Winter Geometrid Moths in Oak Forests: Is Monitoring a Single Species Reliable to Predict Defoliation Risk?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Moths
2.2. Statistical Data Analysis
3. Results
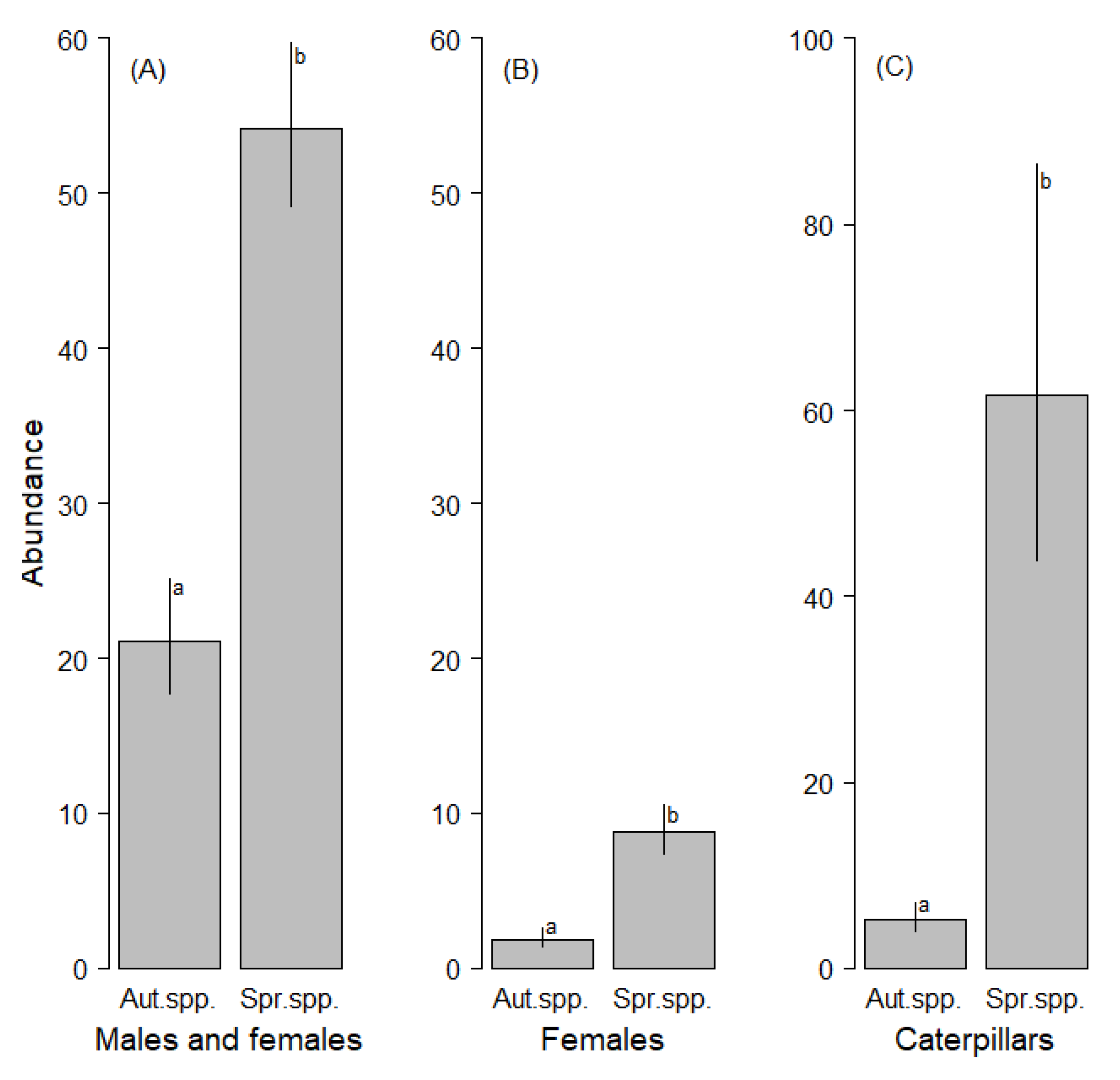
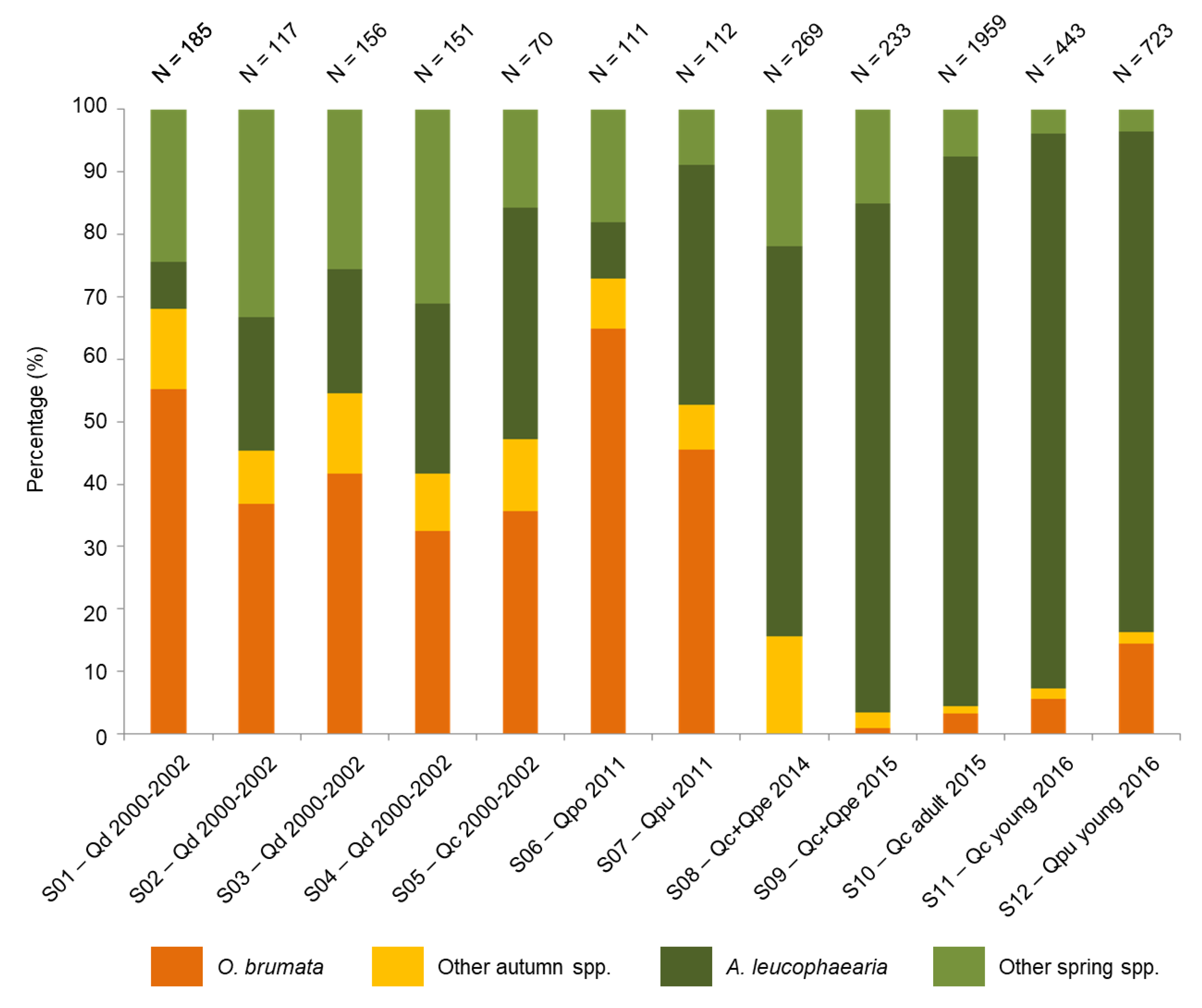
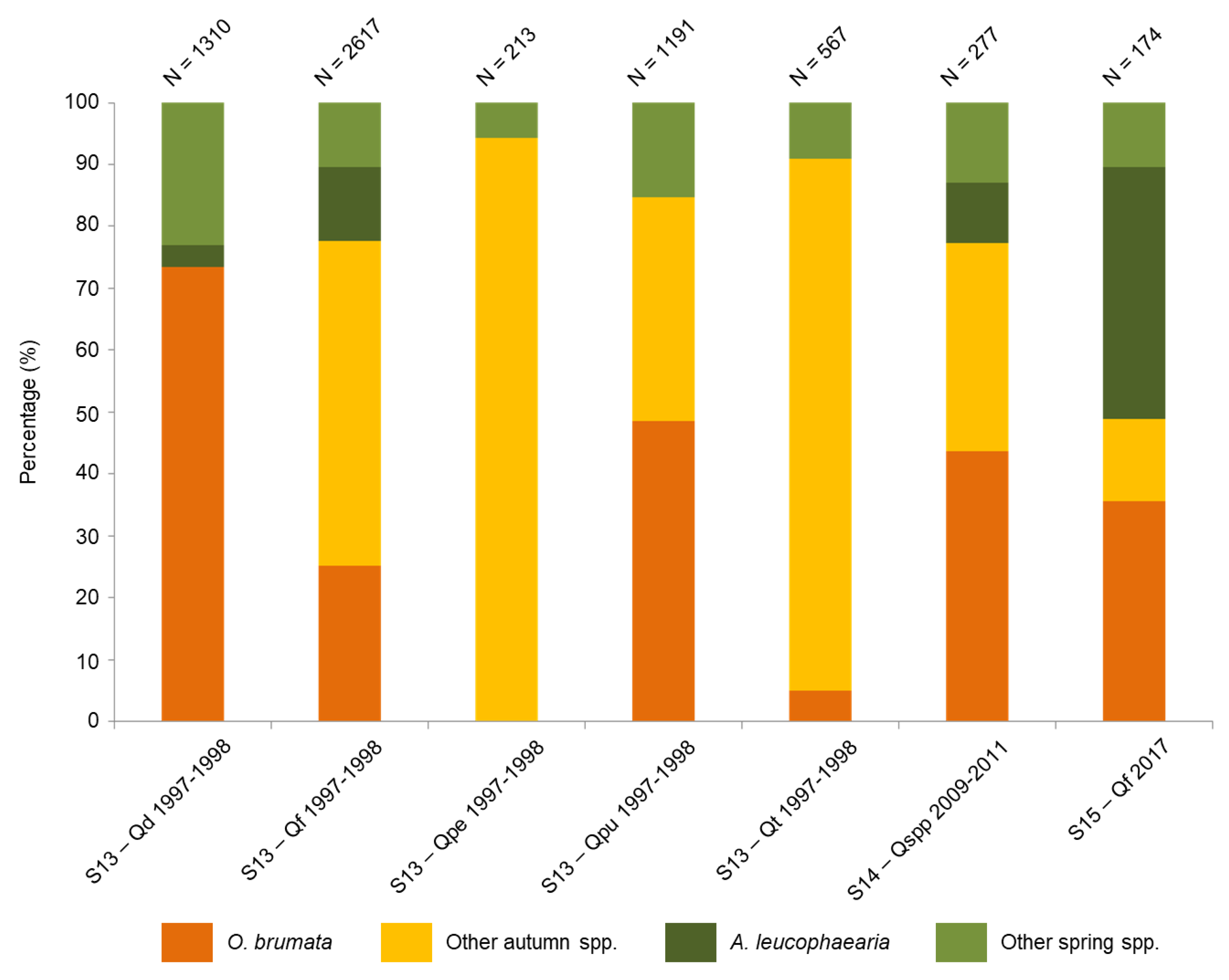
3.1. Assemblages of Adult Moths
3.2. Assemblages of Caterpillars
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Patočka, J. Húsenice na Duboch v ČSR, 1st ed.; Štátne pôdohospodárske nakladateľstvo: Bratislava, Czechoslovakia, 1954; p. 264. [Google Scholar]
- Leskó, K.; Szentkirályi, F.; Kádár, F. Araszoló lepkefajok fluktuáció-mintázatanak elemzése hosszú távú (1961–1997) magyarországi fénycsapdázási és kártételi idősorokon. Analysis of fluctuation patterns of geometrid moth species on long-term (1961–1997) time series of Hungarian light trappings and damage estimations. Erdész. Kutat. 1998, 88, 319–333. [Google Scholar]
- Patočka, J.; Krištín, A.; Kulfan, J.; Zach, P. Die Eichenschädlinge und Ihre Feinde, 1st ed.; Institut für Waldökologie der Slowakischen Akademie der Wissenschaften: Zvolen, Slovakia, 1999; p. 396. [Google Scholar]
- Neţoiu, C.; Tomescu, R.; Vladescu, D.; Aldea, D.I.; Buzatu, A. Evolution of infestations with loopermoth (Geometridae) in oak forests from Romania. Ann. Univ. Craiova –Agric. Montanology Cadastre Ser. 2014, 44, 301–310. [Google Scholar]
- Wahlberg, N.; Snäll, N.; Viidalepp, J.; Ruohomäki, K.; Tammaru, T. The evolution of female flightlessness among Ennominae of the Holarctic forest zone (Lepidoptera, Geometridae). Mol. Phylogenet. Evol. 2010, 55, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Schwenke, W. Die Forstschädlinge Europas, Ein Handbuch in Fünf Bäden. Autorenkollektiv: Band 3. Schmetterlinge, 1st ed.; Paul Parey: Hamburg, Berlin, Germany, 1978; p. 467. [Google Scholar]
- Csóka, G. Lepkehernyók (Caterpillars), 1st ed.; Agroinform Kiadóház: Budapest, Hungary, 1995; p. 151. [Google Scholar]
- Hunter, M.D. Interactions between Operophtera brumata and Tortrix viridana on oak: New evidence from time-series analysis. Ecol. Entomol. 1998, 23, 168–173. [Google Scholar] [CrossRef]
- Tikkanen, O.P.; Roininen, H. Spatial pattern of outbreaks of Operophtera brumata in eastern Fennoscandia and their effects on radial growth of trees. For. Ecol. Manag. 2001, 146, 45–54. [Google Scholar] [CrossRef]
- Simmons, M.J.; Lee, T.D.; Ducey, M.J.; Elkinton, J.S.; Boettner, G.H.; Dodds, K.J. Effects of invasive winter moth defoliation on tree radial growth in eastern Massachusetts, USA. Insects 2014, 5, 301–318. [Google Scholar] [CrossRef] [Green Version]
