Host Range of Hymenoscyphus fraxineus in Slovak Arboreta

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
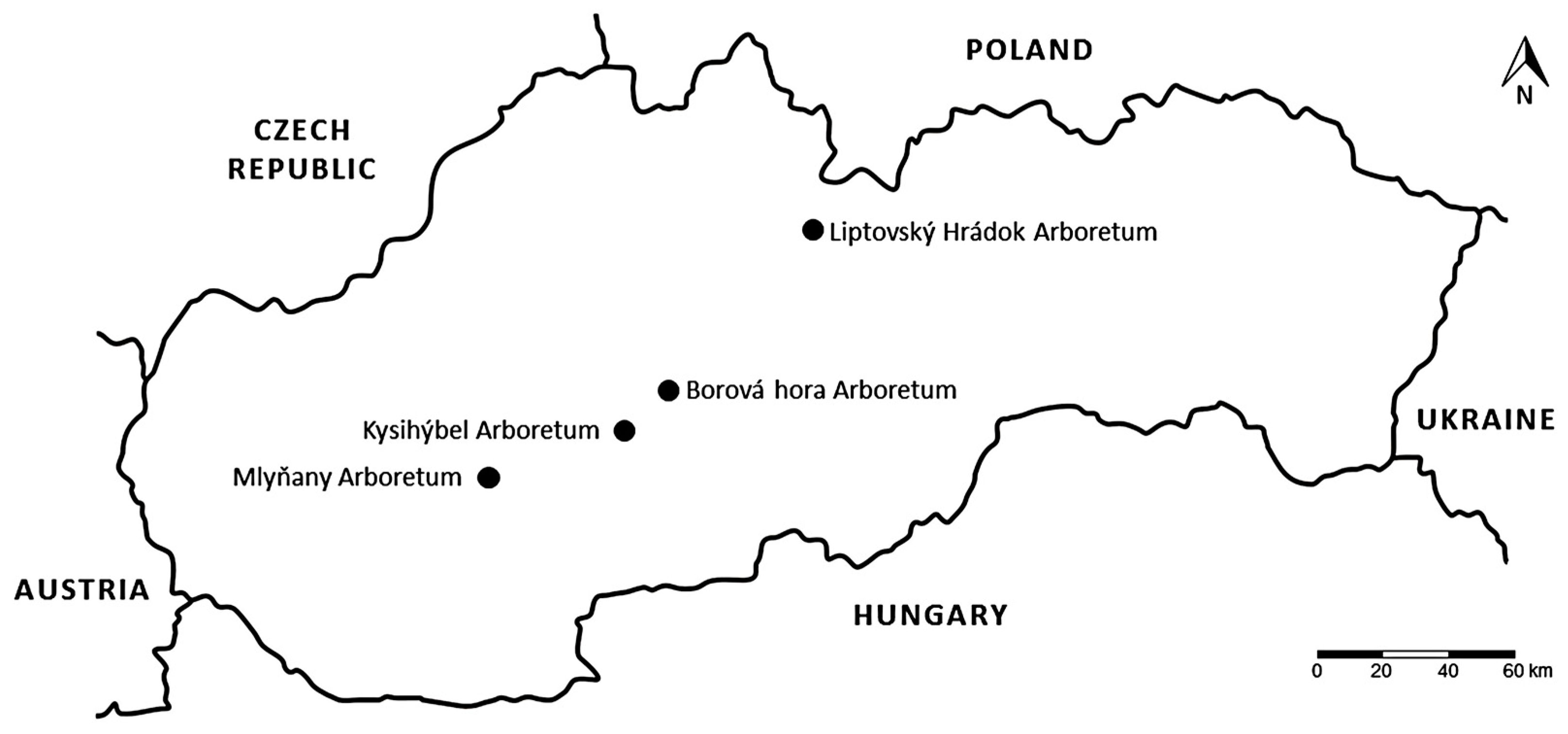
2.1. Study Sites
2.2. Ash Dieback Damage Assessment in Study Sites
2.3. Collection of Field Samples
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Phylogenetic Analysis
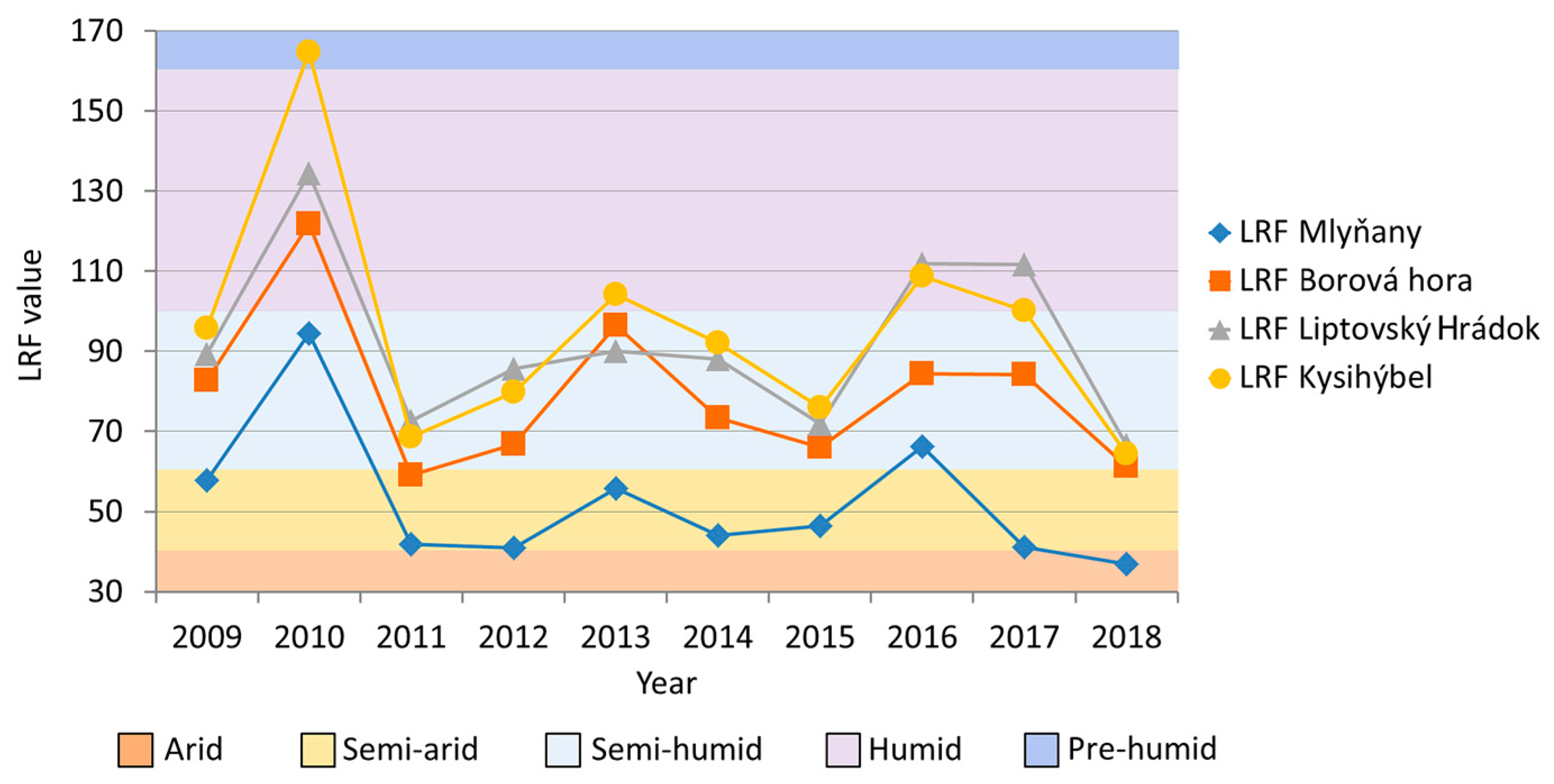
2.6. Statistical Analysis
3. Results
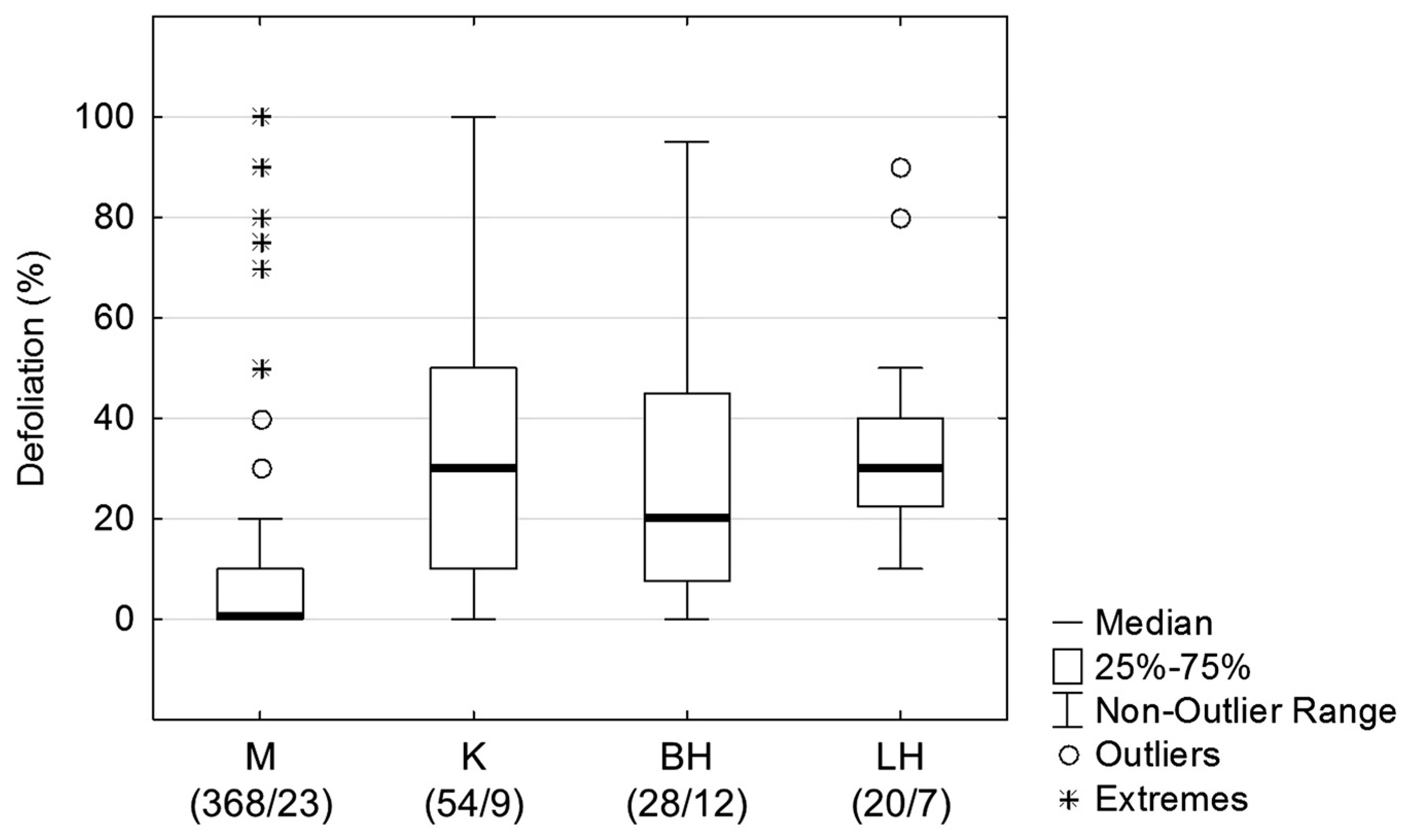
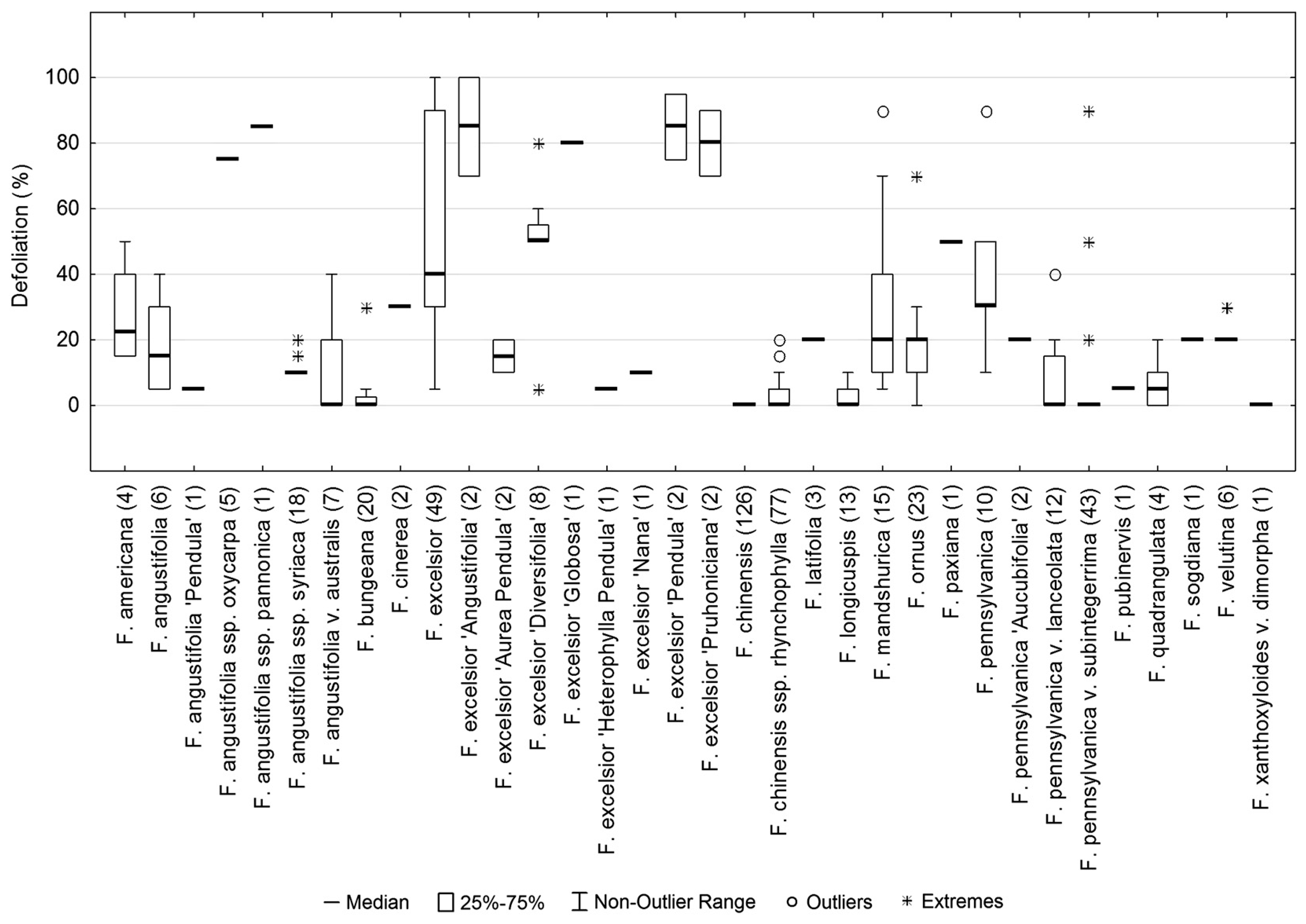
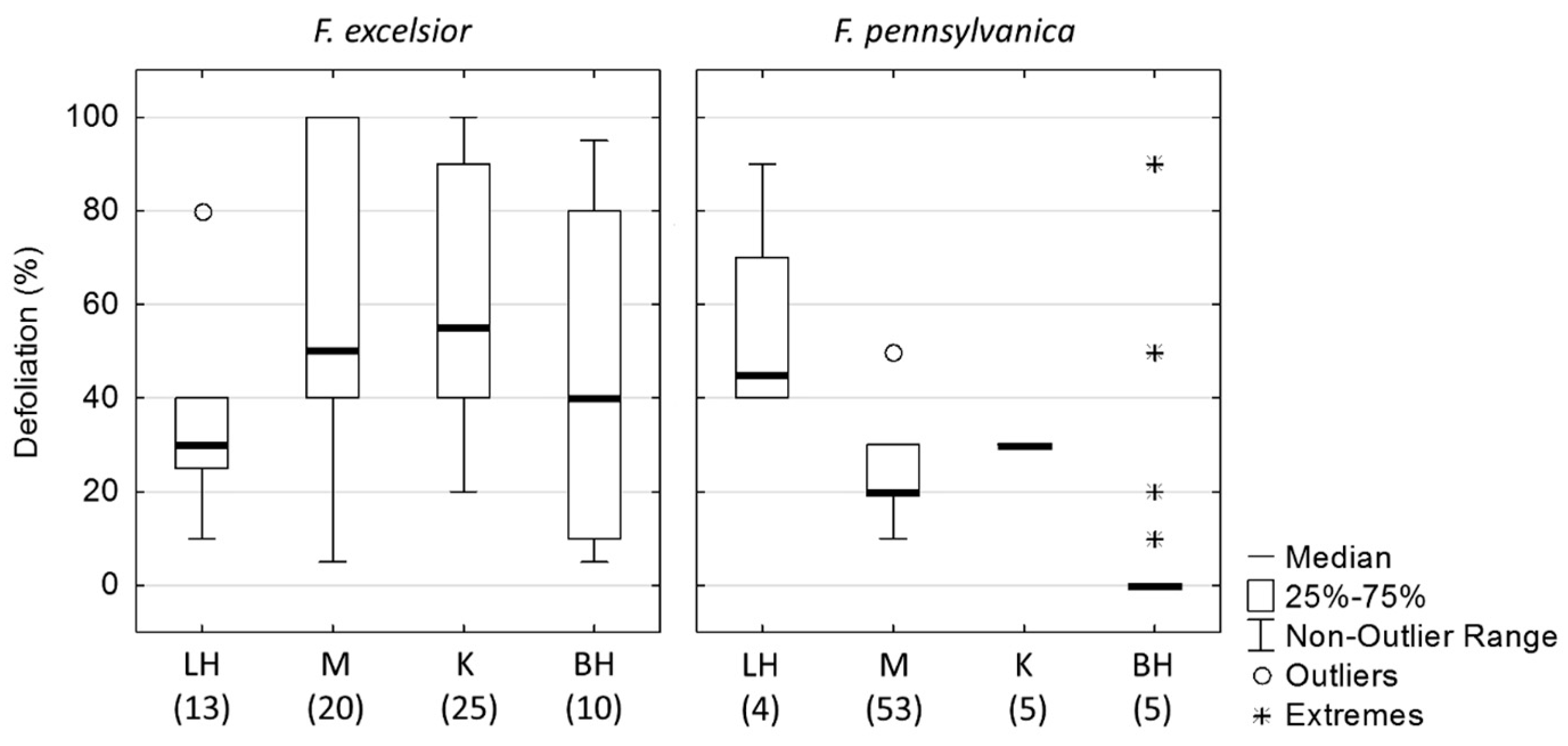
3.1. Defoliation Level of Ash Trees in Slovak Arboreta
3.2. Sequencing Analysis
3.3. Phylogenetic Analysis
3.4. Host Range of H. fraxineus in Slovak Arboreta
