Cinara splendens (Hemiptera: Aphididae: Lachninae)—First Record in Palaearctic Region

Abstract
:1. Introduction
2. Materials and Methods
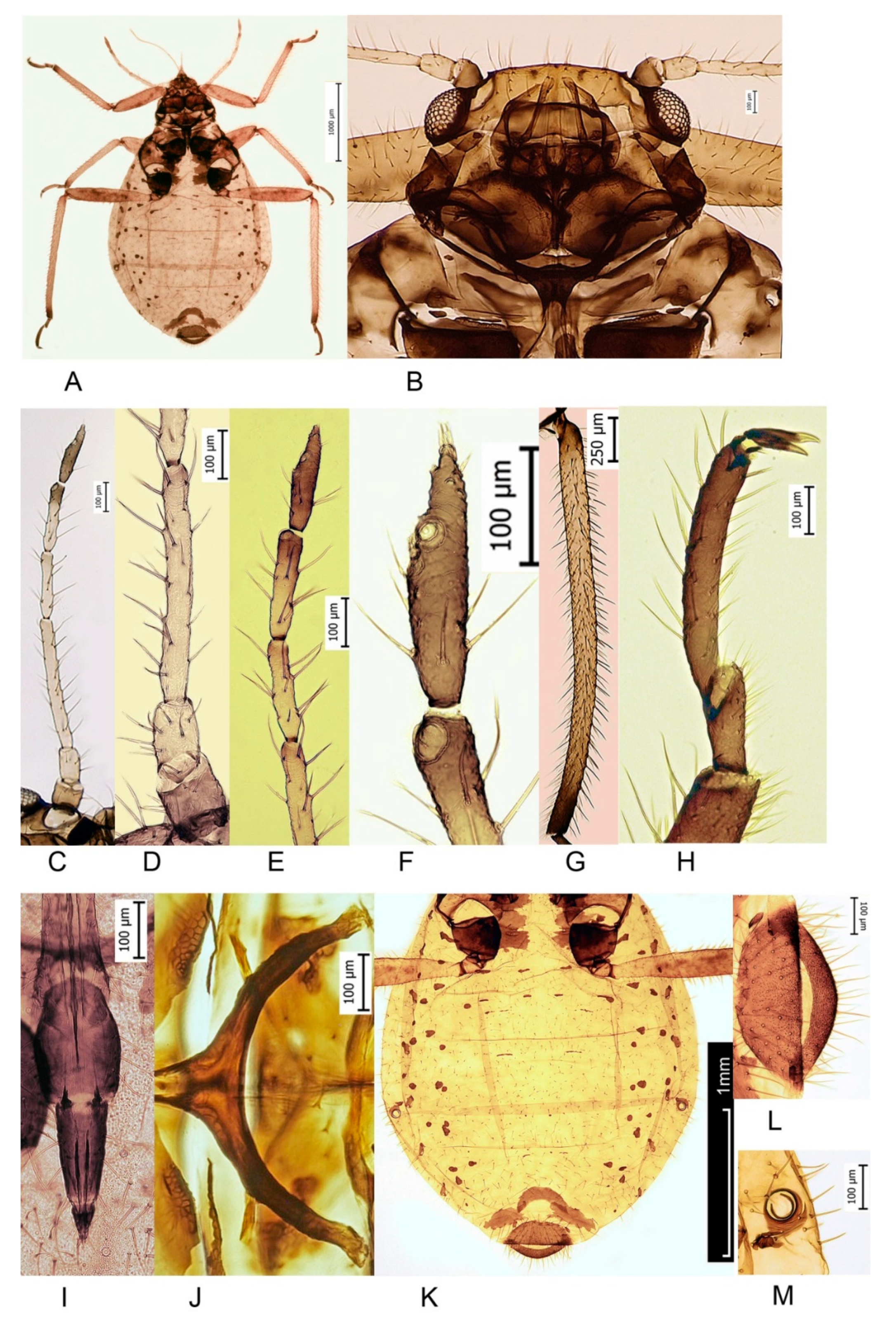
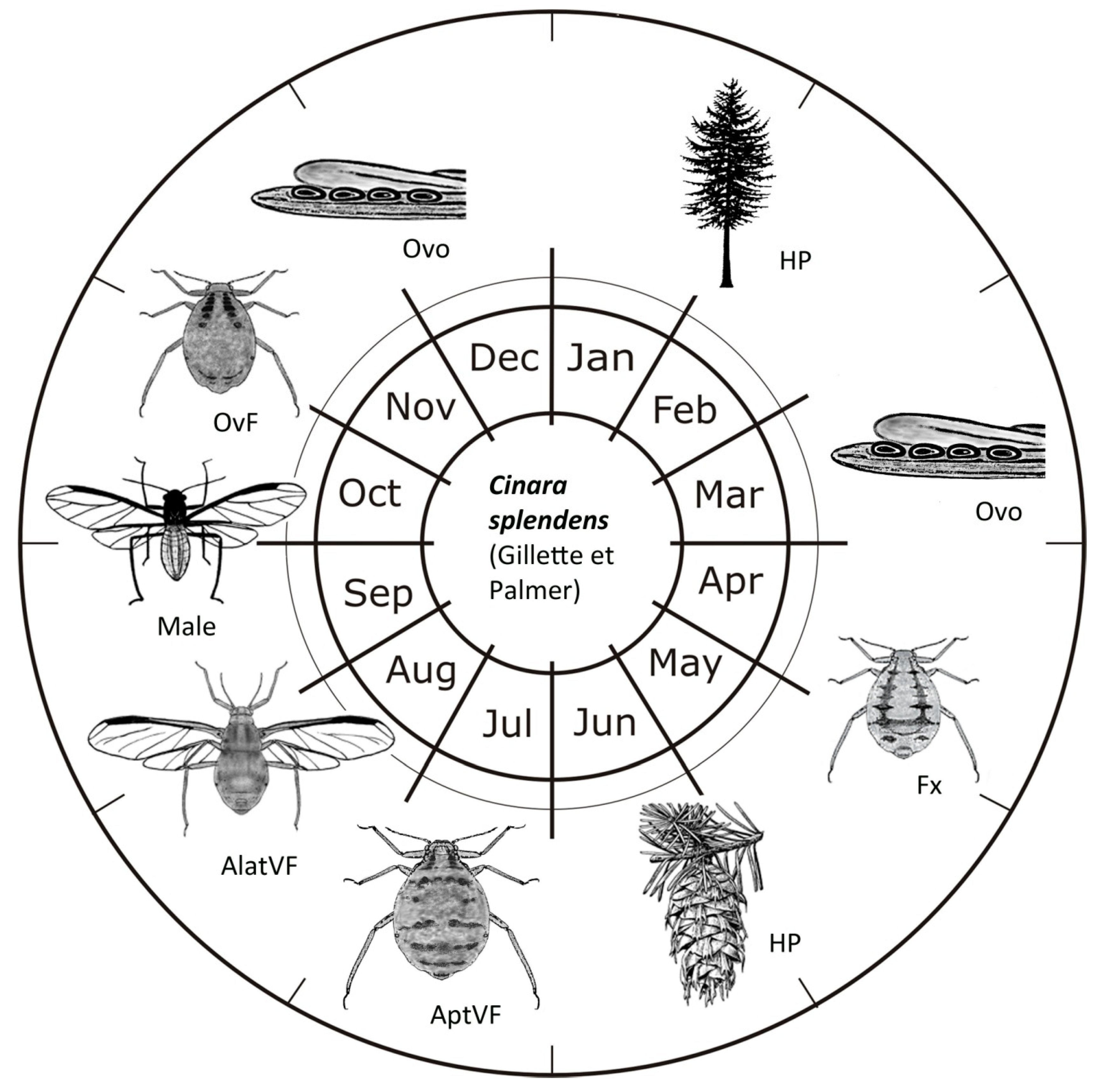
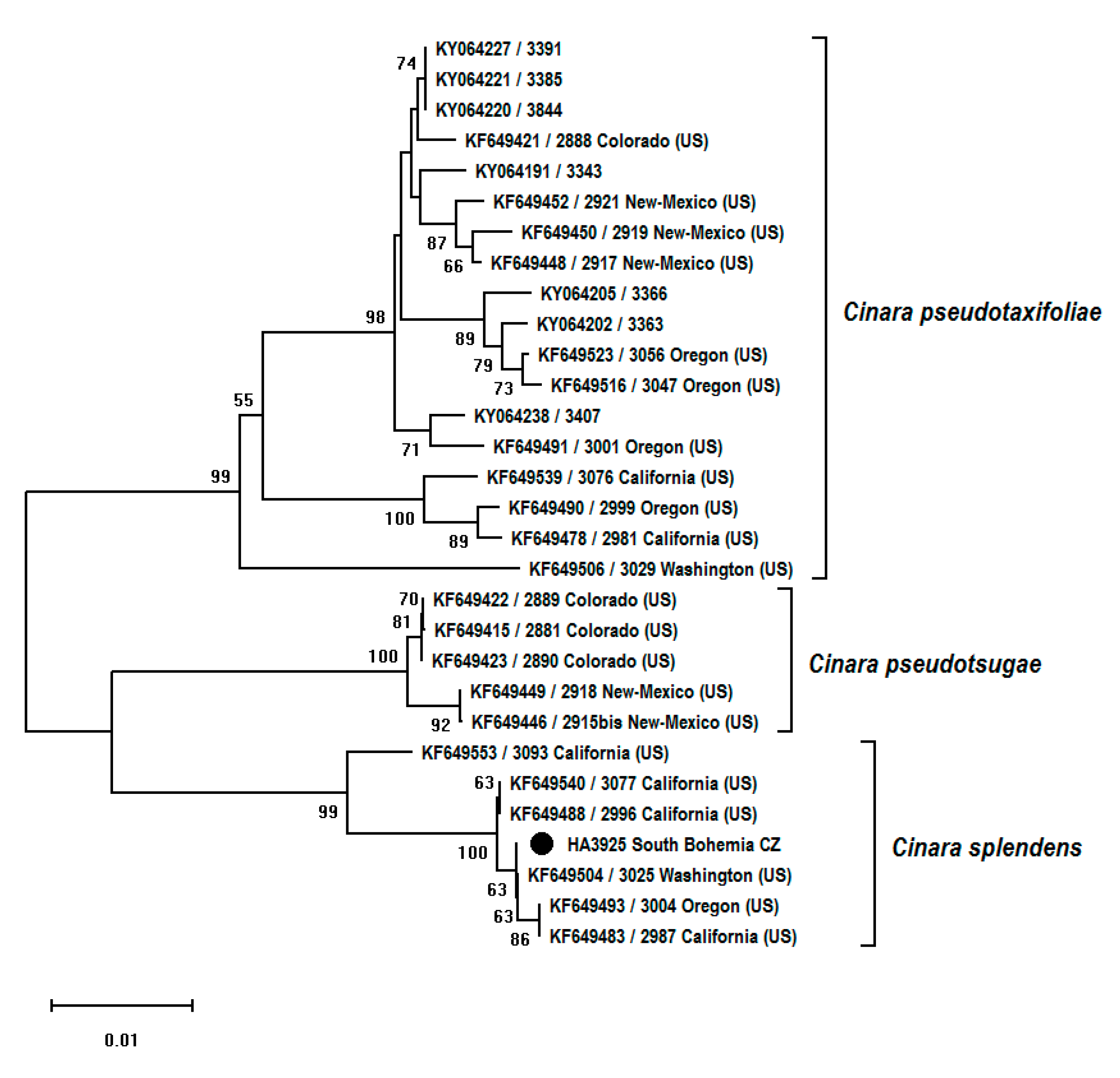
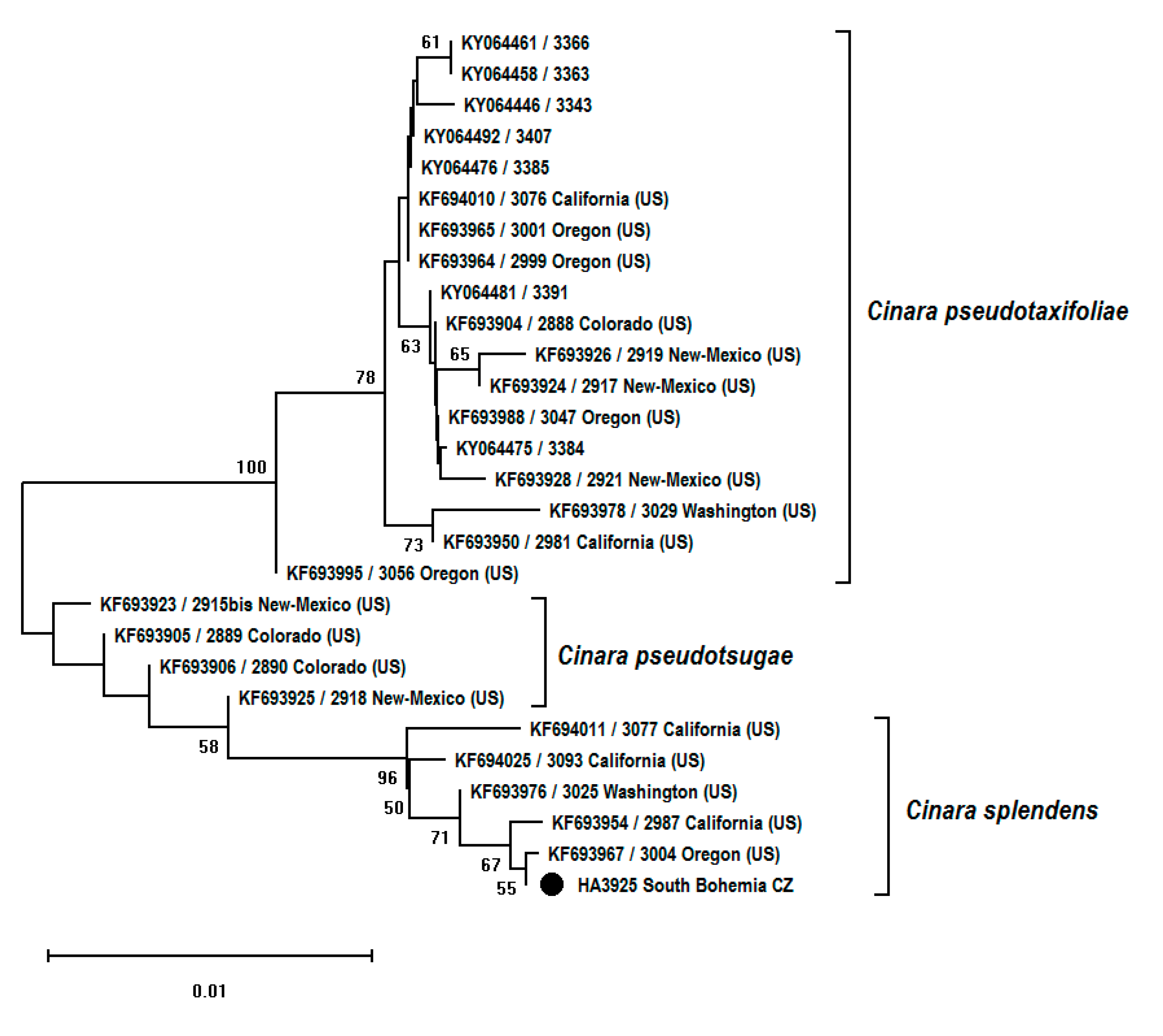
3. Results
- 1.
- Siphunculi pores on broad pigmented conical bases 0.4 mm or more in diameter. (Antennal hairs numerous, 0.08 mm or more long; hind tibial hairs 0.10–0.15 mm long). – Cinara (Cinara) commatula.
- –
- Siphunculi cones very small, with pigmented area less than 0.2 mm in diameter. – 2
- 2.
- All tibiae almost wholly pale, except that hind tibiae have contrastingly black apices. – Cinara (Cinara) splendens.
- –
- At least hind tibia dark or shading to dark over distal third or more of length. – 3
- 3.
- Longest abdominal hairs more than 0.10 mm long, longer than hairs on dorsal side of hind tibia, which are about 0.08 mm long. – Cinara (Cinara) vagabunda.
- –
- Longest abdominal hairs less than 0.10 mm long or, if longer, then they are shorter than longest hairs on dorsal side of hind tibia. – 4
- 4.
- Longest hairs on dorsal side of hind tibia 0.09–0.16 mm. length of sclerotised part of stylet groove 0.8–1.0 mm. – Cinara (Cinara) pseudotsugae.