- Zúbrik, M.; Kunca, A.; Rell, S.; Galko, J.; Nikolov, C.; Leontovyč, R.; Gubka, A.; Vakula, J.; Špilda, I. Výskyt listožravých a cicavých hmyzích škodcov v lesoch Slovenska v období 1960–2015. In Aktuálne Problémy v Ochrane Lesa 2017, Zborník Referátov z 26. Medzinárodnej Konferencie Aktuálne Problémy v Ochrane Lesa 2017, Kongresové Centrum Kúpeľov Nový Smokovec, a.s., Slovakia, 26.–27.1.2017; Kunca, A., Ed.; Národné lesnícke centrum: Zvolen, Slovakia, 2017; pp. 54–70. [Google Scholar]
- Ambrus, A.; Csóka, G. Studien über das Schwärmen und die Dichte-Abschätzung des Frostspanners, Operophthera brumata L. (Lep., Geometridae) mit Hilfe von Markierungen und Pheromonfallen in Ungar. [Density estimation and swarming studies on the wintermoth, Operophthera brumata L. (Lep., Geometridae) by marking and using pheromone traps]. Anz. Schädlingskd. Pfl. Umwelt. 1992, 65, 88–92. [Google Scholar]
- Kolk, A.; Starzyk, J.R.; Kinelski, S.; Dzwonkowski, R. Atlas Szkodliwych Owadów Leśnych, 1st ed.; Multico O.W.: Warszawa, Poland, 1996; p. 705. [Google Scholar]
- Kunca, A.; Zúbrik, M.; Novotný, J. Škodlivé Činitele Lesných Drevín A Ochrana Pred Nimi, 1st ed.; Národné lesnícke centrum: Zvolen, Slovakia, 2007; p. 208. [Google Scholar]
- Connel, J. Wirtsbaumwahl durch aufbaumende Weibchen des Kleinen Frostspanners, Operophtera brumata L.: Ergebnisse einer vierjährigen Studie in Wien. Forstsch. Aktuell 2013, 57/58, 22–26. [Google Scholar]
- Tenow, O.; Nilssen, A.C.; Bylund, H.; Pettersson, R.; Battisti, A.; Bohn, U.; Caroulle, F.; Ciornei, C.; Csóka, G.; Delb, H.; et al. Geometrid outbreak waves travel across Europe. J. Animal Ecol. 2013, 82, 84–95. [Google Scholar] [CrossRef]
- Bublitz, T. Monitoring der Eichenschädlinge. FVA-Einblick 2016, 1, 15–17. [Google Scholar]
- Delb, H. Monitoring und Prognose der Schadorganismen im Wald: Eine Kernaufgabe des Waldschutzes. FVA-Einblick 2016, 1, 4–9. [Google Scholar]
- Horváth, B.; Ambrus, A. A tollascsápú araszoló (Colotis pennaria), nagy téliaraszoló (Erannis defoliaria) és aranysárga téliaraszoló (Erannis aurantiaria) vizsgálata jelölés-visszafogás módszerrel [Mark-recapture study on the featheres thorn (Colotis pennaria), mottled umber (Erannis defoliaria) and scarce umber (Erannis aurantiaria).]. Erdészettud. Közlem. 2017, 7, 59–67. [Google Scholar]
- Topp, W.; Kulfan, J.; Mergel, S.; Zach, P. Massenvermehrung von phyllophagen Schmetterlingen in Laubwäldern des Rheinlands. Anz. Schädlingskd. Pfl. Umwelt. 1998, 71, 88–93. [Google Scholar] [CrossRef]