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wallander, E. Systematics and floral evolution in Fraxinus (Oleaceae). Belg. Dendrol. Belg. 2013, 2012, 38–58. [Google Scholar]
- Kowalski, T. Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Thomsen, I.M.; Stenlid, J. Occurrence and pathogenicity of fungi in necrotic and non-symptomatic shoots of declining common ash (Fraxinus excelsior) in Sweden. Eur. J. For. Res. 2009, 128, 51–60. [Google Scholar] [CrossRef]
- Timmermann, V.; Børja, I.; Hietala, A.M.; Kirisits, T.; Solheim, H. Ash dieback: Pathogen spread and diurnal patterns of ascospore dispersal, with special emphasis on Norway. Bull. OEPP 2011, 41, 14–20. [Google Scholar] [CrossRef]
- van Opstal, N.A. Introduction to the EPPO Workshop on Chalara fraxinea, a major threat for ash trees in Europe. Bull. OEPP 2011, 41, 1–2. [Google Scholar] [CrossRef]
- Queloz, V.; Grunig, C.R.; Berndt, R.; Kowalski, T.; Sieber, T.N.; Holdenrieder, O. Cryptic speciation in Hymenoscyphus albidus. For. Pathol. 2011, 41, 133–142. [Google Scholar] [CrossRef]
- Gross, A.; Han, J.G. Hymenoscyphus fraxineus and two new Hymenoscyphus species identified in Korea. Mycol. Prog. 2015, 14, 1–13. [Google Scholar] [CrossRef]
- Gross, A.; Hosoya, T.; Zhao, Y.J.; Baral, H.O. Hymenoscyphus linearis sp. nov: Another close relative of the ash dieback pathogen H. fraxineus. Mycol. Prog. 2015, 14, 1–15. [Google Scholar] [CrossRef]
- Kowalski, T.; Bilanski, P. Hymenoscyphus pusillus, a new species on leaves of Fraxinus pennsylvanica in Poland. For. Pathol. 2019, 49, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Drenkhan, R.; Solheim, H.; Bogacheva, A.; Riit, T.; Adamson, K.; Drenkhan, T.; Maaten, T.; Hietala, A.M. Hymenoscyphus fraxineus is a leaf pathogen of local Fraxinus species in the Russian Far East. Plant Pathol. 2017, 66, 490–500. [Google Scholar] [CrossRef]
- Thomas, P.A. Biological flora of the British Isles: Fraxinus excelsior. J. Ecol. 2016, 104, 1158–1209. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases. U.S. National Fungus Collections, ARS, USDA. Continuously Updated. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 9 May 2019).
