1. Introduction
In recent years, the resilience of Norway spruce (
Picea abies (L.) H. Karst.) has been increasingly challenged by climate change [
1]. Climate-associated severe events, such as storms, droughts, and heat waves, amplify the susceptibility of Norway spruce to secondary damage, like bark beetle outbreaks [
1,
2]. This issue is more pronounced in regions where spruce trees have been planted outside their optimal natural ecological range, such as in lowland areas, and resulted in flooding of global markets with damaged timber in just a few years [
3]. As the frequency of climate-associated severe events will likely increase due to global warming [
4], bark beetle attacks will become more common due to the increasing amount of damaged timber.
Several past studies have examined what makes individual Norway spruce trees more susceptible to spruce bark beetle (
Ips typographus L.) attacks. The host selection strategy was found to be flexible [
5,
6,
7], attacking physiologically stressed trees first [
6] and then moving on to healthy trees as the population increases, thus enabling the stronger defense of healthy trees to be overcome. Past studies have confirmed that crown length, crown structure, and crown social class can also play a role in the susceptibility of Norway spruce trees to bark beetle attacks [
8,
9]. Trees from the dominant crown class were attacked more frequently than trees from other social classes. Furthermore, it was shown that height, diameter at breast height, and bark thickness were positively connected with bark beetle attacks, while phloem thickness had no influence [
9]. Diameter at breast height (DBH) and other tree properties (tree height, crown length ratio, age, and basal area) were confirmed as the best predictors of a bark beetle attack, with changes in tree mortality during the outbreak [
10].
In botany, the term bark refers to all tissue outside the vascular cambium in woody plants. It consists of dead outer bark, also called rhytidome, and living inner bark. Bark tissue is the product of the activity of two lateral meristems: the vascular cambium and the cork cambium. The former produces phloem, while the latter produces phellem (cork) and phelloderm [
11]. The bark structure is very diverse; it is composed of different cell types that perform numerous vital functions in the living tree: e.g., transport of photosynthates and signal molecules to the different parts of the tree; storage of non-structural carbohydrates and water and other compounds; mechanical support; and protection against solar radiation, drought, fire, frost, physical damage, plant pathogens, and herbivores [
12]. Apart from its crucial role for the performance of trees, the bark mostly remains an underutilized industrial by-product. It has mainly been used for energy production, while its potential for new value-added products is not yet fully exploited. However, numerous recent studies show a growing commercial interest towards its more efficient and comprehensive use, which is necessary for its transformation to a biocircular economy [
13]. In this respect, data on bark thickness on standing trees for different tree species and regions would allow more accurate estimates of the wood log volume without bark overall and availability of bark biomass to determine its potential use as a raw material [
14,
15]. The bark of Norway spruce has been relatively well studied (see, e.g., in [
13,
15,
16,
17]), which is undoubtedly due to the large share and economic value of spruce timber on global markets [
3]. Consequently, there is a great interest in developing a more efficient use of the large available quantities of spruce bark.
Tree mortality due to bark beetle attacks interacts with the presence of drought [
18], which is why adding only bark thickness to prediction models will likely not be enough. Other indicators that reflect the potential presence of drought or the general vitality of the examined trees are therefore needed, such as the vitality and vigor of the trees. Although they cannot be measured directly, they can be measured indirectly by various indicators, such as crown condition, stem radial growth, cambial electrical resistance, etc. [
19,
20]. An example of such a method is the impedance measurement in standing trees using twin-needle probes in cambium/xylem, which was confirmed to be useful for vitality assessment in trees [
20]. This method is based on the principle that trees with higher vitality have a higher moisture content, which is reflected in the amount of ions and cations in the vascular tissue [
20]. The measurements are relatively fast and non-destructive, meaning the method could be applied in practice. Additionally, Stephenson et al. [
21] have shown that drought can directly influence host tree selection, opening up the possibility of using sapwood moisture content as an indicator of drought stress to be included in vulnerability assessments of individual stands. Vigorous trees offer more resources to bark beetles due to their thicker phloem, provided that they can be “captured” by the bark beetles. Such trees are not attacked immediately at the beginning of the outbreak, but only in later stages. This adaptive host selection behavior increases the possibility to try to determine the ratio of stressed-to-vigorous trees in a given stand and to use such ratios in models to predict vulnerability to large-scale bark beetle attacks.