- –
- Longest hairs on dorsal side of hind tibia 0.050–0.075 mm. length of sclerotised part of stylet groove 1.1–1.5 mm. – Cinara (Cinara) pseudotaxifoliae.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mlíkovský, J.; Stýblo, P. Nepůvodní druhy fauny a flóry ČR; ČSOP: Praha, Czech Republic, 2006; 495p. [Google Scholar]
- Petrović-Obradović, O.; Tomanović, Ž.; Poljaković-Pajnik, L.; Vučetić, A. An invasive species of aphid, Prociphilus fraxinifolii (Hemiptera, Aphididae, Eriosomatinae), found in Serbia. Arch. Biol. Sci. Belgrade 2007, 59, 9–10. [Google Scholar] [CrossRef]
- Petrović-Obradović, O.; Tomanović, Ž.; Poljaković-Pajnik, L.; Hrnčić, S.; Vučetić, A.; Radonjić, S. New invasive species of aphids (Hemiptera, Aphididae) in Serbia and Montenegro. Arch. Biol. Sci., Belgrade 2010, 62, 775–780. [Google Scholar] [CrossRef]
- Junkiert, Ł.; Wieczorek, K.; Wojciechowski, W. Periphyllus californiensis Shinji, 1917 (Hemiptera: Aphidoidea)—An invasive aphid species new to Poland. Pol. J. Entomol. 2011, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Hałaj, R.; Osiadacz, B. On foreign land: The conquest of Europe by Cinara curvipes (Patch, 1912). Dt. Entomol. Z. 2015, 62, 261–265. [Google Scholar] [CrossRef]
- Kanturski, M.; Lee, Y.; Depa, Ł. New records of an alien aphid species Tinocallis (Sappocallis) takachihoensis from countries in central and northern Europe (Hemiptera, Aphididae, Calaphidinae). ZooKeys 2018, 730, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilov, J.; Havelka, J.; Rakauskas, R. New for Lithuania aphid species of the tribe Eulachnini (Hemiptera: Aphididae, Lachninae): Is there any threat to local coniferous forests? Balt. For. 2019, 25, 25–31. [Google Scholar] [CrossRef]
- Wieczorek, K.; Chłond, D. Description of the previously unknown sexual morphs of Eucarazzia elegans from Iran and Pakistan and the northernmost record of viviparous generation from Europe. Bull. Insectol. 2019, 72, 177–186. [Google Scholar]
- Brandl, H. Erganzende untersuchungen zur ertragslage der baumarten fichte, kiefer, buche und eiche in Baden-Württemberg. Allg. Forst Jagdztg. 1989, 160, 91–99. [Google Scholar]
- Heidingsfelder, A.; Knoke, T. Douglasie versus Fichte; J.D. Sauerländer’s Verlag: Frankfurt, Germany, 2004. [Google Scholar]
- Hanewinkel, M. Klimawandel: Arealverschiebung von Hauptbaumarten in Südwest Deutschland. Available online: http://www.waldwissen.net (accessed on 8 February 2010).
- Schwappach, A. Denkschrift betreffend die Ergebnisse der in den Jahren 1981 bis 1890 in den PreuBischen Staatsforsten ausgef Uhrten Anbauversuchen mit fremdlandischen Holzarten. Z. Forst Jagdwes. 1891, 23, 18–34, 81–102, 148–164. [Google Scholar]
- Kandler, G.; Schmidt, M.; Breidenbach, J. Der Wald in Baden-Württemberg-Ergebnisse der Bundeswaldinventur 2. Available online: www.fva-bw.de/forschunglindex2.html (accessed on 8 February 2010).
