|---|
| 1 | “Nave” | Sicily | Southern Italy | 3 canopy | 377.544 | 151.311 | 20 |
| 2 | “Cento Cavalli” | Sicily | Southern Italy | 4 canopy | 377.501 | 151.304 | 22 |
| 3 | “Castagno Vespa” | Sicily | Southern Italy | 1 canopy | 376.802 | 150.820 | |
| 4 | Sicily | Southern Italy | 1 canopy | 375.139 | 140.252 | 7.3 | |
| 5 | Sicily | Southern Italy | 1 canopy | 385.139 | 150.260 | 4.7 | |
| 6 | “Castagno Ruossa” | Sicily | Southern Italy | 1 canopy | 373.860 | 150.120 | 5.7 |
| 7 | Sicily | Southern Italy | 1 canopy | 374.438 | 150.489 | 4.0 | |
| 8 | Sicily | Southern Italy | 1 canopy | 374.432 | 150.506 | 3.5 | |
| 9 | Sicily | Southern Italy | 1 root suckers, 1 top canopy | 374.438 | 150.489 | 5.5 | |
| 10 | Sicily | Southern Italy | 1 canopy | 377.625 | 150.921 | 9 | |
| 11 | Sicily | Southern Italy | 1 canopy | 374.574 | 150.921 | 6.10 | |
| 12 | Sicily | Southern Italy | 1 root suckers, 1 canopy | 374.591 | 150.631 | 4.5 | |
| 13 | Sicily | Southern Italy | 1 canopy | 374.521 | 150.656 | 5.7 | |
| 14 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 381.749 | 160.219 | 9.50 | |
| 15 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 381.749 | 160.221 | 5.40 | |
| 16 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 381.741 | 160.219 | 6.90 | |
| 17 | Calabria | Southern Italy | 2 canopy | 381.781 | 160.263 | 6.40 | |
| 18 | Calabria | Southern Italy | 1 root suckers, 2 top canopy | 381.781 | 160.265 | 700 | |
| 19 | Calabria | Southern Italy | 1 root suckers, 2 top canopy | 381.777 | 160.264 | >5 | |
| 20 | Calabria | Southern Italy | 1 root suckers, 1 top canopy | 384.742 | 163.293 | 6.00 | |
| 21 | Calabria | Southern Italy | 1 root suckers, 1 top canopy | 384.738 | 163.285 | 9.80 | |
| 22 | Calabria | Southern Italy | 1 root suckers, 1 top canopy | 384.745 | 163.304 | 6 | |
| 23 | Calabria | Southern Italy | 1 canopy | 384.744 | 163.306 | 5.8 | |
| 24 | Calabria | Southern Italy | 1 canopy | 384.749 | 163.304 | 6.7 | |
| 25 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 390.410 | 167.227 | 9.3 | |
| 26 | Calabria | Southern Italy | 1 canopy | 390.406 | 167.236 | 6.7 | |
| 27 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 390.412 | 167.237 | 8.5 | |
| 28 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 390.402 | 167.224 | 9.1 | |
| 29 | Calabria | Southern Italy | 1 canopy | 390.377 | 167.257 | 7.9 | |
| 30 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 393.051 | 166.569 | 6.5 | |
| 31 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 391.530 | 163.703 | 8.6 | |
| 32 | Calabria | Southern Italy | 1 root sucker, 1 canopy | 391.530 | 163.704 | 8.3 | |
| 33 | Calabria | Southern Italy | 1 root suckers, 1 canopy | 391.529 | 163.702 | 6.2 | |
| 34 | Calabria | Southern Italy | 1 canopy | 391.945 | 161.128 | 6.7 | |
| 35 | Calabria | Southern Italy | 1 root sucker | 397.257 | 159.061 | 8.5 | |
| 36 | Calabria | Southern Italy | 1 root sucker | 397.256 | 159.062 | 13.3 | |
| 37 | Calabria | Southern Italy | 1 root sucker, 1 canopy | 397.254 | 159.062 | 8.7 | |
| 38 | Calabria | Southern Italy | 1 root sucker, 1 canopy | 397.264 | 158.954 | 5.7 | |
| 39 | Latium | Central Italy | 1 root sucker, 1 canopy | 427.035 | 119.181 | >5 | |
| 40 | Umbria | Central Italy | 1 canopy | 425.153 | 120.704 | 7.42 | |
| 41 | Umbria | Central Italy | 1 canopy | 423.826 | 122.122 | 6.50 | |
| 42 | Umbria | Central Italy | 1 canopy | 422.724 | 123.545 | 6.30 | |
| 43 | Umbria | Central Italy | 1 canopy | 422.948 | 124.148 | 8.80 | |
| 44 | Umbria | Central Italy | 1 root sucker, 1 canopy | 422.948 | 124.148 | 7.43 | |
| 45 | Umbria | Central Italy | 1 root sucker, 1 canopy | 422.954 | 12.413 | 7.74 | |
