The Effect of Eutypella parasitica on the Wood Decay of Three Maple Species




Abstract
:1. Introduction
2. Materials and Methods
2.1. Decay Test
2.2. Light and Scanning Electron Microscopy
3. Results
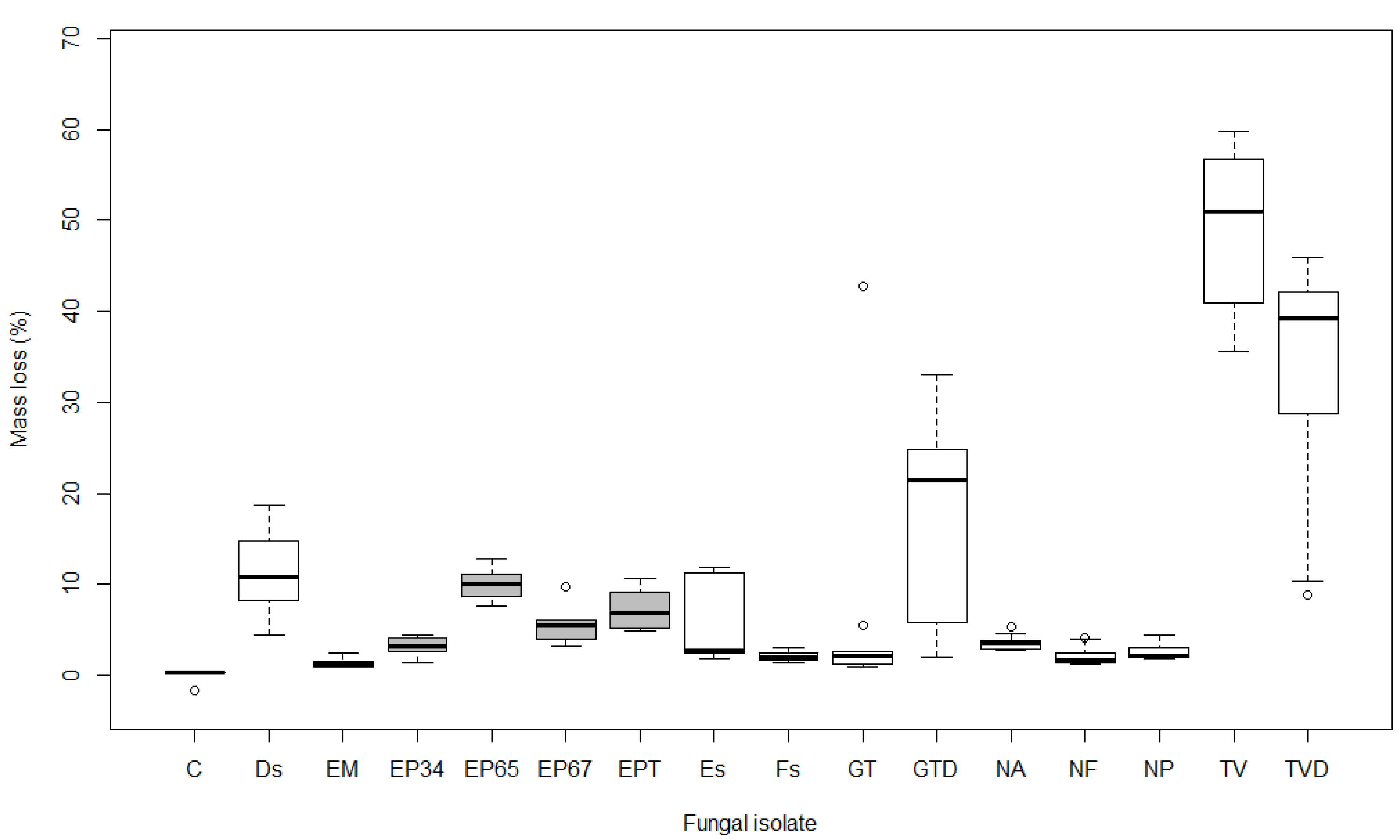
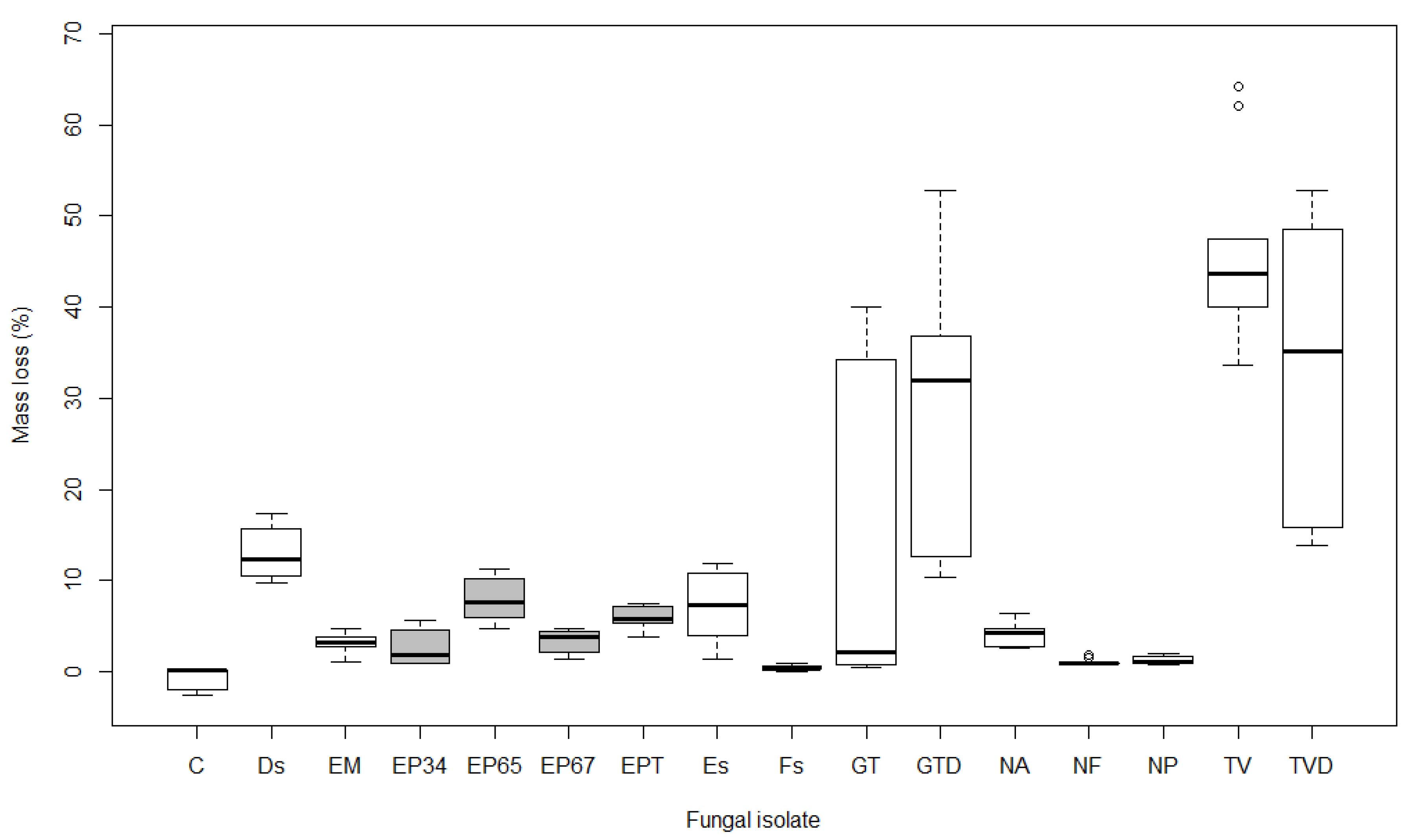
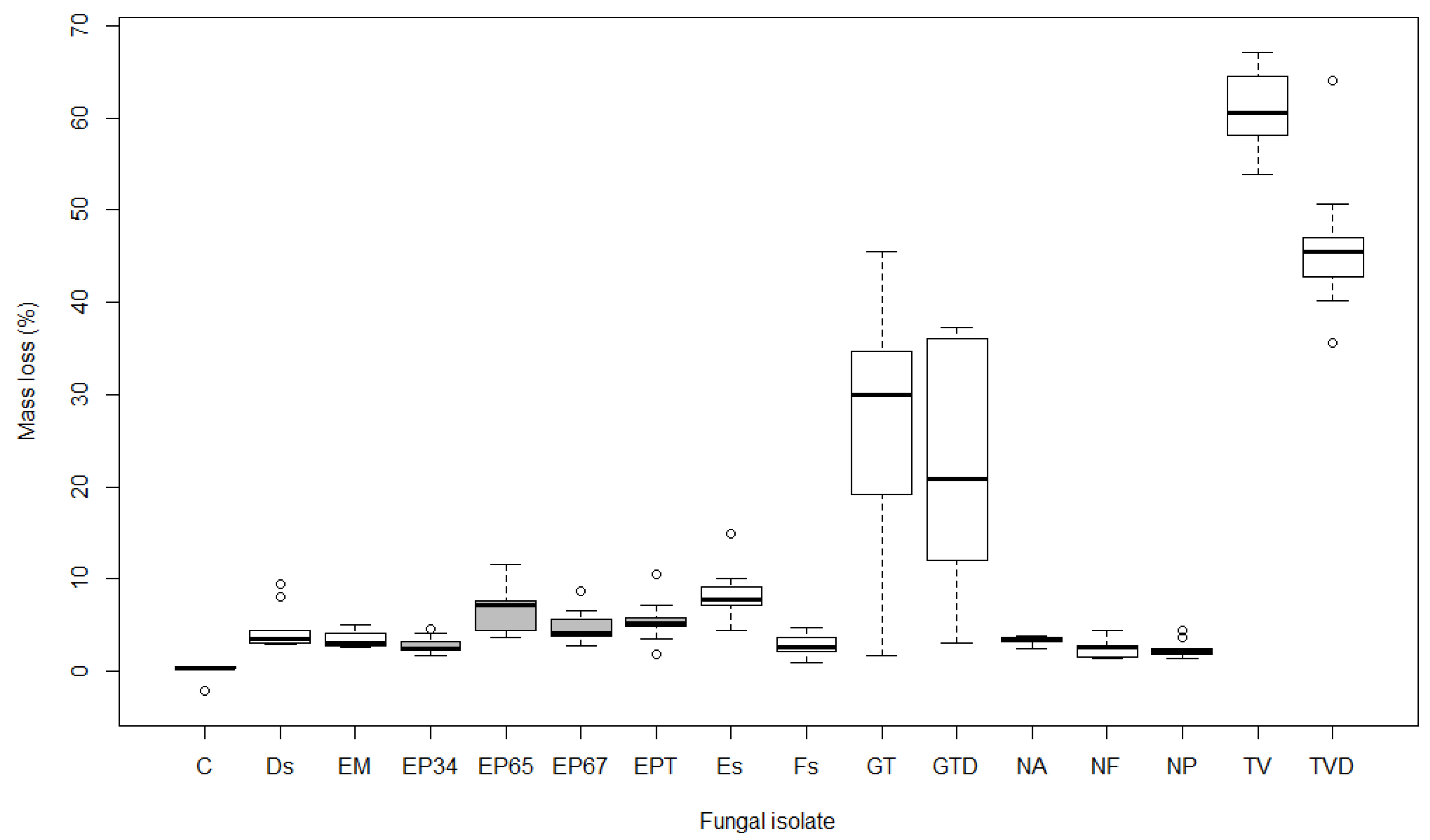
3.1. Mass Loss
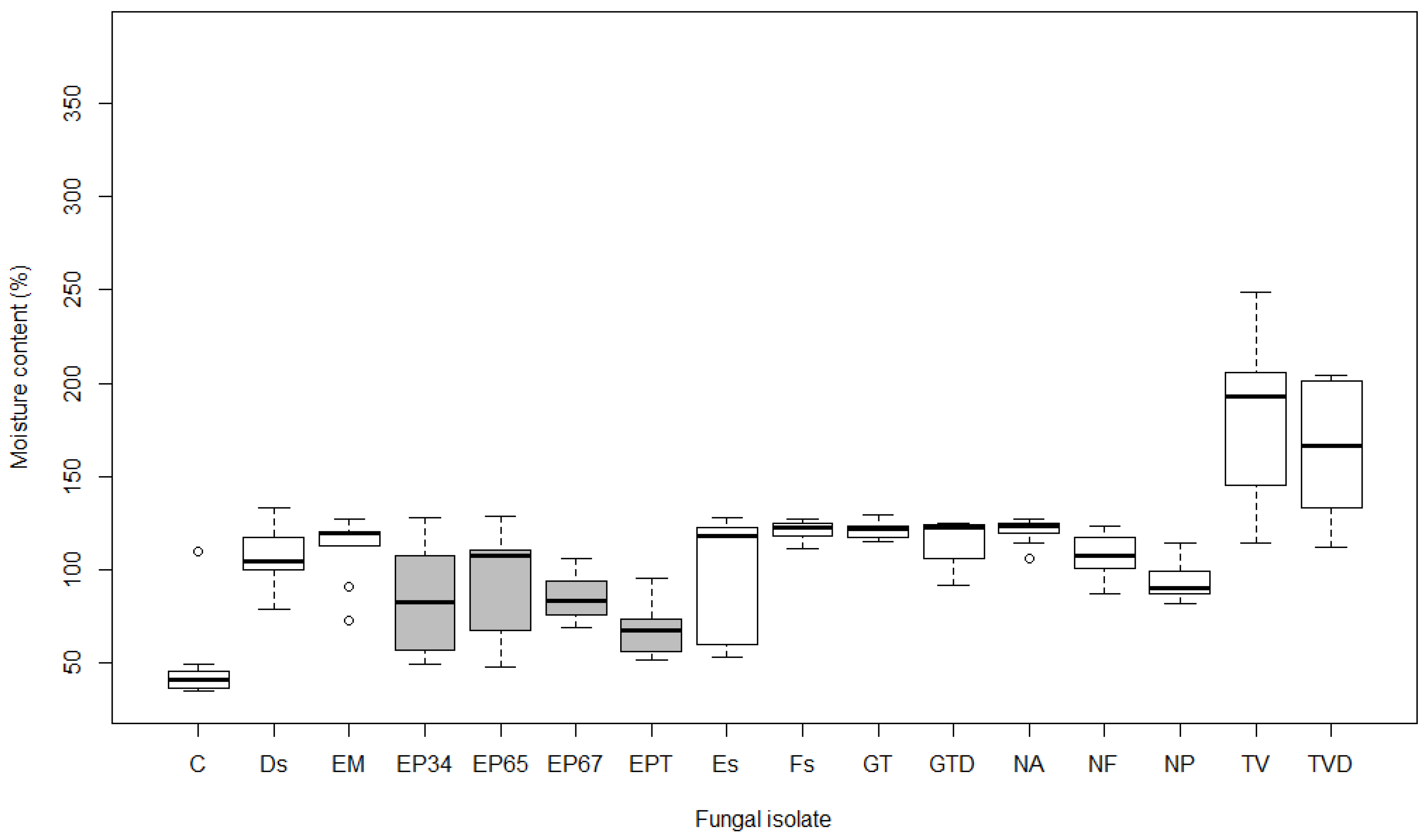
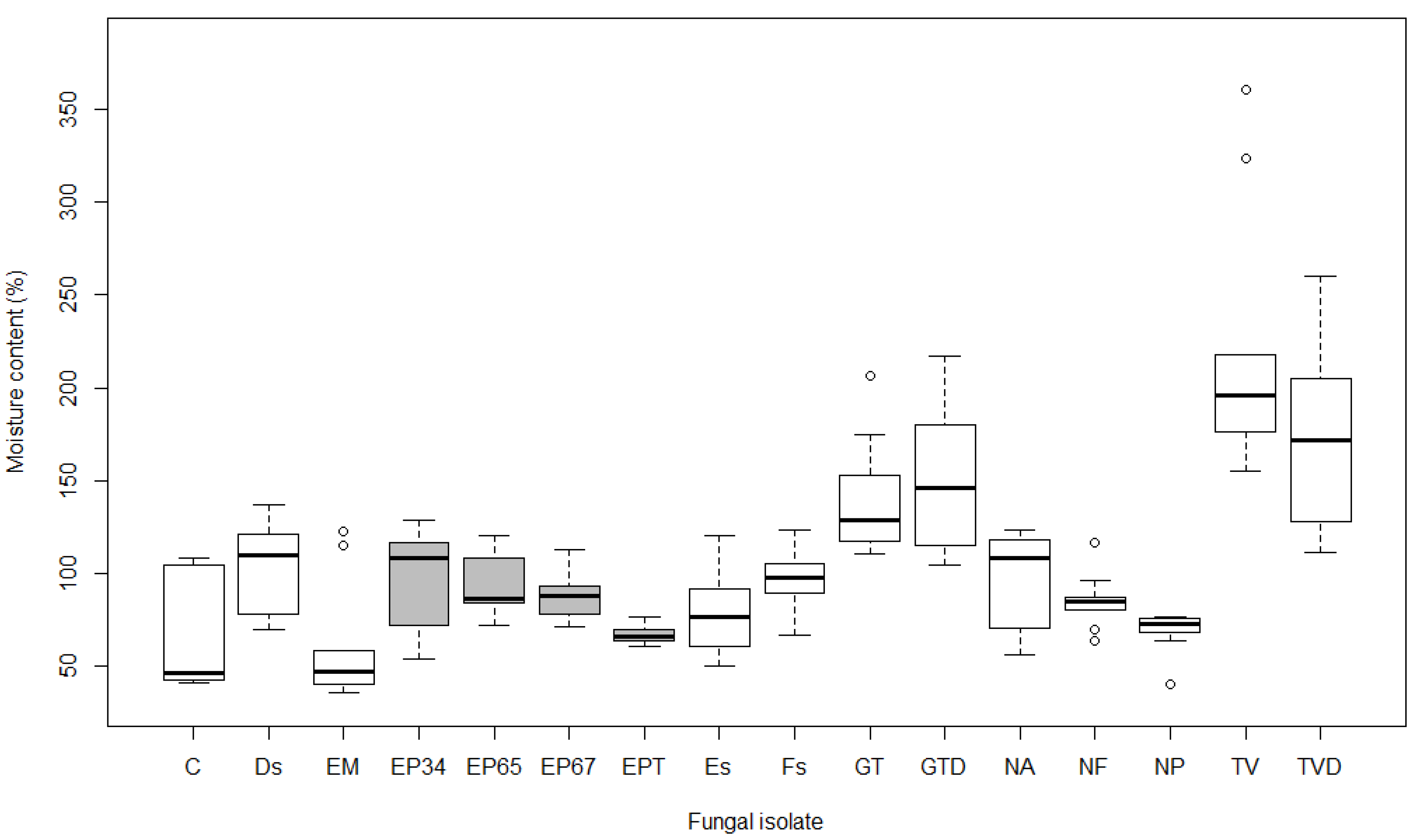
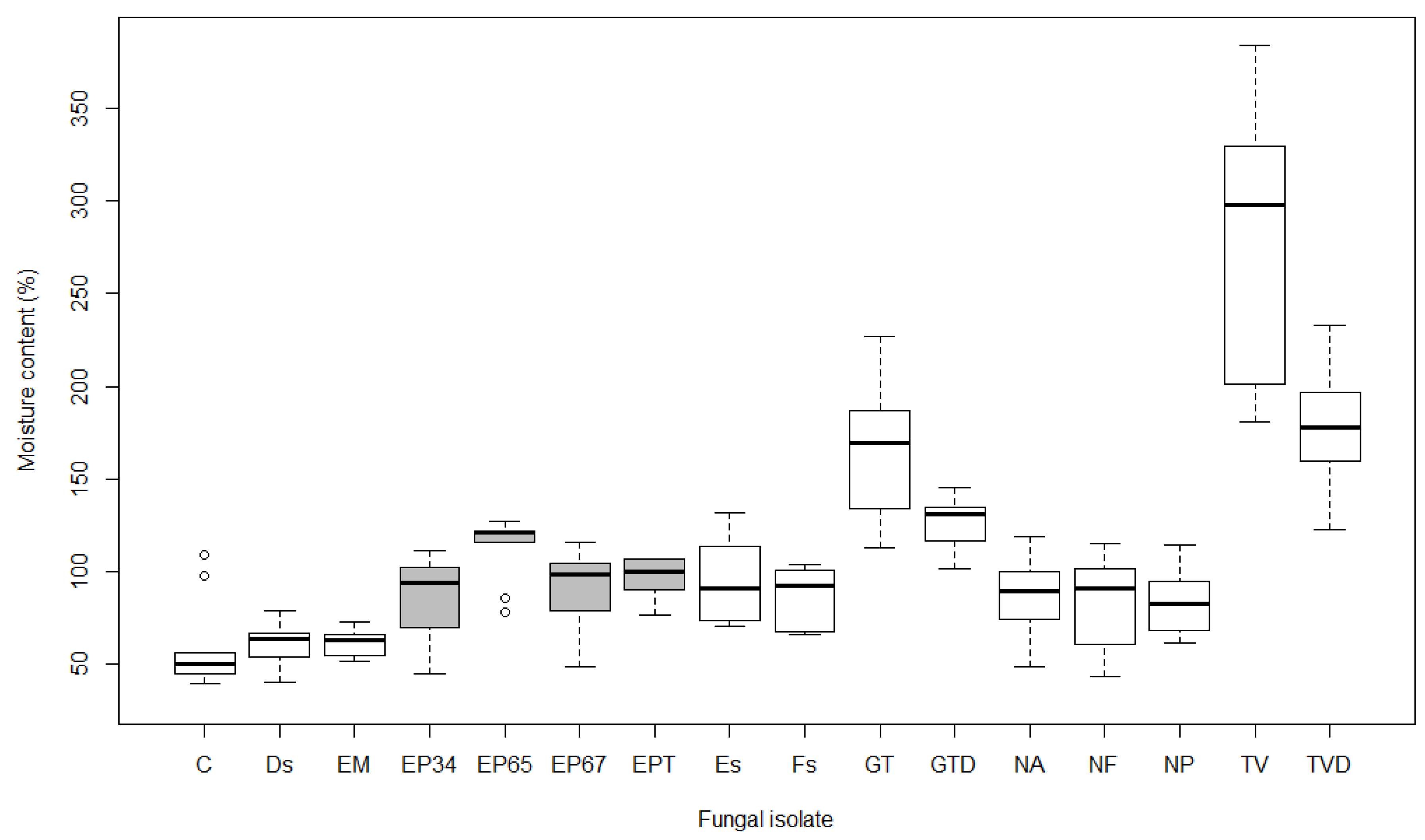
3.2. Moisture Content
3.3. Wood Density
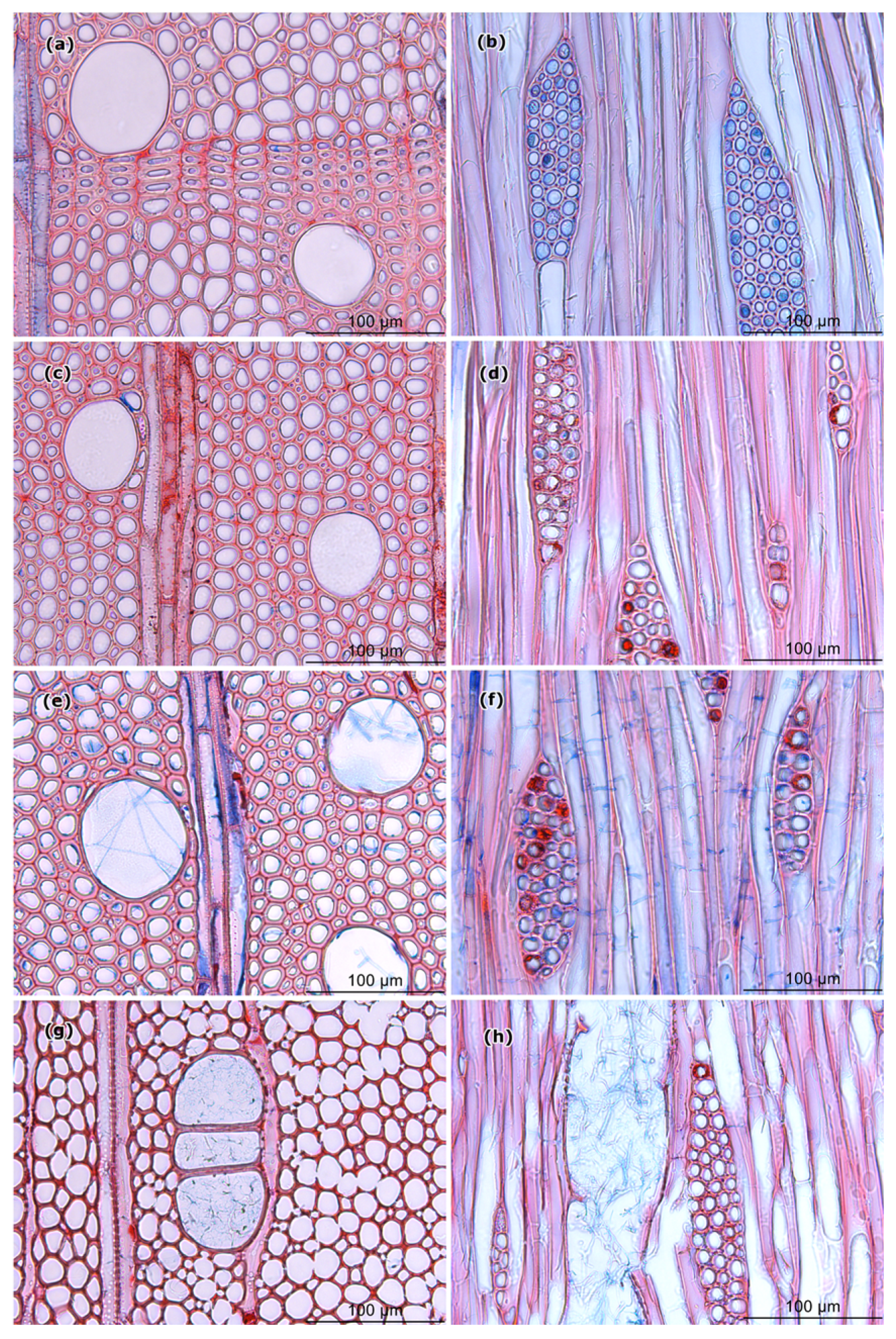
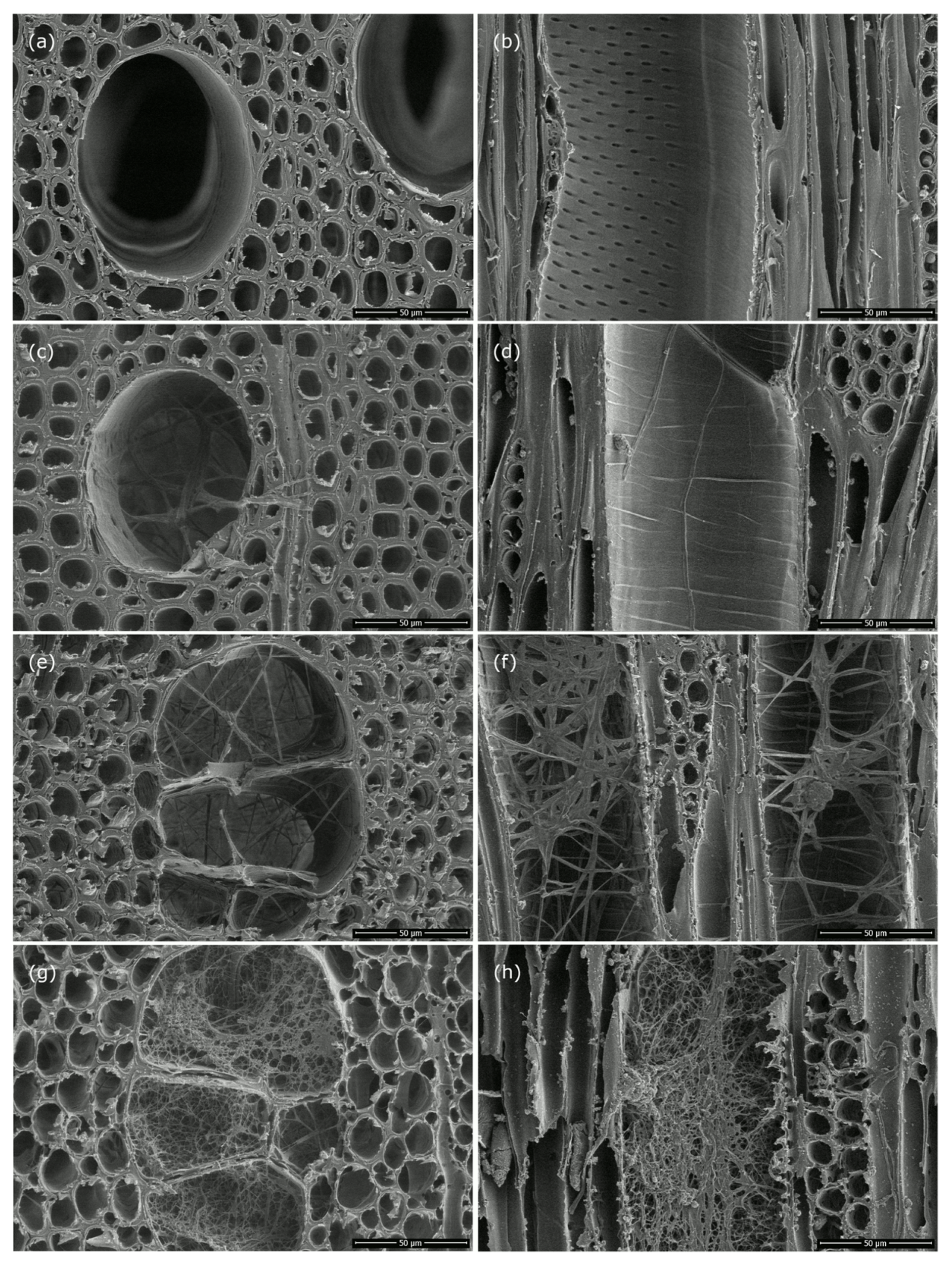
3.4. Light and Scanning Electron Microscopy
4. Discussion
4.1. Mass Loss
4.2. Moisture Content
4.3. Wood Structural Changes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shortle, W.C.; Dudzik, K.R. Wood Decay in Living and Dead Trees: A Pictorial Overview; United States Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2012; p. 26. [Google Scholar]