- Drenkhan, R.; Hanso, M. New host species for Chalara fraxinea. New Dis. Rep. 2010, 22, 16. [Google Scholar] [CrossRef] [Green Version]
- Drenkhan, R.; Adamson, K.; Hanso, M. Fraxinus sogdiana, a Central Asian ash species, is susceptible to Hymenoscyphus fraxineus. Plant Protect. Sci. 2015, 51, 150–152. [Google Scholar] [CrossRef] [Green Version]
- Kirisits, T.; Schwanda, K. First definite report of natural infection of Fraxinus ornus by Hymenoscyphus fraxineus. For. Pathol. 2015, 45, 430–432. [Google Scholar] [CrossRef]
- Nielsen, L.R.; McKinney, L.V.; Hietala, A.M.; Kjær, E.D. The susceptibility of Asian, European and North American Fraxinus species to the ash dieback pathogen Hymenoscyphus fraxineus reflects their phylogenetic history. Eur. J. For. Res. 2017, 136, 59–73. [Google Scholar] [CrossRef] [Green Version]
- Department for Environment, Food & Rural Affairs (Defra). Ash Dieback Found on Three New Host Species of Tree in the UK. Available online: https://www.gov.uk/government/news/ash-dieback-found-on-three-new-host-species-of-tree-in-the-uk--2 (accessed on 9 August 2018).
- Kunca, A.; Brutovský, D.; Finďo, S.; Gubka, A.; Konôpka, B.; Konôpka, J.; Leontovyč, R.; Longauerová, V.; Minďáš, J.; Novotný, J.; et al. Výskyt Škodlivých Činiteľov v Lesoch Slovenska za rok 2005 a ich Prognóza na rok 2006, 1st ed.; National Forest Centre: Zvolen, Slovakia, 2006; 89p. [Google Scholar]
- Adamčíková, K.; Kádasi Horáková, M.; Jankovský, L.; Havrdová, L. Identification of Hymenoscyphus fraxineus, the causal agent of ash dieback in Slovakia. Biologia 2015, 70, 559–564. [Google Scholar] [CrossRef]
- Longauerová, V.; Maľová, M.; Adamčíková, K.; Kunca, A.; Kádasi Horáková, M.; Pastirčáková, K. Zdravotný stav jaseňov v semenných sadoch Slovenska v súvislosti s chradnutím vplyvom Chalara fraxinea (Hymenoscyphus fraxineus). In Proceedings of the Aktuálne Problémy v Ochrane Lesa 2015, Nový Smokovec, Slovakia, 29–30 January 2015; Kunca, A., Ed.; National Forest Centre: Zvolen, Slovakia, 2015; pp. 78–82. [Google Scholar]
- Kádasi Horáková, M.; Adamčíková, K.; Pastirčáková, K.; Longauerová, V.; Maľová, M. Natural infection of Fraxinus angustifolia by Hymenoscyphus fraxineus in Slovakia. Balt. For. 2017, 23, 52–55. [Google Scholar]
- Lukáčik, I.; Sarvašová, I. Arborétum Borová Hora 1965–2015, 1st ed.; Technical University in Zvolen: Zvolen, Slovakia, 2017; 131p. [Google Scholar]
- Jankovič, J. Lesnícke Arboretum Kysihýbel—Odborný Sprievodca; National Forest Centre—Forest Research Institute: Zvolen, Slovakia, 2015; 24p. [Google Scholar]
- Makara, O.; Meluš, J.; Pavlík, J. Sprievodca Zbierkami Chráneného Areálu Hrádocké Arboretum, 1st ed.; Secondary Forestry and Wood Technology School of Joseph Decret Matejovie in Liptovský Hrádok: Liptovský Hrádok, Slovakia, 2016; 56p. [Google Scholar]
- Hoťka, P.; Barta, M. Dreviny Arboréta Mlyňany SAV 2012, 1st ed.; Veda: Bratislava, Slovakia, 2012; 136p. [Google Scholar]
- Sobíšek, B. Meteorologický Slovník, Výkladový a Terminologický, 1st ed.; Academia: Prague, Czech Republic, 1993; 594p. [Google Scholar]
- Müller, E.; Stierlin, H.R. Sanasilva Tree Crown Photos with Percentages of Foliage Loss, 2nd ed.; Swiss Federal Institute for Forest, Snow and Landscape Research: Birmensdorf, Switzerland, 1990; 129p. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR protocols: A guide to methods and applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Johansson, S.B.K.; Vasaitis, R.; Ihrmark, K.; Barklund, P.; Stenlid, J. Detection of Chalara fraxinea from tissue of Fraxinus excelsior using species-specific ITS primers. For. Pathol. 2010, 40, 111–115. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetic analysis version 7.0 for bigger data sets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit, Biological Sequence Alignment Editor for Windows 95/98/NT/2000/XP/7, Version 7.2.5; Ibis Biosciences: Carlsbad, CA, USA. Available online: https://www.mybiosoftware.com/bioedit-7-0-9-biological-sequence-alignment-editor.html (accessed on 9 November 2019).
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Tropicos.org. Missouri Botanical Garden: St. Louis, MO, USA. Available online: http://www.tropicos.org/ (accessed on 10 February 2020).