As demonstrated above, the properties of individual trees are among the most useful variables in predicting bark beetle attacks in Norway spruce. However, tree properties are often correlated to each other and can also be influenced by outside factors (e.g., site, exposition, and bedrock). This research study aims to investigate the effect of tree properties (crown social class, DBH, and tree height) on bark thickness and sapwood moisture content in Norway spruce at two different locations. The study has two specific hypothesis: (i) DBH and tree height positively influence bark thickness and sapwood moisture content, and (ii) crown social class influences the relationships between the two examined variables: DBH and tree height. The first hypothesis is based on the fact that healthier fast growing trees, with an often large diameter and tree height for a climax species like Norway spruce, have larger crowns and transport/store more organic matter. However, diameters at breast height do not always directly reflect tree’s social status, as the social status can change several times throughout a lifetime of a tree. The second hypothesis is based on the previous work of Krajnc et al. [
22], where different crown social classes of Norway spruce trees were found to have significantly different crown properties. This could have a direct effect on either bark thickness or sapwood moisture content, or an indirect effect by affecting the relationships between the two examined variables: DBH and tree height.
The two easily measurable variables could potentially be used to improve predictions of bark beetle attacks. It has been reported that bark thickness varies between attacked and non-attacked trees [
9], while the sapwood moisture content indirectly reflected the vitality and vigor of individual trees [
20]. However, it is very likely that both variables are influenced by other confounding factors, for example, DBH, tree height, crown social class, and site. Such relationships would need to be taken into account when using any of the listed variables in predicting bark beetle attacks. This study aims to quantify the direction and magnitude of the effect of the listed variables on bark thickness and sapwood moisture content, so that they can be appropriately included in future research studies.
2. Materials and Methods
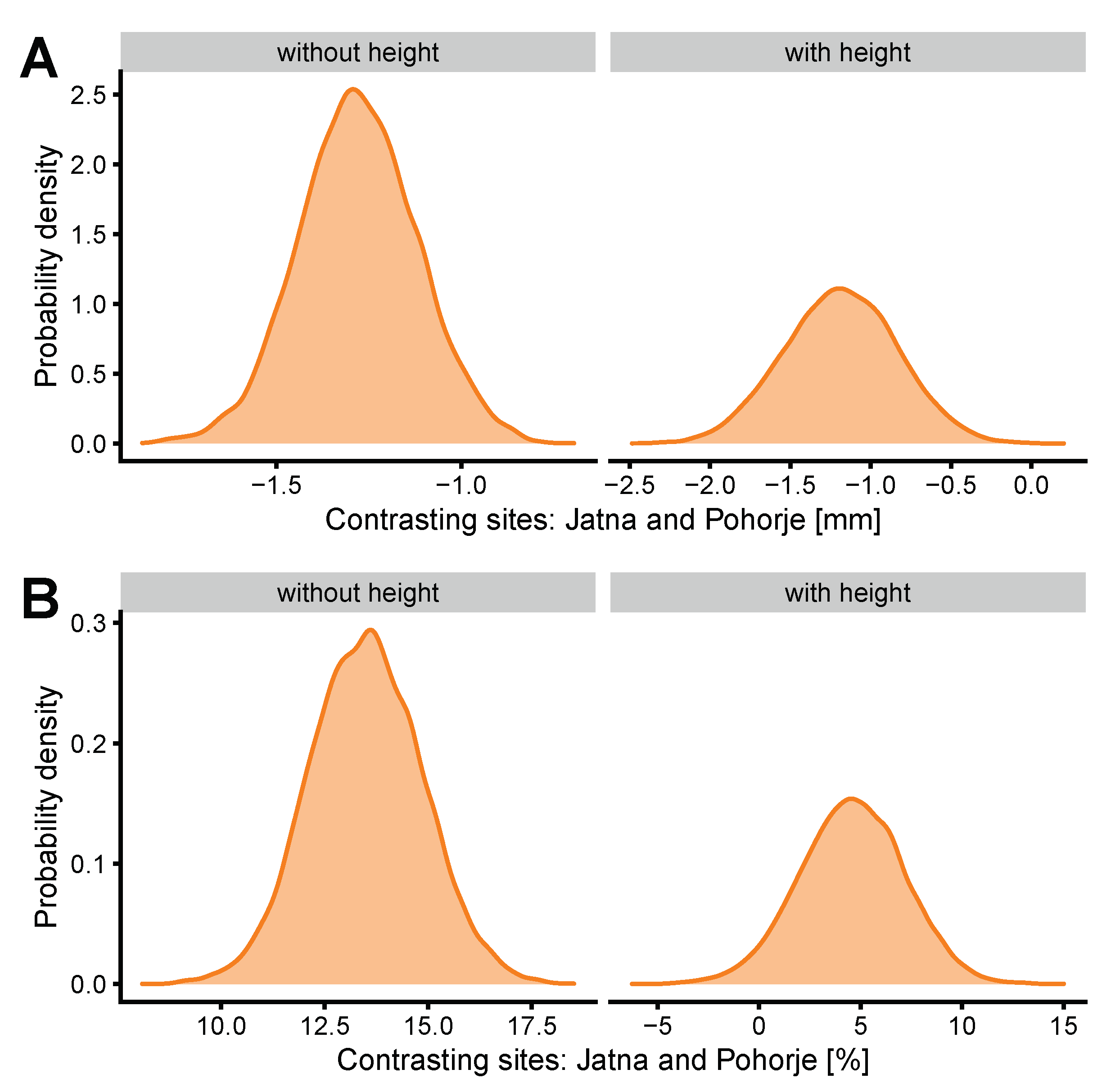
This research study was carried out in November 2019 on two even-aged monoculture stands of Norway spruce in Slovenia, Europe. The stands were approximately 120 years old and had been managed by selection thinning in the past. The first stand was located on Pohorje (NE Slovenia, 46°27′0.1″ N 15°25′18.1″ E, 1200 m a.s.l.) and the second one on Jatna (SE Slovenia, 46°2′30.2″ N 15°5′39.8″ E, 750 m a.s.l.). Both stands were of a similar site productivity, as determined by the volume tariff class for Norway spruce by the Slovenian Forest Service in their internal information system. The Pohorje stand was located on a higher elevation in an area with an extremely high probability of bark beetle attacks over the last few years, while the Jatna stand was located on a lower elevation in an area with a very low probability of bark beetle attacks [
