Effect of Micromelalopha sieversi (Staudinger) Oviposition Behavior on the Transcriptome of Two Populus Section Aigeiros Clones



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Plant Treatments
2.3. RNA Quality Detection and cDNA Library Preparation
2.4. Sequence Alignment and Analysis of Differential Gene Expression
3. Results
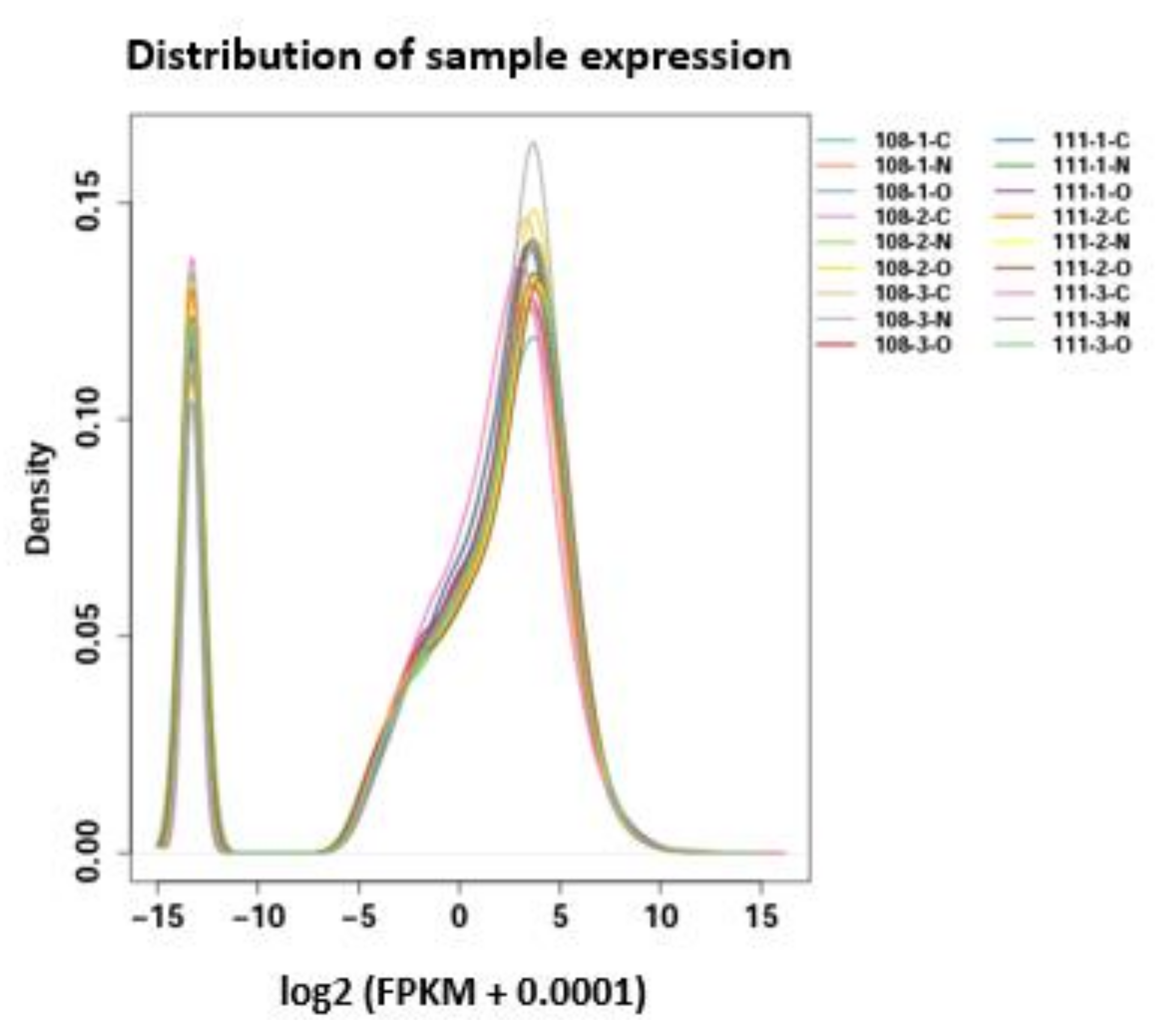
3.1. Transcriptome Sequencing and Reference Genome Comparisons of Each Treatment
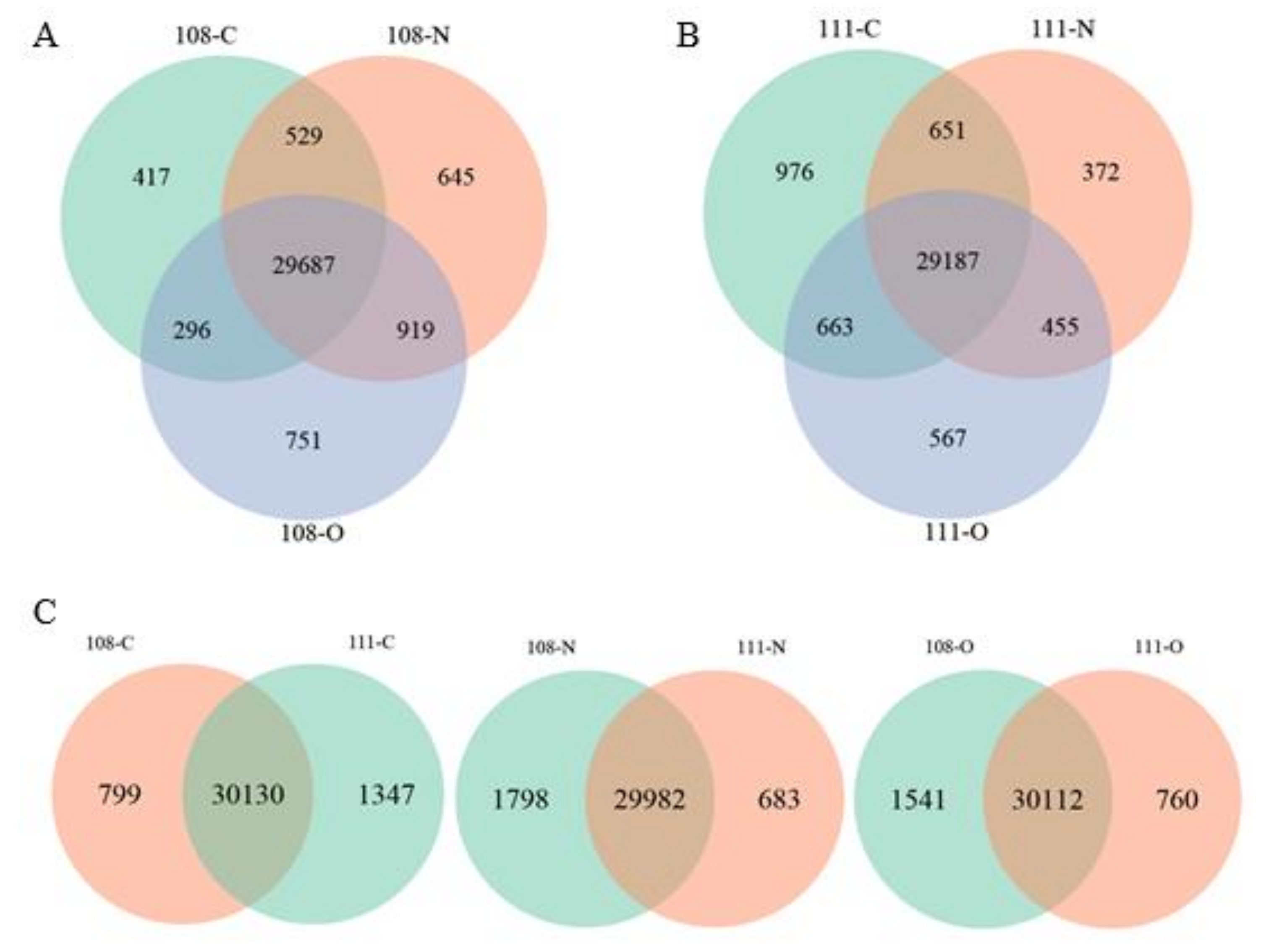
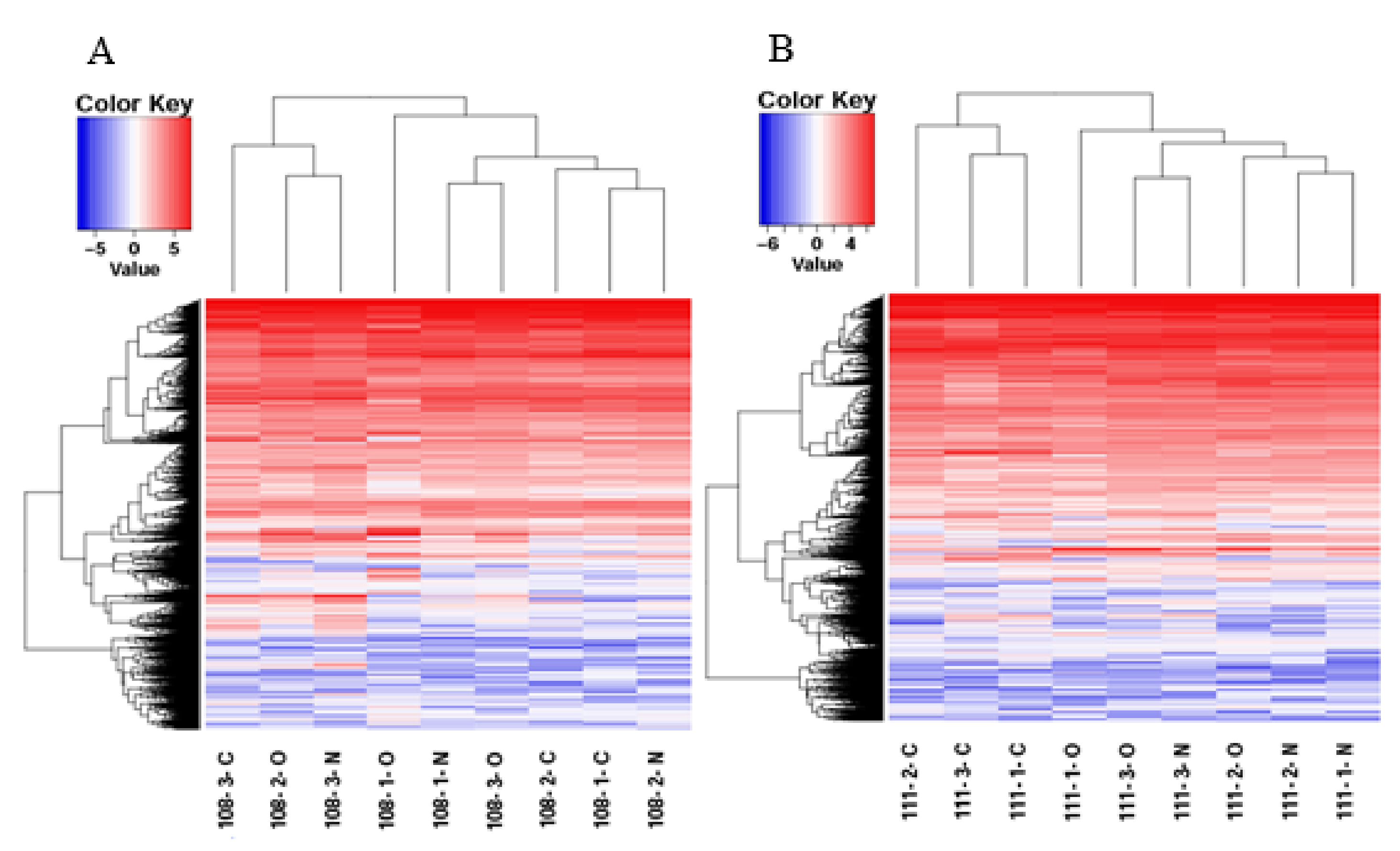
3.2. Gene Expression Analysis of the Different Treatment Groups
3.3. Analysis of DEGs
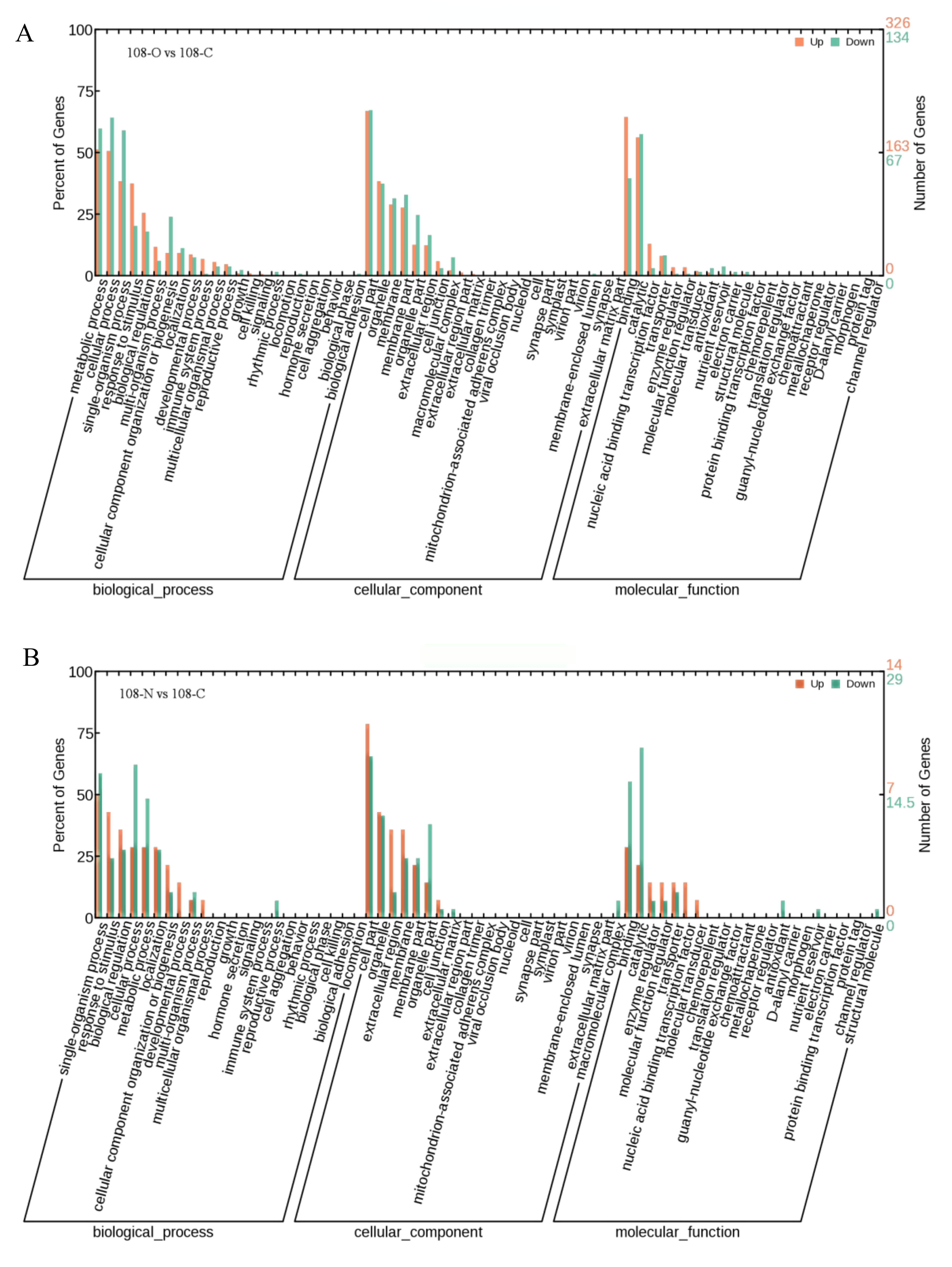
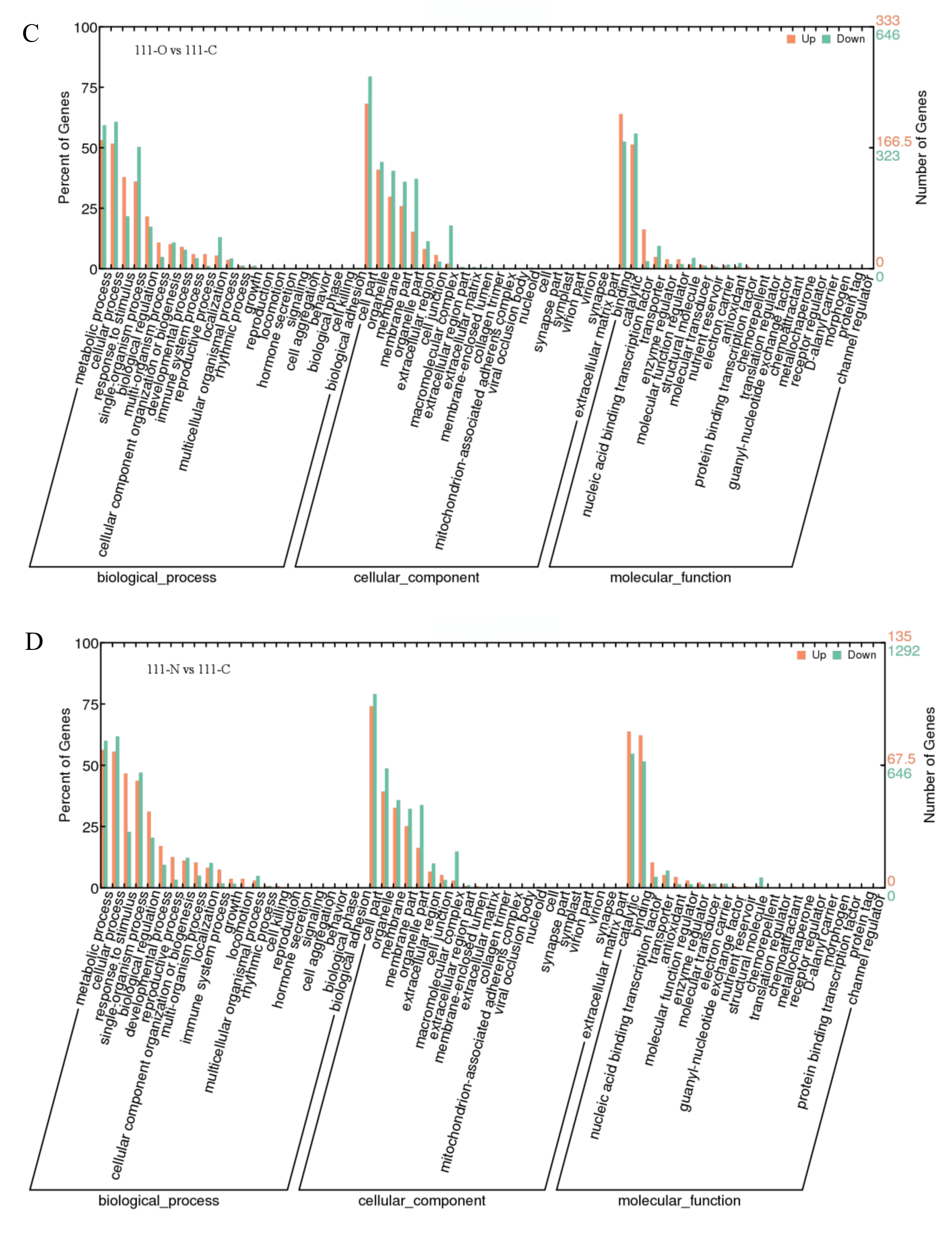
3.4. GO Functional Enrichment Analysis of DEGs
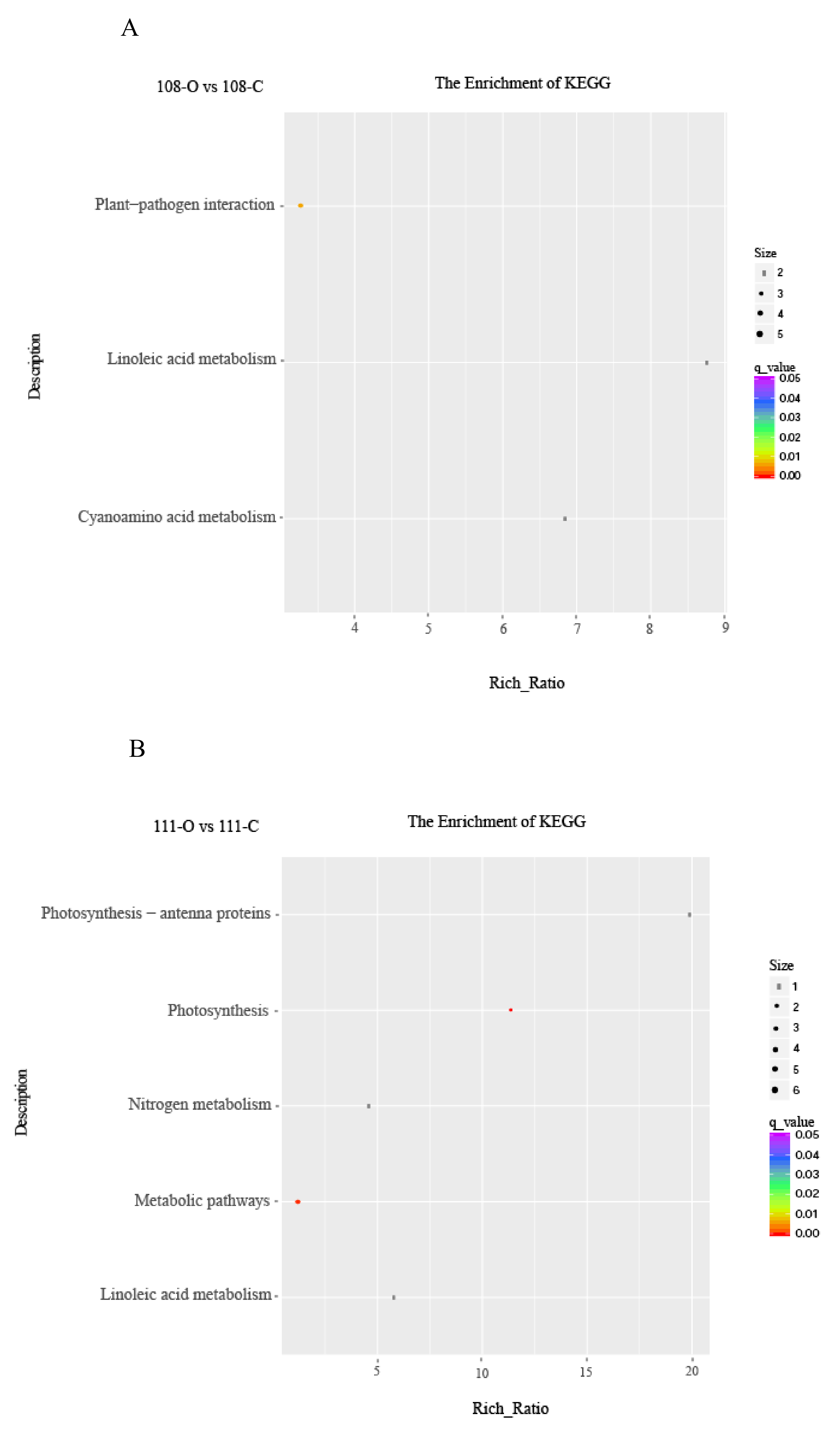
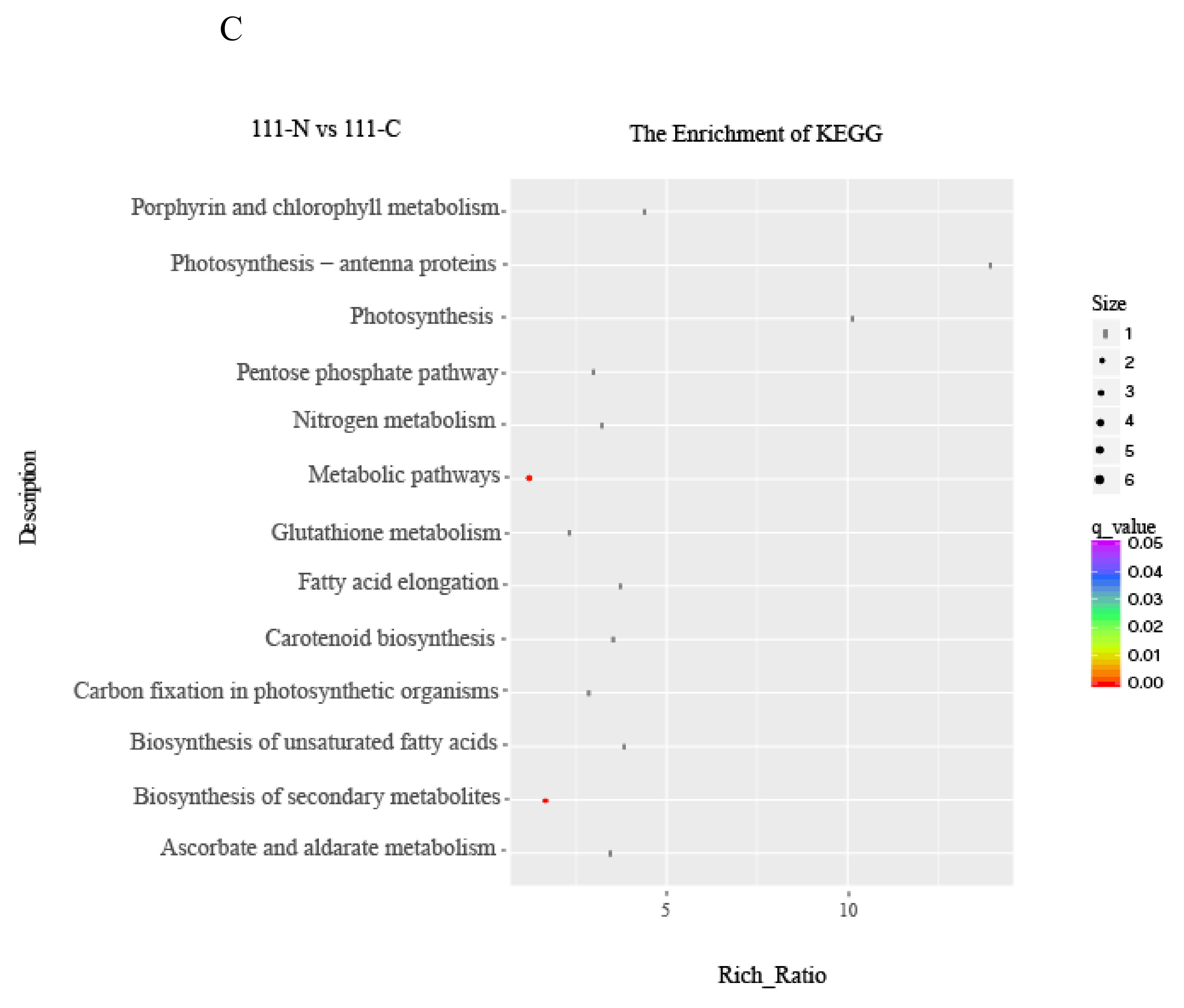
3.5. Pathway Enrichment Analysis of DEGs
3.6. Differential Expression of Important Defense Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Futuyma, D.J.; Mitter, C. Insect-plant interactions: The evolution of component communities. Philos. Trans. R. Soc. B 1996, 351, 1361–1366. [Google Scholar]
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Ann. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef]
- Jwa, N.S.; Agrawal, G.K.; Tamogami, S.; Yonekura, M. Role of defense/stress-related marker genes, proteins and secondary metabolites in defining rice self-defense mechanisms. Plant Physiol. Biochem. 2006, 44, 261–273. [Google Scholar] [CrossRef]
- Doss, R.P.; Proebsting, W.M.; Potter, S.W.; Clement, S.L. Response of Np mutant of pea (Pisum sativum L.) to pea weevil (Bruchus pisorum L.) oviposition and extracts. J. Chem. Ecol. 1995, 21, 97–106. [Google Scholar] [CrossRef]
- Doss, R.P.; Oliver, J.E.; Proebsting, W.M.; Potter, S.W.; Kuy, S.R.; Clement, S.L.; Williamson, R.T.; Carney, J.R.; Devilbiss, E.D. Bruchins-insect-derived plant regulators that stimulate neoplasm formation. Proc. Natl. Acad. Sci. USA 2000, 97, 6218–6223. [Google Scholar] [CrossRef] [Green Version]
- Hilker, M.; Fatouros, N.E. Plant responses to insect egg deposition. Ann. Rev. Entomol. 2015, 60, 493–515. [Google Scholar] [CrossRef]
- Schröder, R.; Forstreuter, M.; Hilker, M. A plant notices insect egg deposition and changes its rate of photosynthesis. Plant Physiol. 2005, 138, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Blenn, B.; Bandoly, M.; Küffner, A.; Otte, T.; Geiselhardt, S.; Fatouros, N.E.; Hilker, M. Insect egg deposition induces indirect defense and epicuticular wax changes in Arabidopsis thaliana. J. Chem. Ecol. 2012, 38, 882–892. [Google Scholar] [CrossRef]
- Conti, E.; Salerno, G.; Leombruni, B.; Frati, F.; Bin, F. Short-range allelochemicals from a plant-herbivore association: A singular case of oviposition-induced synomone for an egg parasitoid. J. Exp. Biol. 2010, 213, 3911–3919. [Google Scholar] [CrossRef] [Green Version]