- Koukol, O.; Haňáčková, Z.; Dvořák, M.; Havrdová, L. Unseen, but still present in Czechia: Hymenoscyphus albidus detected by real-time PCR, but not by intensive sampling. Mycol. Prog. 2016, 15, 6. [Google Scholar] [CrossRef]
- Kirisits, T.; Matlakova, M.; Mottinger-Kroupa, S.; Halmschlager, E.; Lakatos, F. Chalara fraxinea associated with dieback of narrow-leafed ash (Fraxinus angustifolia). Plant Pathol. 2010, 59, 411. [Google Scholar] [CrossRef]
- Vasaitis, R.; Enderle, R. Dieback of European Ash (Fraxinus spp.): Consequences and Guidelines for Sustainable Management; Report on European Cooperation in Science and Technology (COST) Action FP1103 FRAXBACK; SLU Service/Repro: Uppsala, Sweden, 2017; 300p. [Google Scholar]
- Hosoya, T.; Otani, Y.; Furuya, K. Materials for the fungus flora of Japan (46). Trans. Mycol. Soc. Jpn. 1993, 34, 429–432. [Google Scholar]
- Zhao, Y.J.; Hosoya, T.; Baral, H.O.; Hosaka, K.; Kakishima, M. Hymenoscyphus pseudoalbidus, the correct name for Lambertella albida reported from Japan. Mycotaxon 2012, 122, 25–41. [Google Scholar] [CrossRef]
- Zheng, H.D.; Zhuang, W.Y. Hymenoscyphus albidoides sp. nov. and H. pseudoalbidus from China. Mycol. Prog. 2014, 13, 625–638. [Google Scholar] [CrossRef]
- Baral, H.O.; Bemmann, M. Hymenoscyphus fraxineus vs. H. albidus—A comparative light microscopic study on the causal agent of European ash dieback and related foliicolous, stroma-forming species. Mycology 2014, 5, 228–290. [Google Scholar] [CrossRef] [Green Version]
- Han, J.G.; Shrestha, B.; Hosoya, T.; Lee, K.H.; Sung, G.H.; Shin, H.D. First report of the ash dieback pathogen Hymenoscyphus fraxineus in Korea. Mycobiology 2014, 42, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Cleary, M.; Nguyen, D.; Marčiulynienė, D.; Berlin, A.; Vasaitis, R.; Stenlid, J. Friend or foe? Biological and ecological traits of the European ash dieback pathogen Hymenoscyphus fraxineus in its native environment. Sci. Rep. 2016, 6, 21895. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, T.; Bilański, P.; Holdenrieder, O. Virulence of Hymenoscyphus albidus and H. fraxineus on Fraxinus excelsior and F. pennsylvanica. PLoS ONE 2015, 10, e0141592. [Google Scholar] [CrossRef] [Green Version]
- Gross, A.; Sieber, T. Virulence of Hymenoscyphus albidus and native and introduced Hymenoscyphus fraxineus on Fraxinus excelsior and Fraxinus pennsylvanica. Plant Pathol. 2016, 65, 655–663. [Google Scholar] [CrossRef]
- Kirisits, T.; Krumboeck, S.; Kraeutler, K. Nucleotide. Accession No. KP994902, Hymenoscyphus fraxineus voucher ST/FP/4-2 18S Ribosomal RNA Gene, Partial Sequence; Internal Transcribed Spacer 1, 5.8S Ribosomal RNA Gene, and Internal Transcribed Spacer 2, Complete Sequence; and 28S Ribosomal RNA Gene, Partial Sequence; National Library of Medicine, National Center for Biotechnology Information: Bethesda, MD, USA, 1988. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KP994902 (accessed on 21 February 2020).
- Kirisits, T.; Matlakova, M.; Mottinger-Kroupa, S.; Cech, T.L.; Halmschlager, E. The current situation of ash dieback caused by Chalara fraxinea in Austria. SDU Fac. For. J. Ser. A 2009, 97–119. [Google Scholar]
- Timmermann, V.; Nagy, N.E.; Hietala, A.M.; Børja, I.; Solheim, H. Progression of ash dieback in Norway related to tree age, disease history and regional aspects. Balt. For. 2017, 23, 150–158. [Google Scholar]
- Kessler, M.; Cech, T.L.; Brandstetter, M.; Kirisits, T. Dieback of ash (Fraxinus excelsior and Fraxinus angustifolia) in Eastern Austria: Disease development on monitoring plots from 2007 to 2010. J. Agric. Ext. Rural Dev. 2012, 4, 223–226. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Wilhelm, G.J.; Thomsen, I.M.; Metzler, B.; Kirisits, T.; Havrdová, L.; Enderle, R.; Dobrowolska, D.; Cleary, M.; Clark, J. Silvicultural strategies for Fraxinus excelsior in response to dieback caused by Hymenoscyphus fraxineus. Forestry 2017, 90, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Meluš, J.; Secondary Vocational School of Forestry and Wood Technology of Joseph Decret Matejovie, Liptovský Hrádok, Slovakia. Personal communication, 2018.
- Slávik, M.; Kysihýbel Arboretum, National Forest Centre, Zvolen, Slovakia. Personal communication, 2020.