23]. The two stands were included in the study to examine any potential differences between them that could have an influence on susceptibility to bark beetle attacks. The trees in each stand were randomly sampled, starting from one tree and then moving from one tree to next closest tree in the same cardinal direction. All trees were alive without any signs of being attacked by bark beetles at the time of measurement. Diameter at breast height (DBH) of each tree was measured using a diameter tape and each tree was assigned to one of the five crown social classes as proposed by the Kraft classification system, which uses relative crown size and position in relation to neighboring trees to classify individual trees [
24]. Tree height was also measured on a subset of trees with a Vertex IV hypsometer (Haglöf Sweden AB, Långsele, Sweden); the subset was created by measuring every other tree height. Bark thickness (BT) was measured in millimeters once per tree using a bark gauge (Haglöf Sweden AB, Långsele, Sweden), and sapwood moisture content (SMC) was measured using the Gann Hydromette RTU 600 moisture meter with the MH 34 active electrode (Gann Mess- u. Regeltechnik GmbH, Gerlingen, Germany), which is able to measure wood moisture content above fibre saturation point in coniferous wood. The values of the SMC are in given in percent, the depth of needle penetration was 16 millimeters, and the measurements were conducted in accordance with the manufacturer’s instructions. The electrode pins were perpendicular to the grain direction. As sapwood is responsible for water and nutrient transportation, its moisture content is above fiber saturation point, and has the highest moisture content when looking at radial moisture content distribution. The absolute values are season-dependent and also likely fluctuate daily due to the differences in evapotranspiration. All trees were measured by the same person in the same time span within an individual day (before noon) and in a span of a few days in the same season to avoid any potential influence of season, day, or person collecting the data. The data were imported and further analyzed in an open source statistical environment R [
25] using a Bayesian general linear model. This approach incorporates the uncertainty into the model itself and provides more informative conclusions. Two model sets were constructed: the first set on the complete data and the second on the smaller data subset with tree heights. Both sets were constructed with BT as the dependent variable, followed by SMC as the dependent variable. The four models were implemented in Stan [
26], with the help of the R package rethinking 2.11 [
27]. The priors used were weakly informative (see
Table 1), the model was specified as follows.
The two separate dependent variables were BT and SMC, while the independent variables were DBH, site, and crown social class in the first two models. An interaction between crown social class and DBH was also included in the model. In the second model set tree height was added, along with an interaction between tree height and crown social class. The two model sets are not directly comparable because the datasets were different between the two sets. However, they can provide an informative comparison. All models were checked for convergence and effective numbers of samples as recommended by McElreath [
28].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