- Podrázský, V.; Kubeček, J.; Čermák, R.; Štefančík, I. Zhodnocení dosavadního výzkumu douglasky tisolisté v České republice–přehled. In Proceedings of the Central European Silviculture, Kostelec nad Černými lesy, Czech Republic, 2–3 July 2013; Baláš, M., Podrázský, V., Kučeravá, B., Eds.; ČZU: Praha, Czech Republic, 2013; pp. 192–203. [Google Scholar]
- Palmer, M.A. Aphids of the Rocky Mountain Region; Thomas Say Foundation; Hirschfeld Press: Denver, Colorado, 1952; Volume 5, 452p. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Trees: An Identification and Information Guide; CAB International: Oxon, UK, 1994. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vriehoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- von Dohlen, C.D.; Kurosu, U.; Aoki, S. Phylogenetics and evolution of the eastern Asian eastern North American disjunct aphid tribe, Hormaphidini (Hemiptera: Aphididae). Mol. Phylogenet. Evol. 2002, 23, 257–267. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; STecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jousselin, E.; Cruad, A.; Genson, G.; Chevenet, F.; Fottitt, R.G.; Coeur d’Acier, A. Is ecological speciation a major trend in aphids? Insights from a molecular phylogeny of the conifer-feeding genus Cinara. Front. Zool. 2013, 10, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseguer, A.; Manzano-Marin, A.; Coeur d’Acier, A.; Clamens, A.-L.; Godefroid, M.; Jousselin, E. Buchnera has changed flatmate but the repeated replacement of co-obligate symbionts is not associated with the ecological expansions of their aphid hosts. Mol. Ecol. 2017, 26, 2363–2378. [Google Scholar] [CrossRef] [PubMed]
- Smith, J. Differential growth of roots and shoots of Douglas-fir (Pseudotsuga menziesii) seedlings infested with Cinara pseudotsugae, and population dynamics of a parasitoid wasp (Pauesia sp.) of C. splendens. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1997. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Sample | Locality | GPS | Date | Aphid Morph |
|---|---|---|---|---|
| 09HA3366 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2009-04-25 | fx, aptvf |
| 09HA3367 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2009-04-25 | fx, aptvf |
| 09HA3369 | Č. Budějovice, J. Opletala str. | 48°58’51.3” N 14°26’44.1” E | 2009-04-25 | fx, aptvf |
| 09HA3377 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-04-27 | fx, aptvf |
| 09HA3376 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-04-27 | fx, aptvf |
| 09HA3501 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-05-25 | aptvf |
| 09HA3502 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-05-25 | aptvf |
| 09HA3503 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-05-25 | aptvf |
| 09HA3504 | Praha 6, Břevnov park | 50°05’05.4” N 14°22’03.4” E | 2009-05-30 | aptvf |
| 09HA3546 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-06-15 | aptvf |
| 09HA3989 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2009-10-07 | aptvf |
| 09HA4027 | Č. Budějovice, Mariánské square | 48°58’46.5” N 14°28’22.0” E | 2009-11-10 | male, ovipara |
| 09HA4043 | Č. Budějovice, Mariánské square | 48°58’46.5” N 14°28’22.0” E | 2009-11-10 | male, ovipara |
| 11HA4255 | Č. Budějovice, Mariánské square | 48°58’46.5” N 14°28’22.0” E | 2011-04-28 | fx, aptvf |
| 11HA4256 | Č. Budějovice, Mariánské square | 48°58’46.5” N 14°28’22.0” E | 2011-04-28 | fx, aptvf |
| 12HA4549 | Stráž nad Nežárkou | 49°04’10.7” N 14°54’08.1” E | 2012-06-20 | aptvf |
| 13HA4682 | Č. Budějovice, Stromovka-park | 48°58’09.4” N 14°27’17.7” E | 2013-10-10 | aptvf |
| 15HA4669 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 15HA4670 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 15HA4671 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 15HA4775 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 15HA4781 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 15HA4782 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 15HA4783 | Č. Budějovice, Destinové str. | 48°58’49.9” N 14°26’57.0” E | 2015-05-25 | aptvf, alvf |
| 16HA5106 | Č. Budějovice, Stromovka park | 48°58’09.4” N 14°27’17.7” E | 2016-11-11 | male |
| 16HA5107 | Č. Budějovice, Stromovka park | 48°58’09.4” N 14°27’17.7” E | 2016-11-11 | male |