- Fatouros, N.E.; Lucas-Barbosa, D.; Weldegergis, B.T.; Pashalidou, F.G.; van Loon, J.J.A. Plant volatiles induced by herbivore egg deposition affect insects of different trophic levels. PLoS ONE 2012, 7, e43607. [Google Scholar] [CrossRef]
- Meiners, T.; Hilker, M. Induction of plant synomones by oviposition of a phytophagous insect. J. Chem. Ecol. 2000, 26, 221–232. [Google Scholar] [CrossRef]
- Hilker, M.; Kobs, C.; Varama, M.; Schrank, K. Insect egg deposition induces Pinus sylvestris to attract egg parasitoids. J. Exp. Biol. 2002, 205, 455–461. [Google Scholar] [PubMed]
- Colazza, S.; Fucarino, A.; Peri, E.; Salerno, G.; Conti, E.; Bin, F. Insect oviposition induces volatile emission in herbaceous plants that attracts egg parasitoids. J. Exp. Biol. 2004, 207, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.R.; Goggin, F.L. Effects of jasmonate-induced defenses in tomato on the potato aphid, Macrosiphum Euphorbiae. Entomol. Exp. Appl. 2005, 115, 107–115. [Google Scholar] [CrossRef]
- Geiselhardt, S.; Yoneya, K.; Blenn, B.; Drechsler, N.; Gershenzon, J.; Kunze, R.; Hilker, M. Egg laying of cabbage white butterfly (Pieris brassicae) on Arabidopsis thaliana affects subsequent performance of the larvae. PLoS ONE 2013, 8, e59661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Tooker, J.F.; Luthe, D.S.; Moraes, C.M.D.; Felton, G.W. Insect eggs can enhance wound response in plants: A study system of tomato Solanum lycopersicum L. and Helicoverpa zea Boddie. PLoS ONE 2012, 7, e37420. [Google Scholar] [CrossRef] [Green Version]
- Leopold, R.A.; Degrugillier, M.E. Sperm penetration of housefly eggs: Evidence for involvement of a female accessory secretion. Science 1973, 181, 555–557. [Google Scholar] [CrossRef]
- Köpke, D.; Beyaert, I.; Gershenzon, J.; Hilker, M.; Schmidt, A. Species-specific responses of pine sesquiterpene synthases to sawfly oviposition. Phytochemistry 2010, 71, 909–917. [Google Scholar] [CrossRef]
- Beyaert, I.; Köpke, D.; Stiller, J.; Hammerbacher, A.; Yoneya, K.; Schmidt, A.; Gershenzon, J.; Hilker, M. Can insect egg deposition ‘warn’ a plant of future feeding damage by herbivorous larvae? Proc. Biol. Sci. 2012, 279, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Ryan, C.A. Protease Inhibitors in plants: Genes for improving defenses against insects and pathogens. Ann. Rev. Phytopathol. 1990, 28, 425–449. [Google Scholar] [CrossRef]