- Downer, A.J.; Perry, E.J. Wood decay fungi in landscape trees. In UC IPM Pest Notes; UC ANR Publication: Oakland, CA, USA, 2019; Volume 74109, p. 6. [Google Scholar]
- Schwarze, F.W.M.R. Wood decay under the microscope. Fungal Biol. Rev. 2007, 21, 133–170. [Google Scholar] [CrossRef]
- Mester, T.; Varela, E.; Tien, M. Wood degradation by brown-rot and white-rot fungi. In Genetics and Biotechnology; Kück, U., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 355–368. [Google Scholar]
- Shupe, T.; Lebow, S.; Ring, D. Causes and Control of Wood Decay, Degradation and Stain; Louisiana State University Agriculture Center: Baton Rouge, LA, USA, 2008; p. 27. [Google Scholar]
- Schwarze, F.W.M.R. Forest pathology: Heart rot and wood decay. In Encyclopedia of Forest Sciences; Burley, J., Evans, J., Youngquist, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 808–816. [Google Scholar]
- Worrall, J.J.; Anagnost, S.E.; Zabel, R.A. Comparison of wood decay among diverse lignicolous fungi. Mycologia 1997, 89, 199–219. [Google Scholar] [CrossRef]
- Shary, S.; Ralph, S.A.; Hammel, K.E. New insights into the ligninolytic capability of a wood decay ascomycete. Appl. Environ. Microb. 2007, 73, 6691–6694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäkelä, M.R.; Marinović, M.; Nousiainen, P.A.; Liawanag, A.J.M.; Benoit, I.; Sipilä, J.; Hatakka, A.I.; de Vries, R.; Hilden, S.K. Aromatic metabolism of filamentous fungi in relation to the presence of aromatic compounds in plant biomass. Adv. Appl. Microbiol. 2015, 91, 63–137. [Google Scholar] [CrossRef] [PubMed]