| 16HA5108 | Č. Budějovice, Stromovka-park | 48°58’09.4” N 14°27’17.7” E | 2016-11-11 | ovipara |
| 16HA5109 | Č. Budějovice, Stromovka park | 48°58’09.4” N 14°27’17.7” E | 2016-11-11 | ovipara |
| 16HA5110 | Č. Budějovice, Stromovka park | 48°58’09.4” N 14°27’17.7” E | 2016-11-11 | ovipara |
| 16HA5110 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2016-06-16 | aptvf, alvf |
| 19HA5939 | Č. Budějovice, Stromovka park | 48°58’09.4” N 14°27’17.7” E | 2019-01-25 | eggs |
| 19HA5445 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2019-06-06 | aptvf, alvf |
| 19HA5448 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2019-06-06 | aptvf, alvf |
| 19HA5450 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2019-06-06 | aptvf, alvf |
| 19HA5451 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2019-06-06 | aptvf, alvf |
| 19HA5452 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2019-06-06 | aptvf, alvf |
| 19HA5454 | Č. Budějovice, Šumava distr. | 48°58’89.3” N 14°07’66.4” E | 2019-06-06 | aptvf, alvf |
| Voucher [Reference] | Host Species | Location | GenBank Accession no. COI Fragment | GenBank Accession no. EF-1α Fragment |
|---|---|---|---|---|
| Cinara pseudotaxifoliae | ||||
| 2888 [21] | Pseudotsuga sp. | Colorado (US) | KF649421 | KF693904 |
| 2917 [21] | Pseudotsuga sp. | New-Mexico (US) | KF649448 | KF693924 |
| 2919 [21] | Pseudotsuga sp. | New-Mexico (US) | KF649450 | KF693926 |
| 2981 [21] | Pseudotsuga sp. | California (US) | KF649478 | KF693950 |
| 2999 [21] | Pseudotsuga menziesii | Oregon (US) | KF649490 | KF693964 |
| 3001 [21] | Pseudotsuga menziesii | Oregon (US) | KF649491 | KF693965 |
| 3029 [21] | Pseudotsuga sp. | Washington (US) | KF649506 | KF693978 |
| 3047 [21] | Pseudotsuga sp. | Oregon (US) | KF649516 | KF693988 |
| 3056 [21] | Pseudotsuga sp. | Oregon (US) | KF649523 | KF693995 |
| 3076 [21] | Pseudotsuga sp. | California (US) | KF649539 | KF694010 |
| 2921 [21] | Abies sp. | New-Mexico (US) | KF649452 | KF693928 |
| 3407 [22] | Pseudotsuga menziesii | – | KY064238 | KY064492 |
| 3391 [22] | Pseudotsuga menziesii | – | KY064227 | KY064481 |
| 3385 [22] | Pseudotsuga menziesii | – | KY064221 | KY064476 |
| 3384 [22] | Pseudotsuga menziesii | – | KY064220 | KY064475 |
| 3366 [22] | Pseudotsuga menziesii | – | KY064205 | KY064461 |
| 3363 [22] | Pseudotsuga menziesii | – | KY064202 | KY064458 |
| 3343 [22] | Pseudotsuga menziesii | – | KY064191 | KY064446 |
| Cinara pseudotsugae | ||||
| 2881 [21] | Pseudotsuga sp. | Colorado (US) | KF649415 | – |
| 2889 [21] | Pseudotsuga sp. | Colorado (US) | KF649422 | KF693905 |
| 2918 [21] | Pseudotsuga sp. | New-Mexico (US) | KF649449 | KF693925 |
| 2915bis [21] | Pseudotsuga sp. | New-Mexico (US) | KF649446 | KF693923 |
| 2890 [21] | Pseudotsuga sp. | Colorado (US) | KF649423 | KF693906 |
| Cinara splendens | ||||
| 2987 [21] | Pseudotsuga sp. | California (US) | KF649483 | KF693954 |
| 2996 [21] | Pseudotsuga menziesii | California (US) | KF649488 | – |
| 3004 [21] | Pseudotsuga sp. | Oregon (US) | KF649493 | KF693967 |
| 3025 [21] | Pseudotsuga macrocarpa | Washington (US) | KF649504 | KF693976 |
| 3077 [21] | Pseudotsuga sp. | California (US) | KF649540 | KF694011 |
| 3093 [21] | Pseudotsuga sp. | California (US) | KF649553 | KF694025 |
| HA3925 | Pseudotsuga menziesii | Czech Republic | MT708506 | MT708504 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havelka, J.; Havelka, J.; Starý, P. Cinara splendens (Hemiptera: Aphididae: Lachninae)—First Record in Palaearctic Region. Forests 2020, 11, 911. https://doi.org/10.3390/f11090911
Havelka J, Havelka J, Starý P. Cinara splendens (Hemiptera: Aphididae: Lachninae)—First Record in Palaearctic Region. Forests. 2020; 11(9):911. https://doi.org/10.3390/f11090911
Chicago/Turabian StyleHavelka, Jan, Jekaterina Havelka, and Petr Starý. 2020. "Cinara splendens (Hemiptera: Aphididae: Lachninae)—First Record in Palaearctic Region" Forests 11, no. 9: 911. https://doi.org/10.3390/f11090911
APA StyleHavelka, J., Havelka, J., & Starý, P. (2020). Cinara splendens (Hemiptera: Aphididae: Lachninae)—First Record in Palaearctic Region. Forests, 11(9), 911. https://doi.org/10.3390/f11090911