- Little, D.; Gouhier-Darimont, C.; Bruessow, F.; Reymond, P. Oviposition by Pierid Butterflies Triggers Defense Responses in Arabidopsis. Plant Physiol. 2007, 143, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büchel, K.; McDowell, E.; Nelson, W.; Descour, A.; Gershenzon, J.; Hilker, M.; Soderlund, C.; Gang, D.R.; Fenning, T.; Meiners, T. An elm EST database for identifying leaf beetle egg-induced defense genes. BMC Genom. 2012, 13, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, T.P.; Uknes, S.; Vernooij, B.; Friedrich, L.; Weymann, K.; Negrotto, D.; Gaffney, T.; Gut-Rella, M.; Kessmann, H.; Ward, E.; et al. A central role of salicylic acid in plant disease resistance. Science 1994, 266, 1247–1250. [Google Scholar] [CrossRef] [Green Version]
- Gundlach, H.; Müller, M.J.; Kutchan, T.M.; Zenk, M.H. Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl. Acad. Sci. USA 1992, 89, 2389–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, C.; Dixon, R.A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. 1997, 48, 251–275. [Google Scholar] [CrossRef]
- Reymond, P. Perception, signaling and molecular basis of oviposition-mediated plant responses. Planta 2013, 238, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Buchala, A. Insect eggs suppress plant defence against chewing herbivores. Plant J. 2010, 62, 876–885. [Google Scholar]
- Gouhier-Darimont, C.; Schmiesing, A.; Bonnet, C.; Lassueur, S.; Reymond, P. Signalling of Arabidopsis thaliana response to Pieris brassicae eggs shares similarities with PAMP-triggered immunity. J. Exp. Bot. 2013, 64, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- Ding, C.J.; Zhang, W.X.; Gao, M.; Huang, Q.J.; Chu, Y.G.; Su, X.H. Analysis of transcriptome differences among Populus deltoides with different growth potentials. Sci. Silvae Sin. 2016, 52, 47–58. [Google Scholar]
- Guo, L.; Liu, F.; Zhang, S.F.; Kong, X.B.; Zhang, Z. Egg deposition of Micromelalopha sieversi (Staudinger) on clones of Populus from section Aigeiros induces resistance in neighboring plants. Forests 2019, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Baren, M.J.V.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and abundance estimation from RNA-Seq reveals thousands of new transcripts and switching among isoforms. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Loon, L.C.V.; Pierpoint, W.S.; Boller, T.H.; Conejero, V. Recommendations for naming plant pathogenesis-related proteins. Plant Mol. Biol. Rep. 1994, 12, 245–264. [Google Scholar] [CrossRef]
- Sels, J.; Mathys, J.; De, C.; Barbara, M.A.; Cammue, B.P.A.; De, B.; Miguel, F.C. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef]
- Mosolov, V.V.; Valueva, T.A. Proteinase inhibitors and their function in plants: A review. Appl. Biochem. Microbiol. 2005, 41, 227–261. [Google Scholar] [CrossRef]
- Yang, G.; Guan, X.; Wang-Pruski, G.; Wei, H.; You, M.S. Advance on induced plant defense genes against insect pests. J. Agric. Biotechnol. 2007, 15, 157–166. [Google Scholar]