- Csóka, G.; Szabóky, C. Checklist of herbivorous insects of native and exotic oaks in Hungary I. (Lepidoptera). Acta Silv. Lign. Hung. 2005, 1, 59–72. [Google Scholar]
- Berard, R.; Bordon, J.; Colomb, C.; Savourey, M.; Audibert, C.; Rozier, Y.; Clary, J. Les macrohétérocères de la région Rhône-Alpes. Cah. Mus. Confl. 2010, 1, 9–42. [Google Scholar]
- Glavendekić, M.M.; Medarević, M.J. Insect defoliators and their influence on oak forests in the Djerdap National Park, Serbia. Arch. Biol. Sci. 2010, 62, 1137–1141. [Google Scholar] [CrossRef]
- Tóth, B.; Babics, J.; Benedek, B. Contributions to knowledge of the geometrid fauna of Bulgaria and Greece, with four species new for the Greek fauna (Lepidoptera: Geometridae) (plate 12). Esperiana 2013, 18, 221–224. [Google Scholar]
- Georgieva, M.; Takov, D.; Georgiev, G.; Pilarska, D.; Pilarski, P.; Mirchev, P.; Humber, R. Studies on non-target phyllophagous insects in oak forests as potential hosts of Entomophaga maimaiga (Entomophthorales: Entomophthoraceae) in Bulgaria. Acta Zool. Bulgar. 2014, 66, 115–120. [Google Scholar]
- Kulfan, J.; Sarvašová, L.; Parák, M.; Dzurenko, M.; Zach, P. Can late flushing trees avoid attack by moth larvae in temperate forests? Plant Protect. Sci. 2018, 54, 272–283. [Google Scholar]
- Hébert, C.; St-Antoine, L. Oviposition trap to sample eggs of Operophtera bruceata (Lepidoptera: Geometridae) and other wingless geometrid species. Can. Entomol. 1999, 131, 557–565. [Google Scholar] [CrossRef]
- Macek, J.; Procházka, J.; Traxler, L. Motýli a Housenky Střední Evropy. Noční Motýli III. Píďalkovití, 1st ed.; Academia: Praha, Czech, 2012; p. 424. [Google Scholar]
- Patočka, J. Die Raupen und Puppen der Eichenschmetterlinge Mitteleuropas, 1st ed.; Paul Parey: Hamburg/Berlin, Germany, 1980; p. 188. [Google Scholar]
- Kalapanida, M.; Petrakis, P.V. Temporal partitioning in an assemblage of insect defoliators feeding on oak on a Mediterranean mountain. Eur. J. Entomol. 2012, 109, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Kulfan, M. Structure of lepidopterocenoses on oaks Quercus dalechampii and Q. cerris in Central Europe and estimation of the most important species. Mun. Ent. Zool. 2012, 7, 732–741. [Google Scholar]
- Parák, M.; Kulfan, J.; Svitok, M. Lepidopteran larval assemblages associated with three oak (Quercus spp.) species in the Čachtické Karpaty Mts (western Slovakia). Folia Faun. Slovaca 2012, 17, 247–256. (In Slovak) [Google Scholar]