| 46 | Umbria | Central Italy | 1 root sucker, 1 canopy | 424.986 | 126.910 | 7.50 | |
| 47 | Umbria | Central Italy | 1 root sucker, 1 canopy | 422.955 | 124.127 | 8.65 | |
| 48 | Umbria | Central Italy | 1 canopy | 422.905 | 124.152 | 8.45 | |
| 49 | Umbria | Central Italy | 1 canopy | 426.512 | 126.943 | 5.3 | |
| 50 | Umbria | Central Italy | 1 canopy | 426.511 | 126.943 | >5 | |
| 51 | Umbria | Central Italy | 1 canopy | 426.507 | 126.943 | >5 | |
| 52 | Umbria | Central Italy | 1 canopy | 426.533 | 126.943 | >5 | |
| 53 | Umbria | Central Italy | 1 root sucker, 1 canopy | 426.598 | 126.944 | 5.8 | |
| 54 | Umbria | Central Italy | 1 root sucker, 1 canopy | 426.493 | 126.942 | 6.0 | |
| 55 | Umbria | Central Italy | 1 root sucker, 1 canopy | 427.342 | 127.954 | 5.3 | |
| 56 | Umbria | Central Italy | 1 root sucker, 1 canopy | 426.286 | 126.997 | 7.7 | |
| 57 | Umbria | Central Italy | 1 canopy | 426.288 | 126.692 | 8.1 | |
| 58 | Umbria | Central Italy | 1 canopy | 426.291 | 127.010 | 9.2 | |
| 59 | Umbria | Central Italy | 1 canopy | 426.294 | 127.013 | 7.2 | |
| 60 | Umbria | Central Italy | 1 root sucker, 2 canopy | 427.393 | 127.912 | 5.20 | |
| 61 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 430.301 | 110.945 | 6.2 | |
| 62 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 430.276 | 110.949 | 7.0 | |
| 63 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 430.282 | 110.944 | 8.0 | |
| 64 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 431.660 | 109.935 | 7.20 | |
| 65 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 431.664 | 109.934 | 7.8 | |
| 66 | “Miraglia” | Tuscany | Central Italy | 1 root sucker, 1 canopy | 455.715 | 114.415 | 8.5 |
| 67 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 436.007 | 119.722 | 9.6 | |
| 68 | Tuscany | Central Italy | 1 root sucker | 43.600 | 119.714 | 7.0 | |
| 69 | Tuscany | Central Italy | 1 root sucker | 436.012 | 119.718 | 7.0 | |
| 70 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 440.784 | 103.311 | 7.3 | |
| 71 | Tuscany | Central Italy | 1 canopy | 440.791 | 103.314 | 5.50 | |
| 72 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 440.780 | 103.310 | 8.0 | |
| 73 | “Roggio” | Tuscany | Central Italy | 1 root sucker, 1 canopy | 440.222 | 101.809 | 10.0 |
| 74 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 442.230 | 103.500 | 8.40 | |
| 75 | “Prato Maleta” | Tuscany | Central Italy | 1 root sucker, 1 canopy | 442.244 | 103.539 | 9.6 |
| 76 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 442.237 | 103.512 | 6.5 | |
| 77 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 442.240 | 103.509 | 10.0 | |
| 78 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 442.246 | 103.514 | 9.0 | |
| 79 | Tuscany | Central Italy | 2 canopy | 442.232 | 103.491 | 8.8 | |
| 80 | “Prato Fosco” | Tuscany | Central Italy | 1 canopy | 441.748 | 104.712 | 8.2 |
| 81 | “Volpiglione” | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.725 | 104.733 | 8.5 |
| 82 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.727 | 104.731 | 10 | |
| 83 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.773 | 104.670 | 8.2 | |
| 84 | “Marchi” | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.082 | 105.262 | 10.5 |
| 85 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.079 | 105.262 | 8.0 | |
| 86 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.075 | 101.053 | 7.5 | |
| 87 | Tuscany | Central Italy | 1 top canopy | 441.060 | 105.368 | 11.0 | |
| 88 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.061 | 105.365 | 10.0 | |
| 89 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.059 | 105.366 | 9.20 | |
| 90 | Tuscany | Central Italy | 1 root sucker, 1 canopy | 441.055 | 105.369 | 8.20 | |