- Jurc, D.; Ogris, N.; Slippers, B.; Stenlid, J. First report of Eutypella canker of Acer pseudoplatanus in Europe. Plant Pathol. 2006, 55, 577. [Google Scholar] [CrossRef]
- Davidson, R.W.; Lorenz, R.C. Species of Eutypella and Schizoxylon associated with cankers of maple. Phytopathology 1938, 28, 733–745. [Google Scholar]
- Ogris, N.; Jurc, D.; Jurc, M. Spread risk of Eutypella canker of maple in Europe. Bull. OEPP/EPPO Bull. 2006, 36, 475–485. [Google Scholar] [CrossRef]
- French, W.J. Eutypella Canker on Species of Acer in New York State. Ph.D. Thesis, State University College of Forestry at Syracuse University, Syracuse, NY, USA, 1967. [Google Scholar]
- Pildain, M.B.; Novas, M.V.; Carmarán, C.C. Evaluation of anamorphic state, wood decay and production of lignin-modifying enzymes for diatrypaceous fungi from Argentina. J. Agric. Technol. 2005, 1, 81–96. [Google Scholar]
- Kleist, G.; Schmitt, U. Characterisation of a soft rot-like decay pattern caused by Coniophora puteana (Schum.) Karst. in sapelli Wood (Entandrophragma cylindricum Sprague). Holzforschung 2001, 55, 573–578. [Google Scholar] [CrossRef]
- Brglez, A.; Piškur, B.; Ogris, N. Eutypella parasitica and other frequently isolated fungi in wood of dead branches of young sycamore maple (Acer pseudoplatanus) in Slovenia. Forests 2020, 11, 467. [Google Scholar] [CrossRef] [Green Version]
- Bravery, F. A miniaturised wood-block test for the rapid evaluation of wood preservative fungicides. In Proceedings of the International Research Group on Wood Protection, Peebles, UK, 18–22 September 1978; pp. 57–65. [Google Scholar]
- Sutter, H.P. A new technique for screening fungicides for wood preservatives. Int. Biodeterior. 1987, 14, 95–99. [Google Scholar]