- Eyles, A.; Bonello, P.; Ganley, R.; Mohammed, C. Induced resistance to pests and pathogens in trees. New Phytol. 2010, 185, 893–908. [Google Scholar] [CrossRef]
- Anderson, M.A.; Van, H.R.; West, J.; Bateman, K.; Lee, M.; Christeller, J.T.; McDonald, G.; Heath, R.L. Proteinase inhibitors from Nicotiana alata enhance plant resistance to insect pests. J. Insect Physiol. 1997, 43, 833. [Google Scholar]
- Hollick, J.B.; Gordon, M.P. A poplar tree proteinase inhibitor-like gene promoter is responsive to wounding in transgenic tobacco. Plant Mol. Biol. 1993, 22, 561–572. [Google Scholar] [CrossRef]
- Philippe, R.N.; Bohlmann, J. Poplar defense against insect herbivores. Rev. Can. De Bot. 2007, 85, 1111–1126. [Google Scholar] [CrossRef]
- Deng, M.R.; Long, L.K.; Zhu, H.H.; Qu, X.Y. Recombinant expression and antimicrobial assay of Hevein-like peptide AbAMP1. Microbiology 2008, 35, 5–9. [Google Scholar]
- Zhao, M.; Ma, Y.; Pan, Y.H.; Zhang, C.H.; Yuan, W.X. A hevein-like protein and a class I chitinase with antifungal activity from leaves of the paper mulberry. Biomed. Chromatogr. 2011, 25, 908–912. [Google Scholar] [CrossRef]
- Zwicker, S.; Mast, S.; Stos, V.; Pfitzner, A.J.P.; Pfitzner, U.M. Tobacco NIMIN2 proteins control PR gene induction through transient repression early in systemic acquired resistance. Mol. Plant Pathol. 2010, 8, 385–400. [Google Scholar] [CrossRef]
- Weigel, R.R.; Bäuscher, C.; Pfitzner, A.J.; Pfitzner, A.J.P.; Pfitzner, U.M. NIMIN-1, NIMIN-2 and NIMIN-3, members of a novel family of proteins from Arabidopsis that interact with NPR1/NIM1, a key regulator of systemic acquired resistance in plants. Plant Mol. Biol. 2001, 46, 143–160. [Google Scholar] [CrossRef]
- Spoel, S.H.; Koornneef, A.; Claessens, S.M.; Korzelius, P.; Pelt, J.A.V.; Mueller, M.J.; Buchala, A.J.; Métraux, J.; Brown, R.; Kazan, K.; et al. NPR1 modulates cross-talk between salicylate- and jasmonate-dependent defense pathways through a novel function in the cytosol. Plant Cell 2003, 15, 760–770. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Calis, O.; Patrick, E.; Zhang, G.; Charoenwattana, P.; Muskett, P.; Parker, J.E.; Turner, J.G. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis. Plant J. 2010, 42, 95–110. [Google Scholar] [CrossRef]
- Wang, W.; Devoto, A.; Turner, J.G.; Xiao, S.Y. Expression of the membrane-associated resistance protein RPW8 enhances basal defense against biotrophic pathogens. Mol. Plant Microbe Interact. 2007, 20, 966. [Google Scholar] [CrossRef] [Green Version]
- Thygesen, P.W.; Dry, I.B.; Robinson, S.P. Polyphenol oxidase in potato. A multigene family that exhibits differential expression patterns. Plant Physiol. 1995, 109, 525–531. [Google Scholar] [PubMed] [Green Version]