| 91 | Tuscany | Central Italy | 1 canopy | 441.063 | 105.369 | 9.20 | |
| 92 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.068 | 105.374 | 8.20 | |
| 93 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.077 | 105.374 | 10.0 | |
| 94 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.077 | 105.366 | ||
| 95 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.078 | 105.267 | 9.40 | |
| 96 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.089 | 105.269 | 8.0 | |
| 97 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.094 | 105.270 | 7.20 | |
| 98 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.061 | 105.261 | 7.30 | |
| 99 | Tuscany | Central Italy | 1 root sucker, 1 top canopy | 441.058 | 105.262 | 6.15 |
| Locus | Na | Ne | Ho | He | uHe | Fis |
|---|---|---|---|---|---|---|
| CsCAT1 | 9 | 3.25 | 0.80 | 0.75 | 0.77 | −0.134 |
| CsCAT2 | 16 | 7.43 | 0.83 | 0.90 | 0.91 | 0.069 |
| CsCAT3 | 27 | 5.95 | 0.78 | 0.88 | 0.89 | 0.082 |
| CsCAT6 | 16 | 6.11 | 0.82 | 0.88 | 0.89 | 0.042 |
| CsCAT14 | 5 | 3.20 | 0.68 | 0.71 | 0.71 | 0.029 |
| CsCAT16 | 7 | 2.80 | 0.67 | 0.68 | 0.68 | −0.019 |
| EMCs15 | 8 | 2.97 | 0.53 | 0.70 | 0.71 | 0.228 ** |
| EMCs38 | 15 | 6.08 | 0.81 | 0.88 | 0.89 | 0.058 * |
| QrZAG7 | 4 | 1.69 | 0.24 | 0.45 | 0.46 | 0.422 ** |
| Mean | 11.88 | 4.40 | 0.68 | 0.75 | 0.76 | 0.088 |
| Sampling Site | N Genotypes | Na | Ne | Ho | He | uHe | Fis | Ar | Par |
|---|---|---|---|---|---|---|---|---|---|
| Sicily | 13 | 6.11 | 4.19 | 0.65 | 0.69 | 0.71 | 0.081 | 6.11 | 0.59 |
| Calabria | 30 | 8.44 | 4.56 | 0.65 | 0.73 | 0.74 | 0.089 * | 6.91 | 0.91 |
| Lazio-Umbria | 20 | 7.67 | 4.92 | 0.71 | 0.71 | 0.73 | −0.001 | 6.88 | 0.75 |
| Tuscany | 43 | 9.22 | 4.47 | 0.73 | 0.74 | 0.75 | 0.041 | 6.77 | 1.06 |
| Total | 106 | ||||||||
| Mean | 7.86 | 4.53 | 0.68 | 0.71 | 0.73 | 0.052 | 6.67 | 0.82 |
| Marker | L | Gaps (Min-Max) | p (Min-Max) | Eta | S | Pi | Theta (Per Site) | PICs | H | Hd |
|---|---|---|---|---|---|---|---|---|---|---|
| trnH-psbA | 528 | 3 (1–7) | (0.00–0.01) | 14 | 6 | 0.0009 | 0.0026 | 1 | 6 | 0.6772 |
| trnK-matK | 638 | 1 (1) | (0.00–0.006) | 5 | 4 | 0.0009 | 0.0014 | 3 | 4 | 0.3377 |
| Joined markers | 1166 | 4 (1–7) | (0.00–0.006) | 19 | 10 | 0.0009 | 0.002 | 4 | 10 | 0.7428 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioni, C.; Ranzino, L.; Cherubini, M.; Leonardi, L.; La Mantia, T.; Castellana, S.; Villani, F.; Simeone, M.C. Monuments Unveiled: Genetic Characterization of Large Old Chestnut (Castanea sativa Mill.) Trees Using Comparative Nuclear and Chloroplast DNA Analysis. Forests 2020, 11, 1118. https://doi.org/10.3390/f11101118
Mattioni C, Ranzino L, Cherubini M, Leonardi L, La Mantia T, Castellana S, Villani F, Simeone MC. Monuments Unveiled: Genetic Characterization of Large Old Chestnut (Castanea sativa Mill.) Trees Using Comparative Nuclear and Chloroplast DNA Analysis. Forests. 2020; 11(10):1118. https://doi.org/10.3390/f11101118
Chicago/Turabian StyleMattioni, Claudia, Liliana Ranzino, Marcello Cherubini, Luca Leonardi, Tommaso La Mantia, Simone Castellana, Fiorella Villani, and Marco Cosimo Simeone. 2020. "Monuments Unveiled: Genetic Characterization of Large Old Chestnut (Castanea sativa Mill.) Trees Using Comparative Nuclear and Chloroplast DNA Analysis" Forests 11, no. 10: 1118. https://doi.org/10.3390/f11101118
APA StyleMattioni, C., Ranzino, L., Cherubini, M., Leonardi, L., La Mantia, T., Castellana, S., Villani, F., & Simeone, M. C. (2020). Monuments Unveiled: Genetic Characterization of Large Old Chestnut (Castanea sativa Mill.) Trees Using Comparative Nuclear and Chloroplast DNA Analysis. Forests, 11(10), 1118. https://doi.org/10.3390/f11101118