- Brown, J.; Caswell, S.; Williams, G.R. Development of miniblock test method for rapid evaluation of preservative performance against Basidiomycete fungi. In Proceedings of the International Research Group on Wood Preservation, Kyoto, Japan, 20–24 May 1991; pp. 1–11. [Google Scholar]
- CEN. EN 113—Wood Preservatives—Test Method for Determining the Protective Effectiveness against Wood Destroying Basidiomycetes.–Determination of the Toxic Values; European Committee for Standardization: Brussels, Belgium, 2002; p. 34. [Google Scholar]
- van der Wef, G.W.; Sass-Klaassen, U.G.W.; Mohren, G.M.J. The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 2007, 25, 103–112. [Google Scholar] [CrossRef]
- Srebotnik, E.; Messner, K. A Simple method that uses differential staining and Llight microscopy to assess the selectivity of wood delignification by white rot fungi. Appl. Environ. Microb. 1994, 60, 1383–1386. [Google Scholar] [CrossRef] [Green Version]
- von Aufsess, H. Mikroskopische Darstellung des Verholzungsgrades durch Färbemethoden. Holz als Roh-und Werkstoff 1973, 31, 24–33. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, 3.6.1; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Signorell, A. DescTools: Tools for Descriptive Statistics. R Package Version 0.99.30.; 2019. Available online: https://cran.r-project.org/web/packages/DescTools/index.html (accessed on 15 March 2020).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplry: A Grammar of Data Manipulation. R Package Version 0.8.3.; 2019. Available online: https://cran.r-project.org/web/packages/dplyr/index.html (accessed on 15 March 2020).