- Mchenry, J.Z.; Christeller, J.T.; Slade, E.A.; Laing, W.A. The major extracellular proteinases of the silverleaf fungus, Chondrostereum purpureum, are metalloproteinases. Plant Pathol. 2010, 45, 552–563. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Jaros, A.; Yip, L.; Tran, L.; Kanellis, A.K.; Constabel, C.P. Evaluating ascorbate oxidase as a plant defense against leaf-chewing insects using transgenic poplar. J. Chem. Ecol. 2008, 34, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Garciapineda, E.; Castromercado, E.; Lozoyagloria, E. Gene expression and enzyme activity of pepper (Capsicum annuum L.) ascorbate oxidase during elicitor and wounding stress. Plant Sci. 2004, 166, 237–243. [Google Scholar] [CrossRef]
- Ananchanok, T.; Theingi, S.T.A.; Ashley, B.M.; Aine, L.P. Stress-responsive α-dioxygenase expression in tomato roots. J. Exp. Bot. 2005, 56, 713–723. [Google Scholar]
- Glazebrook, J.; Zook, M.; Mert, F.; Kagan, I.; Rogers, E.E.; Crute, I.R.; Holub, H.E.; Hammerschmidt, R.; Ausube, F.M. Phytoalexin-deficient mutants of Arabidopsis reveal that PAD4 encodes a regulatory factor and that four PAD genes contribute to downy mildew resistance. Genetics 1997, 146, 381–392. [Google Scholar] [PubMed]
- Rietz, S.; Stamm, A.; Malonek, S.; Wagner, S. Different roles of enhanced disease susceptibility1 (EDS1) bound to and dissociated from phytoalexin deficient4 (PAD4) in Arabidopsis immunity. New Phytol. 2011, 191, 107–119. [Google Scholar] [CrossRef]
- Hua, J.; Grisafi, P.; Cheng, S.H.; Fink, G.R. Plant growth homeostasis is controlled by the Arabidopsis BON1 and BAP1 genes. Genes Dev. 2001, 15, 2263–2272. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.J.; Li, Y.P.; Hua, J. The C2 domain protein BAP1 negatively regulates defense responses in Arabidopsis. Plant J. 2010, 48, 238–248. [Google Scholar] [CrossRef]
- Fry, S.C.; Smith, R.C.; Renwick, K.F.; Martin, D.J.; Hodge, S.K.; Matthews, K.J. Xyloglucan endotransglycosylase, a new wall-loosening enzyme activity from plants. Biochem J. 1992, 282, 821–828. [Google Scholar] [CrossRef] [Green Version]
- Bourquin, V.; Nishikubo, N.; Abe, H.; Brumer, H.; Denman, S.; Eklund, M.; Christiernin, M.; Teeri, T.T.; Sundberg, B.; Mellerowicz, E.J. Xyloglucan endotransglycosylases have a function during the formation of secondary cell walls of vascular tissues. Plant Cell 2002, 14, 3073–3088. [Google Scholar] [CrossRef] [Green Version]
- Bonello, P.; Gordon, T.R.; Herms, D.A.; Wood, D.L. Nature and ecological implications of pathogen-induced systemic resistance in conifers: A novel hypothesis. Physiol. Mol. Plant Pathol. 2006, 68, 95–104. [Google Scholar] [CrossRef]
- Yan, H.W.; Liu, S.R.; Xiang, Y. Identification and characterization analysis of specific genes of poplar. Mol. Plant Breed. 2018, 16, 123–129. [Google Scholar]