- Schumacher, J. The general situation regarding ash dieback in Germany and investigations concerning the invasion and distribution strategies of Chalara fraxinea in woody tissue. Bull. OEPP 2011, 40, 7–10. [Google Scholar] [CrossRef]
- Kirisits, T.; Kritsch, P.; Kräutler, K.; Matlakova, M.; Halmschlager, E. Ash dieback associated with Hymenoscyphus pseudoalbidus in forest nurseries in Austria. J. Agric. Ext. Rural Dev. 2012, 4, 230–235. [Google Scholar] [CrossRef]
- Hietala, A.M.; Timmermann, V.; Børja, I.; Solheim, H. The invasive ash dieback pathogen Hymenoscyphus pseudoalbidus exerts maximal infection pressure prior to the onset of host leaf senescence. Fungal Ecol. 2013, 6, 302–308. [Google Scholar] [CrossRef]
- Havrdová, L. An analysis of selected environmental factors affecting the occurrence of Chalara fraxinea. Ph.D. Thesis, Czech University of Life Sciences, Prague, Czech Republic, 2015. [Google Scholar]
- Dvorak, M.; Rotkova, G.; Botella, L. Detection of airborne inoculum of Hymenoscyphus fraxineus and H. albidus during seasonal fluctuations associated with absence of apothecia. Forests 2016, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Ogris, N. Jesenov ožig, Chalara fraxinea. Novice Iz Varstva Gozdov 2008, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Hauptman, T.; Piškur, B.; de Groot, M.; Ogris, N.; Ferlan, M.; Jurc, D. Temperature effect on Chalara fraxinea: Heat treatment of saplings as a possible disease control method. For. Pathol. 2013, 43, 360–370. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on stems and twigs in initial and advanced stages of dieback of European ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Pastirčáková, K.; Ivanová, H.; Pastirčák, M. Species diversity of fungi on damaged branches and leaves of ashes (Fraxinus spp.) in different types of stands in Slovakia. Cent. Eur. For. J. 2018, 64, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Marçais, B.; Husson, C.; Godart, L.; Caël, O. Influence of site and stand factors on Hymenoscyphus fraxineus-induced basal lesion. Plant Pathol. 2016, 65, 1452–1461. [Google Scholar] [CrossRef]
- Kunca, A.; Leontovyč, R.; Zúbrik, M.; Gubka, A. Bark beetle outbreak on weakened ash trees and applied control measures. Bull. OEPP 2011, 41, 11–13. [Google Scholar] [CrossRef]
- Zúbrik, M.; Kunca, A.; Galko, J.; Vakula, J.; Leontovyč, R.; Gubka, A.; Nikolov, C.; Rell, S. Jaseň—Najčastejšie choroby a škodcovia jaseňa. Les & Letokruhy 2017, 7–8, 32–33. [Google Scholar]
- Diminić, D.; Kajba, D.; Milotić, M.; Andrić, I.; Kranjec, J. Susceptibility of Fraxinus angustifolia clones to Hymenoscyphus fraxineus in lowland Croatia. Balt. For. 2017, 23, 233–243. [Google Scholar]
- Adamčíková, K.; Pažitný, J.; Pastirčáková, K. Individual resistance of Fraxinus angustifolia and F. excelsior clones to Hymenoscyphus fraxineus. J. Plant Protect. Res. 2018, 58, 227–233. [Google Scholar]
- Stocks, J.J.; Metheringham, C.L.; Plumb, W.J.; Lee, S.J.; Kelly, L.J.; Nichols, R.A.; Buggs, R.J.A. Genomic basis of European ash tree resistance to ash dieback fungus. Nat. Ecol. Evol. 2019, 3, 1686–1696. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arboretum | Total Area [ha] | Coordinates | Altitude [m a.s.l.] | MAT [°C] | MTGS [°C] | APS [mm] | MPSGS [mm] |
|---|---|---|---|---|---|---|---|
| Borová hora | 47.80 | 48°35′ N 19°08′ E | 291–377 | 9.7 | 16.8 | 763.3 | 432.1 |
| Kysihýbel | 7.78 | 48°27′ N 18°56′ E | 530–563 | 9.1 | 15.8 | 853.3 | 465.8 |
| Liptovský Hrádok | 7.24 | 49°02′ N 19°43′ E | 641–646 | 8.0 | 14.6 | 729.5 | 460.2 |
| Mlyňany | 67.00 | 48°19′ N 18°22′ E | 163–217 | 10.7 | 17.5 | 555.5 | 337.0 |
| Host | Arboretum a | No. of Trees | Age of Trees b [yrs.] | Tree Defoliation c [%] | Year of Apothecia Collection | Testing Results of SSPP d | GenBank Acc. No. ITS |
|---|---|---|---|---|---|---|---|
| F. americana | K | 2 | 118 | 15 | n.f. | ||
| F. americana | M | 2 | 36 | 30; 50 | 2015 | – | |
| F. angustifolia | BH | 6 | 37 | 5–40 | 2018 | + | MK522151 |
| F. angustifolia ‘Pendula’ | M | 1 | 36 | 5 | n.f. | ||
| F. angustifolia ssp. oxycarpa | M | 5 | 36 | 75 | 2015 | + | MF175248 |
| F. angustifolia ssp. pannonica | BH | 1 | 43 | 85 | 2018 | + | MK491641 |
| F. angustifolia ssp. syriaca | M | 18 | 53 | 10–20 | n.f. | ||
| F. angustifolia v. australis | M | 7 | 33 | 0–40 | 2015 | + | MF175249 |
| F. bungeana | K | 4 | 29 | 5 | n.f. | ||
| F. bungeana | LH | 1 | 35 | 30 | 2018 | + | MK491649 |
| F. bungeana | M | 15 | 45 | 0 | n.f. | ||
| F. chinensis | M | 126 | 30–50 | 0–10 | n.f. | ||