- Liang, K.; Zeger, S. Longitudinal data analysis using generalized linear models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. Available online: https://www.R-project.org/ (accessed on 15 November 2019).
- Halekoh, U.; Højsgaard, S.; Yan, J. The R package geepack for generalized estimating equations. J. Stat. Soft. 2006, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Porter, J. The Colour Identification Guide to Caterpillars of the British Isles (Macrolepidoptera), 1st ed.; Viking: London, UK, 1997; p. 275. [Google Scholar]
- Zúbrik, M.; Kunca, A.; Csóka, G. Insects and Diseases Damaging Trees and Shrubs of Europe, 1st ed.; N.A.P. Édition: Verrières-le-Buisson, France, 2013; p. 536. [Google Scholar]
- Elton, C.S. The Forest Canopy: Herbivores. In The Pattern of Animal Communities, 1st ed.; Springer: Dordrecht, The Netherlands, 1966; pp. 190–209. [Google Scholar]
- Tomiczek, C.; Perny, B. Aktuelle Schäden an Bäumen im Stadtbereich. Forstsch. Aktuell 2005, 34, 2–6. [Google Scholar]
- Hikisz, J.; Soszynska-Maj, A. What moths fly in winter? The assemblage of moths active in a temperate deciduous forest during the cold season in central Poland. J. Entomol. Res. Soc. 2015, 17, 59–71. [Google Scholar]
- Csóka, G. Oak Defoliating Insects in Hungary. In Proceedings of the Population Dynamics, Impacts, and Integrated Management of Forest Defoliating Insects, Banská Štiavnica, Slovakia, 18–23 August 1996; McManus, M.L., Liebhold, A.M., Eds.; USDA Forest Service General Technical Report NE-247; USDA: Madison, WI, USA, 1998; pp. 334–335. [Google Scholar]
- Szabó, S.; Árnyas, E.M.; Tóthmérész, B.; Varga, Z. Long-term light trap study on the macro-moth (Lepidoptera: Macroheterocera) fauna of the Aggtelek National Park. Acta Zool. Acad. Sci. Hung. 2007, 53, 257–269. [Google Scholar]
- Tvermyr, S. Sex pheromone in females of Erannis aurantiaria Hb. and Erannis defoliaria Cl. (Lep., Geometridae). Nor. Entomol. Tidsskr. 1969, 16, 25–28. [Google Scholar]
- Purrini, V.K.; Skatulla, U. Über die natürlichen Krankheiten der Frostspanner Operophthera brumata L. und Erannis defoliaria Clerck (Lep., Geometridae) im Spessart, Bayern. Anz. Schädlingskd. Pfl. Umwelt. 1979, 52, 20–24. [Google Scholar] [CrossRef]
- Glavendekić, M.; Mihajlović, L. Fitofagni insekti u hrastovim šumama Nacionalnog parka Đerdap. [Phytophagous insects in oak forests of the National Park Djerdap]. Šumarstvo 2004, 4, 19–30. [Google Scholar]
- Extremera, F.M.; Cobo, A.; Pérez Rodríguez, M.C.; Pérez Guerrero, S.; Vargas Osuna, E. El complejo de lepidópteros defoliadores de Quercus en la provincia de Córdoba. Bol. Sanid. Veg. Plagas 2004, 30, 203–209. [Google Scholar]