- Kenichiro, I. Cytochrome P450 enzymes in biosyntheses of some plant secondary metabolites. Yakugaku Zasshi 2005, 125, 31–49. [Google Scholar]
- Wang, X.M.; Lv, W.; Zhang, Z.; Chen, S.Q.; Liu, W.H.; Zhou, H. A preliminary study on the resistance of poplar to Anoplophora nobilis Ganglbauer. J. Agric. Sci. 1985, 23, 95–99. [Google Scholar]
- Li, D.M.; Zhou, L.Y. Coevolution— The relationship between insects and plants. Entomol. Knowl. 1997, 34, 45–49. [Google Scholar]
- Torikata, H.; Matsui, S. On the polyphenolic substances in the chestnut trees and the relation of the contents of them to the resistance to chestnut gall wasps. Engei Gakkai Zasshi 2007, 35, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Li, H.P.; Wang, Z.G.; Yang, M.S.; Zhang, Y.G.; Huang, D.Z.; Zhang, S.H. The relation between tannin and phenol constituents and resistance to Anoplophora glabripennis of various poplar tree species. J. Hebei Agric. Univ. 2003, 26, 36–39. [Google Scholar]
- Fang, J.; Zhao, B.G. Resistance of different clones of Populus deltoids to Clostera anastomosis (Lepidoptera: Notodontidae) larvae and the related mechanisms. Acta Entomol. Sin. 2011, 54, 1042–1050. [Google Scholar]
- Velikova, V.; Salerno, G.; Frati, F.; Peri, E.; Conti, E.; Colazza, S.; Loreto, F. Influence of feeding and oviposition by phytophagous pentatomids on photosynthesis of herbaceous plants. J. Chem. Ecol. 2010, 36, 629. [Google Scholar] [CrossRef]
- Baldwin, I.T. Jasmonate-induced responses are costly but benefit plants under attack in native populations. Proc. Natl. Acad. Sci. USA 1998, 95, 8113. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 108-O | 108-N | 108-C | 111-O | 111-N | 111-C | |
|---|---|---|---|---|---|---|
| Raw reads number | 50,255,389 | 52,672,759 | 52,355,816 | 51,052,837 | 49,999,297 | 48,190,009 |
| Clean reads number | 45,098,385 (89.74%) | 46,656,841 (88.58%) | 46,289,664 (88.43%) | 45,536,401 (89.20%) | 44,735,416 (89.50%) | 43,503,490 (90.28%) |
| Mapped reads | 28,247,431 (62.67%) | 29,579,017 (63.33%) | 29,966,665 (64.67%) | 28,427,433 (62.33%) | 28,158,929 (63.00%) | 28,140,356 (64.67%) |
| Unmapped reads | 16,850,954 (37.36%) | 17,077,825 (36.60%) | 16,322,999 (35.26%) | 17,108,969 (37.57%) | 16,576,487 (37.05%) | 15,363,134 (35.31%) |
| Multi map reads | 1,061,769 (2.67%) | 1,375,346 (3.00%) | 1,967,076 (4.33%) | 1,355,164 (3.00%) | 893,508 (2.00%) | 1,130,265 (3.00%) |
| 108-O | 108-N | 108-C | 111-O | 111-N | 111-C | |
|---|---|---|---|---|---|---|
| Exon | 13,453,196 (93.68%) | 13,482,984 (93.03%) | 13,358,075 (94.08%) | 13,327,758 (93.84%) | 12,615,311 (92.46%) | 13,056,880 (94.72%) |