| F. chinensis ssp. rhynchophylla | K | 6 | 29 | 5 | 2018 | + | MK491647 |
| F. chinensis ssp. rhynchophylla | LH | 1 | 20 | 10 | n.f. | ||
| F. chinensis ssp. rhynchophylla | M | 70 | 30–45 | 0–20 | n.f. | ||
| F. cinerea | K | 2 | 113 | 30 | 2018 | + | MK491646 |
| F. excelsior | K | 25 | 92–118 | 20–100 | 2018 | + | MK491642 |
| F. excelsior | LH | 12 | 45–80 | 10–40 | 2018 | + | MK491651 |
| F. excelsior | M | 12 | 51 | 5–100 | 2015 | + | MF175245 |
| F. excelsior ‘Angustifolia’ | M | 2 | 53 | 70; 100 | 2015 | + | MF175250 |
| F. excelsior ‘Aurea Pendula’ | BH | 2 | 35; 40 | 10; 20 | 2018 | + | MK491632 |
| F. excelsior ‘Diversifolia’ | BH | 3 | 44 | 5–80 | 2018 | + | MK491635 |
| F. excelsior ‘Diversifolia’ | M | 5 | 36 | 50 | 2015 | + | MF175252 |
| F. excelsior ‘Globosa’ | LH | 1 | 36 | 80 | 2018 | + | MK491652 |
| F. excelsior ‘Heterophylla Pendula’ | BH | 1 | 43 | 5 | 2018 | + | MK491639 |
| F. excelsior ‘Nana’ | BH | 1 | 39 | 10 | 2018 | + | MK491638 |
| F. excelsior ‘Pendula’ | BH | 1 | 39 | 95 | 2018 | + | MK491637 |
| F. excelsior ‘Pendula’ | M | 1 | 60 | 75 | 2015 | + | MF175251 |
| F. excelsior ‘Pruhoniciana’ | BH | 2 | 50 | 70; 90 | 2018 | + | MK491634 |
| F. latifolia * | K | 3 | 112 | 20 | 2018 | + | MK491645 |
| F. longicuspis | M | 13 | 35 | 0–10 | n.f. | ||
| F. mandshurica | BH | 3 | 53 | 40 | 2018 | + | MK491640 |
| F. mandshurica | K | 4 | 29 | 10–20 | 2018 | + | MK491643 |
| F. mandshurica | LH | 1 | 33 | 20 | 2018 | + | MK491648 |
| F. mandshurica | M | 7 | 30–50 | 5–90 | 2015 | + | MF175246 |
| F. ornus | BH | 3 | 55 | 0 | n.f. | ||
| F. ornus | K | 3 | 114 | 10 | n.f. | ||
| F. ornus | M | 17 | 48–57 | 20–70 | 2015 | + | MF175247 |
| F. paxiana | M | 1 | 53 | 50 | n.f. | ||
| F. pennsylvanica | BH | 3 | 55 | 10–50 | 2018 | + | MK491636 |
| F. pennsylvanica | K | 5 | 113 | 30 | 2018 | + | MK491644 |
| F. pennsylvanica | LH | 2 | 45 | 50; 90 | 2018 | + | MK491650 |
| F. pennsylvanica ‘Aucubifolia’ | BH | 2 | 45 | 20 | 2018 | + | MK491633 |
| F. pennsylvanica v. lanceolata | LH | 2 | 32 | 40 | 2018 | + | MK491653 |
| F. pennsylvanica v. lanceolata | M | 10 | 30–45 | 0-20 | 2016 | – | |
| F. pennsylvanica v. subintegerrima | M | 43 | 36 | 0–90 | 2016 | – | |
| F. pubinervis | M | 1 | 46 | 5 | n.f. | ||
| F. quadrangulata | M | 4 | 34 | 0–20 | 2016 | + | MF175253 |
| F. sogdiana ** | M | 1 | 34 | 20 | n.f. | ||
| F. velutina | M | 6 | 40 | 20–30 | n.f. | ||
| F. xanthoxyloides v. dimorpha | M | 1 | 33 | 0 | n.f. |
| Natural Distribution | Fraxinus Species | Number of Asymptomatic/Symptomatic Trees | % of Symptomatic Trees in all Arboreta | ||||
|---|---|---|---|---|---|---|---|
| BH | K | LH | M | All Arboreta | |||
| Asia | bungeana | 4/0 | 0/1 | 15/0 | 19/1 | 5 | |
| chinensis * | 3/3 | 1/0 | 196/0 | 200/3 | 1.5 | ||
| longicuspis | 13/0 | 13/0 | 0 | ||||
| mandshurica | 0/3 | 3/1 | 0/1 | 1/6 | 4/11 | 73.3 | |
| paxiana | 1/0 | 1/0 | 0 | ||||
| pubinervis | 1/0 | 1/0 | 0 | ||||
| sogdiana | 1/0 | 1/0 | 0 | ||||
| Europe to Asia | angustifolia * | 0/7 | 24/7 | 24/14 | 36.8 | ||
| excelsior * | 0/10 | 0/25 | 0/13 | 2/18 | 2/66 | 97.1 | |
| ornus | 3/0 | 3/0 | 14/3 | 20/3 | 13 | ||
| North America | americana | 2/0 | 2/0 | 4/0 | 0 | ||
| cinerea | 0/2 | 0/2 | 100 | ||||
| latifolia | 1/2 | 1/2 | 66.7 | ||||
| pennsylvanica * | 0/5 | 0/5 | 0/4 | 53/0 | 53/14 | 20.9 | |
| quadrangulata | 3/1 | 3/1 | 25 | ||||
| velutina | 6/0 | 6/0 | 0 | ||||
| North Africa | xanthoxyloides * | 1/0 | 1/0 | 0 | |||
| Totals | 3/25 | 16/38 | 1/19 | 333/35 | 353/117 | 24.9 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastirčáková, K.; Adamčíková, K.; Barta, M.; Pažitný, J.; Hoťka, P.; Sarvašová, I.; Kádasi Horáková, M. Host Range of Hymenoscyphus fraxineus in Slovak Arboreta. Forests 2020, 11, 596. https://doi.org/10.3390/f11050596
Pastirčáková K, Adamčíková K, Barta M, Pažitný J, Hoťka P, Sarvašová I, Kádasi Horáková M. Host Range of Hymenoscyphus fraxineus in Slovak Arboreta. Forests. 2020; 11(5):596. https://doi.org/10.3390/f11050596
Chicago/Turabian StylePastirčáková, Katarína, Katarína Adamčíková, Marek Barta, Jozef Pažitný, Peter Hoťka, Ivana Sarvašová, and Miriam Kádasi Horáková. 2020. "Host Range of Hymenoscyphus fraxineus in Slovak Arboreta" Forests 11, no. 5: 596. https://doi.org/10.3390/f11050596
APA StylePastirčáková, K., Adamčíková, K., Barta, M., Pažitný, J., Hoťka, P., Sarvašová, I., & Kádasi Horáková, M. (2020). Host Range of Hymenoscyphus fraxineus in Slovak Arboreta. Forests, 11(5), 596. https://doi.org/10.3390/f11050596