- Martínez-Zurimendi, P.; Rosa y Cubo, E.; Domínguez-Domínguez, M. Defoliadores importantes del quejigo (Quercus faginea Lamck.) en Castilla y León (España). Bol. Sanid. Veg. Plagas 2011, 37, 145–161. [Google Scholar]
- Branco, M.; Bragança, H.; Sousa, E.; Phillips, A.J.L. Pests and diseases in Portuguese forestry: Current and new threats. In Forest Context and Policies in Portugal: Present and Future Challenges, 1st ed.; Reboredo, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 117–154. [Google Scholar]
- Fält-Nardmann, J.; Klemola, T.; Roth, M.; Ruohomäki, K.; Saikkonen, K. Northern geometrid forest pests (Lepidoptera: Geometridae) hatch at lower temperatures than their southern conspecifics: Implications of climate change. Eur. J. Entomol. 2016, 113, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Lecheva, I. Mass reproduction of geometrids (Lepidoptera, Geometridae) and fluctuation in the population densities in Bulgaria. Acta Entomol. Bulg. 1999, 5, 61–65. [Google Scholar]
- Tenow, O.; Nilssen, A.C.; Bylund, H.; Hogstad, O. Waves and synchrony in Epirrita autumnata/Operophtera brumata outbreaks. I. Lagged synchrony: Regionally, locally and among species. J. Animal Ecol. 2007, 76, 258–268. [Google Scholar] [CrossRef]
- Klemola, T.; Andersson, T.; Ruohomäki, K. Fecundity of the autumnal moth depends on pooled geometrid abundance without a time lag: Implications for cyclic population dynamics. J. Animal Ecol. 2008, 77, 597–604. [Google Scholar] [CrossRef]
- Hagen, S.B.; Jepsen, J.U.; Schott, T.; Ims, R.A. Spatially mismatched trophic dynamics: Cyclically outbreaking geometrids and their larval parasitoids. Biol. Lett. 2010, 6, 566–569. [Google Scholar] [CrossRef]
- Jepsen, J.U.; Kapari, L.; Hagen, S.B.; Schott, T.; Vindstad, O.P.L.; Nilssen, A.C.; Ims, R.A. Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-Arctic birch. Glob. Chang. Biol. 2011, 17, 2071–2083. [Google Scholar] [CrossRef]
- Chernev, T. Studies on some species of geometrids in relation to their prognosis. Rastit. Zashchita 1980, 28, 14–19. [Google Scholar]
- Ciornei, C.; Mihalache, G. Integrated control of species of Geometridae in oak forests of Romania. In Proceedings of the Population Dynamics, Impacts, and Integrated Management of Forest Defoliating Insects, Banská Štiavnica, Slovakia, 18–23 August 1996; McManus, M.L., Liebhold, A.M., Eds.; USDA Forest Service General Technical Report NE-247; USDA: Madison, WI, USA, 1998; pp. 222–229. [Google Scholar]