| Intron | 581,278 (4.05%) | 648,371 (4.49%) | 514,929 (3.63%) | 552,302 (3.91%) | 700,806 (5.07%) | 443,171 (3.23%) |
| Intergenic | 326,553 (2.27%) | 360,234 (2.49%) | 514,929 (2.29%) | 320,559 (2.26%) | 337,833 (2.47%) | 282,035 (2.05%) |
| Gene Description | Gene Symbol | Gene ID | 108-O vs. 108-C | 108-N vs. 108-C | 108-O vs. 108-N | |||
|---|---|---|---|---|---|---|---|---|
| Ratio | P Value | Ratio | P Value | Ratio | P Value | |||
| Trypsin and protease inhibitor | TPIN | POPTR_0001s31740g | 52.77 | 0.0003 | 4.32 | 0.6394 | 12.24 | 0.0081 |
| NPR1/NIM1-interacting protein 2 | NIMIN-2 | POPTR_0002s19140g | 577.01 | 0.0000 | 33.21 | 0.0592 | 17.56 | 0.0457 |
| Mildew resistance protein, RPW8 homolog | HR4 | POPTR_0007s115001g | 3.99 | 0.0377 | 1.10 | 0.8886 | 3.65 | 0.0537 |
| Pathogen-responsive α-dioxygenase | POPTR_0012s04690g | 8.10 | 1.0000 | 95.76 | 0.0072 | 0.08 | 0.0066 | |
| Metalloproteinase | POPTR_0012s10640g | 9.19 | 0.0001 | 1.41 | 0.6673 | 6.58 | 0.0135 | |
| L-Ascorbate oxidase | POPTR_0005s08120g | 31.85 | 0.0000 | 0.57 | 0.7684 | 55.79 | 0.0000 | |
| Xyloglucan endotransglycosylase | XTR8 | POPTR_0001s01870g | 17.67 | 0.0010 | 1.69 | 0.6164 | 10.44 | 0.0322 |
| PSI reaction center subunit VI | POPTR_0003s05110g | 0.25 | 0.0604 | 0.42 | 0.0083 | 0.59 | 0.2488 | |
| PSII 5-kD protein | POPTR_0001s26410g | 0.17 | 0.0209 | 0.42 | 0.0072 | 0.41 | 0.0847 | |
| PSII core complex PsbY | PSBY | POPTR_0008s18230g | 0.30 | 0.0241 | 0.53 | 0.0412 | 0.57 | 0.1884 |
| PSII reaction center PsbP | POPTR_0010s21710g | 0.27 | 0.0272 | 0.35 | 0.0021 | 0.76 | 0.6710 | |
| PSII reaction center PsbW | POPTR_0002s25810g | 0.17 | 0.0029 | 0.48 | 0.1582 | 0.36 | 0.0892 | |
| PSII assembly factor HCF136 | HCF136 | POPTR_0007s07780g | 0.23 | 0.0038 | 0.33 | 0.0015 | 0.71 | 0.4393 |
| Divinyl protochlorophyllide a 8-vinyl reductase | PCB2 | POPTR_0008s20940g | 0.28 | 0.0332 | 0.54 | 0.0872 | 0.51 | 0.2402 |
| Chlorophyll a/b-binding protein | LHCB3 | POPTR_0002s22220g | 0.24 | 0.0780 | 0.42 | 0.0079 | 0.58 | 0.2528 |
| Oxygen evolving enhancer 3 (PsbQ) | POPTR_0010s17400g | 0.20 | 0.0004 | 0.42 | 0.0213 | 0.48 | 0.0944 | |
| Ferredoxin | POPTR_0007s09630g | 0.13 | 0.0197 | 0.31 | 0.0098 | 0.44 | 0.3389 | |
| Ferredoxin-thioredoxin reductase | POPTR_0001s34690g | 0.33 | 0.0442 | 0.45 | 0.0155 | 0.74 | 0.5784 | |
| Gene Description | Gene Symbol | Gene ID | 111-O vs. 111-C | 111-N vs. 111-C | 111-O vs. 111-N | |||
|---|---|---|---|---|---|---|---|---|
| Ratio | P Value | Ratio | P Value | Ratio | P Value | |||
| Hevein-like protein | PR4/HEL | POPTR_0013s03890g | 2.70 | 0.0274 | 1.20 | 0.5848 | 2.27 | 0.3236 |
| Phytoalexin-deficient 4 protein/lipase | PAD4 | POPTR_0005s06960g | 5.35 | 0.0026 | 1.76 | 0.4008 | 3.04 | 0.0031 |