- Rubtsov, V.V.; Utkina, I.A. Long-term dynamics of Operophtera brumata L. in the oak stands of forest-steppe. Contemp. Probl. Ecol. 2011, 4, 777–783. [Google Scholar] [CrossRef]
- Vakula, J.; Zúbrik, M.; Kunca, A.; Dubec, M.; Finďo, S.; Galko, J.; Kaštier, P.; Konôpka, B.; Lalkovič, M.; Leontovyč, R.; et al. Nové Metódy Ochrany Lesa; NLC: Zvolen, Slovak, 2015; p. 291. [Google Scholar]
- Albert, V.R.; Bogenschütz, H.; König, E. Studies on the use of pheromone traps for monitoring the population dynamics of Operophthera brumata L. (Lepid., Geometridae). Z. Angew. Entomol. 1984, 98, 286–298. [Google Scholar] [CrossRef]
- Stoakley, J.T. Outbreaks of Winter moth, Operophthera brumata L. (Lep., Geometridae) in young plantations of Sitka spruce in Scotland 1: Insecticidal control and population assessment using the sex attractant pheromone. Z. Angew. Entomol. 1985, 99, 153–160. [Google Scholar] [CrossRef]
- Hand, S.C.; Ellis, N.W.; Stoakley, J.T. Development of a pheromone monitoring system for the winter moth, Operophtera brumata (L.), in apples and in Sitka spruce. Crop Prot. 1987, 6, 191–196. [Google Scholar] [CrossRef]
- Cuming, F.G. The distribution, life history, and economic importance of the winter moth, Operophtera brumata (L.) (Lepidoptera, Geometridae) in Nova Scotia. Can. Entomol. 1961, 93, 135–142. [Google Scholar] [CrossRef]
- Hausmann, A. The Geometrid Moths of Europe, 1st ed.; Apollo Books: Stenstrup, Denmark, 2001; Volume 1, p. 282. [Google Scholar]
- Redondo, V.M.; Gastón, J.; Gimeno, R. Geometridae Ibericae, 1st ed.; Apollo Books: Stenstrup, Denmark, 2009; p. 361. [Google Scholar]
- Hausmann, A.; Viidalepp, J. The Geometrid Moths of Europe, 1st ed.; Apollo Books: Stenstrup, Denmark, 2012; Volume 3, p. 743. [Google Scholar]
- Topp, W.; Kirsten, K. Synchronisation of pre-imaginal development and reproductive success in the winter moth, Operophtera brumata L. J. Appl. Entomol. 1991, 111, 137–146. [Google Scholar] [CrossRef]
- Mrkva, R. Bionomie píďalky podzimní (Operophtera brumata L.)—Motýl a vajíčko. Acta Univ. Agric. (Brno) Ser. C 1968, 37, 223–246. (In Czech) [Google Scholar]
- Graf, B.; Borer, F.; Höpli, H.U.; Hohn, H.; Dorn, S. The winter moth Operophtera brumata L. (Lep., Geometridae) on apple and cherry: Spatial and temporal aspects of recolonization in autumn. J. Appl. Entomol. 1995, 119, 295–301. [Google Scholar] [CrossRef]
- Szöcz, G.; Tóth, M. Evidence and extraction of a female sex pheromone from the winter moth Operophtera brumata (L.) [forest pest, Hungary]. Acta Phytopathol. Acad. Sci. Hung. 1978, 3, 213–217. [Google Scholar]
- Roelofs, W.L.; Hill, A.S.; Linn, C.E.; Meinwald, J.; Jain, S.C.; Herbert, H.J.; Smith, R.F. Sex pheromone of the winter moth, a geometrid with unusually low temperature precopulatory responses. Sci. New Ser. 1982, 217, 657–659. [Google Scholar] [CrossRef]
- Szöcz, G.; Tóth, M.; Francke, W.; Schmidt, F.; Philipp, P.; König, W.A.; Mori, K.; Hansson, B.S.; Löfstedt, C. Species discrimination in five species of winter-flying geometrids (Lepidoptera) based on chirality of semiochemicals and flight season. J. Chem. Ecol. 1993, 19, 2721–2735. [Google Scholar] [CrossRef]
- Van Dongen, S.; Backeljau, T.; Matthysen, E.; Dhondt, A.A. Genetic population structure of the winter moth (Operophtera brumata L.) (Lepidoptera, Geometridae) in a fragmented landscape. Heredity (Edinb) 1998, 80, 92–100. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; LAU, T.F. Sexual dimorphism in the compound eye of the moth Operophtera brumata (Lepidoptera, Geometridae). Invertebr. Biol. 2008, 127, 201–216. [Google Scholar] [CrossRef]
- Kulfan, J.; Sarvašová, L.; Parák, M.; Zach, P. Effects of a host tree on movement and distribution of winter geometrid moths (Lepidoptera): Thickness of trunks and branches. Folia Biol. Oecologica 2019, 46, 83–90. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample | Country | Area | Coordinates | Oak Species | Year | Sample | Data Source |
|---|---|---|---|---|---|---|---|
| S01 | W SVK | Malé Karpaty Mountains | 48°19′ N 17°17′ E | Q. dalechampii | 2000–2002 | 150 branches | Kulfan (2012) |
| S02 | W SVK | Malé Karpaty Mountains | 48°22′ N 17°19′ E | Q. dalechampii | 2000–2002 | 150 branches | Kulfan (2012) |
| S03 | W SVK | Malé Karpaty Mountains | 48°29′ N 17°23′ E | Q. dalechampii | 2000–2002 | 225 branches | Kulfan (2012) |
| S04 | W SVK | Malé Karpaty Mountains | 48°32′ N 17°31′ E | Q. dalechampii | 2000–2002 | 150 branches | Kulfan (2012) |
| S05 | W SVK | Malé Karpaty Mountains | 48°32′ N 17°31′ E | Q. cerris | 2000–2002 | 225 branches | Kulfan (2012) |
| S06 | W SVK | Malé Karpaty Mountains | 48°44′ N 17°46′ E | Q. polycarpa | 2011 | 340 branches | Parák et al. (2012) |
| S07 | W SVK | Malé Karpaty Mountains | 48°44′ N 17°46′ E | Q. pubescens | 2011 | 140 branches | Parák et al. (2012) |
| S08 | SW SVK | Podunajská pahorkatina upland | 48°12′ N 18°24′ E | Q. cerris, Q. petraea | 2014 | 100 branches | original data |
| S09 | SW SVK | Podunajská pahorkatina upland | 48°12′ N 18°24′ E | Q. cerris, Q. petraea | 2015 | 100 branches | original data |
| S10 | S SVK | Krupinská planina plateau | 48°10′ N 18°59′ E | Q. cerris (adult trees) | 2015 | 102 branches | original data |