| BON1-associated protein 1 | BAP1 | POPTR_0014s07470g | 10.39 | 0.0285 | 0.18 | 0.0154 | 58.56 | 0.0000 |
| Xyloglucan endotransglycosylase | XTR8 | POPTR_0001s01870g | 13.70 | 0.0000 | 5.40 | 0.0017 | 2.53 | 0.0016 |
| Cytochrome P450 | CYP83B1 | POPTR_0002s02700g | 1.48 | 0.4434 | 2.70 | 0.0175 | 0.54 | 0.1408 |
| PSI reaction center subunit VI | POPTR_0003s05110g | 0.19 | 0.0000 | 0.24 | 0.0000 | 0.79 | 0.3069 | |
| PSII 5-kD protein | POPTR_0001s26410g | 0.13 | 0.0000 | 0.21 | 0.0000 | 0.65 | 0.1129 | |
| PSII core complex PsbY | PSBY | POPTR_0008s18230g | 0.29 | 0.0017 | 0.29 | 0.0000 | 1.00 | 0.9721 |
| PSII reaction center PsbP | POPTR_0010s21710g | 0.22 | 0.0000 | 0.35 | 0.0079 | 0.63 | 0.1629 | |
| PSII reaction center PsbW | POPTR_0002s25810g | 0.27 | 0.0149 | 0.36 | 0.0000 | 0.76 | 0.3453 | |
| PSII assembly factor HCF136 | HCF136 | POPTR_0007s07780g | 0.22 | 0.0000 | 0.30 | 0.0000 | 0.75 | 0.2693 |
| Divinyl protochlorophyllide a 8-vinyl reductase | PCB2 | POPTR_0008s20940g | 0.24 | 0.0001 | 0.33 | 0.0000 | 0.71 | 0.2143 |
| Chlorophyll a/b-binding protein | LHCB3 | POPTR_0002s22220g | 0.12 | 0.0006 | 0.24 | 0.0000 | 0.50 | 0.0125 |
| Oxygen evolving enhancer 3 (PsbQ) | POPTR_0010s17400g | 0.29 | 0.0005 | 0.38 | 0.0000 | 0.76 | 0.2936 | |
| Cytochrome c biogenesis protein | POPTR_0009s11490g | 0.36 | 0.0074 | 0.32 | 0.0000 | 1.14 | 0.6612 | |
| Ferredoxin | POPTR_0007s09630g | 0.10 | 0.0000 | 0.21 | 0.0001 | 0.46 | 0.0499 | |
| Ferredoxin-thioredoxin reductase | POPTR_0001s34690g | 0.32 | 0.0014 | 0.37 | 0.0000 | 0.86 | 0.5290 | |
| Fructose-1,6-bisphosphatase | POPTR_0005s21360g | 0.36 | 0.0032 | 0.52 | 0.0041 | 0.68 | 0.1035 | |
| Starch synthase | POPTR_0011s15540g | 0.03 | 0.0001 | 0.15 | 0.0001 | 0.22 | 0.1285 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Zhang, S.; Liu, F.; Kong, X.; Zhang, Z. Effect of Micromelalopha sieversi (Staudinger) Oviposition Behavior on the Transcriptome of Two Populus Section Aigeiros Clones. Forests 2020, 11, 1021. https://doi.org/10.3390/f11091021
Guo L, Zhang S, Liu F, Kong X, Zhang Z. Effect of Micromelalopha sieversi (Staudinger) Oviposition Behavior on the Transcriptome of Two Populus Section Aigeiros Clones. Forests. 2020; 11(9):1021. https://doi.org/10.3390/f11091021
Chicago/Turabian StyleGuo, Li, Sufang Zhang, Fu Liu, Xiangbo Kong, and Zhen Zhang. 2020. "Effect of Micromelalopha sieversi (Staudinger) Oviposition Behavior on the Transcriptome of Two Populus Section Aigeiros Clones" Forests 11, no. 9: 1021. https://doi.org/10.3390/f11091021
APA StyleGuo, L., Zhang, S., Liu, F., Kong, X., & Zhang, Z. (2020). Effect of Micromelalopha sieversi (Staudinger) Oviposition Behavior on the Transcriptome of Two Populus Section Aigeiros Clones. Forests, 11(9), 1021. https://doi.org/10.3390/f11091021