| S11 | S SVK | Krupinská planina plateau | 48°10′ N 18°59′ E | Q. cerris (young trees) | 2016 | 60 branches | original data |
| S12 | S SVK | Krupinská planina plateau | 48°10′ N 18°59′ E | Q. pubescens (young trees) | 2016 | 60 branches | original data |
| S13 | N GRC | Mount Holomontas | 40°25′ N 23°30′ E | Q. dalechampii, Q. frainetto, Q. trojana, Q. pubescens, Q. pedunculiflora | 1997–1998 | ≈ 710 branches | Kalapanida and Petrakis (2012) |
| S14 | BGR (N, S, E, W, Central) | 42°03′ N 24°54′ E | Q. cerris, Q. frainetto, Q. petraea, Q. robur, Q. pubescens | 2009–2011 | 300 oaks | Georgieva et al. (2014) | |
| S15 | S BGR | Rhodope Mountains | 41°20′ N 25°22′ E | Q. frainetto | 2017 | 540 branches | original data |
| Moth Group/Species | Wald Statistics | Stat. Significance | Corel. Parameter α |
|---|---|---|---|
| ‘spring species’ (males + females) | 98.7 | p < 0.0001 | 0.18 |
| vs. ‘autumn species’ (males + females) | |||
| ‘spring species’ (females) | 72.9 | p < 0.0001 | 0.02 |
| vs. ‘autumn species’ (caterpillars) | |||
| Agriopis leucophaearia (males + females) | 178.0 | p < 0.0001 | 0.20 |
| vs. Operophtera brumata (males + females) | |||
| Agriopis leucophaearia (females) | 101.4 | p < 0.0001 | 0.02 |
| vs. Operophtera brumata (females) | |||
| ‘spring species’ (caterpillars) | 140.3 | p < 0.0001 | 0.07 |
| vs. ‘autumn species’ (caterpillars) | |||
| Agriopis leucophaearia (caterpillars) | 136.5 | p < 0.0001 | 0.07 |
| vs. Operophtera brumata (caterpillars) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarvašová, L.; Kulfan, J.; Saniga, M.; Zúbrik, M.; Zach, P. Winter Geometrid Moths in Oak Forests: Is Monitoring a Single Species Reliable to Predict Defoliation Risk? Forests 2020, 11, 288. https://doi.org/10.3390/f11030288
Sarvašová L, Kulfan J, Saniga M, Zúbrik M, Zach P. Winter Geometrid Moths in Oak Forests: Is Monitoring a Single Species Reliable to Predict Defoliation Risk? Forests. 2020; 11(3):288. https://doi.org/10.3390/f11030288
Chicago/Turabian StyleSarvašová, Lenka, Ján Kulfan, Miroslav Saniga, Milan Zúbrik, and Peter Zach. 2020. "Winter Geometrid Moths in Oak Forests: Is Monitoring a Single Species Reliable to Predict Defoliation Risk?" Forests 11, no. 3: 288. https://doi.org/10.3390/f11030288
APA StyleSarvašová, L., Kulfan, J., Saniga, M., Zúbrik, M., & Zach, P. (2020). Winter Geometrid Moths in Oak Forests: Is Monitoring a Single Species Reliable to Predict Defoliation Risk? Forests, 11(3), 288. https://doi.org/10.3390/f11030288