- Robinson, D.; Hayes, A. Broom: Convert Statistical Analysis Objects into Tidy Tibbles. R Package Version 0.5.2.; 2019. Available online: https://cran.r-project.org/web/packages/broom/index.html (accessed on 15 March 2020).
- de Vries, W.; Reinds, G.J.; Posch, M.; Sanz, M.J.; Krause, G.H.M.; Calatayud, V.; Renaud, J.P.; Vel, E.M.; Dobertin, M.; Gundersen, P.; et al. Intensive Monitoring of Forest Ecosystems in Europe; European Commission: Brussels, Geneva, 2003; p. 163. [Google Scholar]
- Robinson, S.C.; Laks, P.E.; Richter, D.L.; Pickens, J.B. Evaluating loss of machinability in spalted sugar maple. For. Prod. J. 2007, 57, 33–37. [Google Scholar]
- Humar, M.; Lesar, B.; Thaler, N.; Kržišnik, D.; Kregar, N.; Drnovšek, S. Quality of copper impregnated wood in Slovenian hardware stores. Drvna Ind. 2018, 69, 121–126. [Google Scholar] [CrossRef]
- Reinprecht, L.; Vidholdová, Z.; Gašpar, F. Decay inhibition of maple wood with nano-zinc oxide used in combination with essential oils. Acta Facultatis Xylologiae Zvolen 2016, 58, 51–58. [Google Scholar] [CrossRef]
- Kielmann, B.C.; Adamopoulos, S.; Militz, H.; Mai, C. Decay resistance of ash, beech and maple wood modified with N-methylol melamine and a metal complex dye. Int. Biodeter. Biodegr. 2014, 89, 110–114. [Google Scholar] [CrossRef]
- Meyer, L.; Brischke, C.; Treu, A.; Larsson-Brelid, P. Critical moisture conditions for fungal decay of modified wood by basidiomycetes as detected by pile tests. Holzforschung 2016, 70, 331–339. [Google Scholar] [CrossRef]
- Schmidt, O. Wood and Tree Fungi: Biology, Damage, Protection, and Use; Springer: Berlin/Heidelberg, Germany, 2006; p. 336. [Google Scholar]
- Griffin, D.M. Water potential and wood-decay fungi. Annu. Rev. Phytopathol. 1977, 15, 319–329. [Google Scholar] [CrossRef]
- Zelinka, S.L.; Kirker, G.T.; Bishell, A.B.; Glass, S.V. Effects of wood moisture content and the level of acetylation on brown rot decay. Forests 2020, 11, 299. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.A.; Cowling, E.B. Influence of various initial moisture contents on decay of Sitka spruce and sweetgum sapwood by Polyporus versicolor in the soil-block test. Phytopathology 1973, 63, 235–237. [Google Scholar] [CrossRef]
- Humar, M.; Fabčič, B.; Zupančič, M.; Pohleven, F.; Oven, P. Influence of xylem growth ring width and wood density on durability of oak heartwood. Int. Biodeter. Biodegr. 2008, 62, 368–371. [Google Scholar] [CrossRef]
- Brischke, C.; Unger, W. Potential hazards and degrading agents. In Performance of Bio-Based Building Materials; Jones, D., Brischke, C., Eds.; Elsevier: Amsterdam, Netherlands, 2017; pp. 188–203. [Google Scholar]
- Wilcox, W.W. Comparison of scanning electron microscopy and light microscopy for the diagnosis of early stages of brown rot wood decay. IAWA J. 1993, 14, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, W.A.; Lyon, H.H.; Johnson, W.T. Eutypella canker of maple. In Diseases of Trees and Shrubs; Cornell University Press: Ithaca, NY, USA, 1987; pp. 220–221. [Google Scholar]
- Ogris, N.; Piškur, B.; Jurc, D. Some morphological aspects of Eutypella canker of maple (Eutypella parasitica). In Proceedings of the IUFRO Working Party 7.02.02, Eğirdir, Turkey, 11–16 May 2009; pp. 150–161. [Google Scholar]
- Ogris, N.; Jurc, D.; Jurc, M. Javorov rak (Eutypella parasitica: Ascomycota: Fungi) na gorskem javorju in maklenu: Značilnosti in razlike. Gozdarski Vestnik 2005, 63, 411–418. [Google Scholar]
- Kliejunas, J.T.; Kuntz, J.E. Eutypella canker, characteristics and control. For. Chron. 1974, 50, 106–108. [Google Scholar] [CrossRef] [Green Version]
- Dadzie, P.K.; Amoah, M. Density, some anatomical properties and natural durability of stem and branch wood of two tropical hardwood species for ground applications. Eur. J. Wood Prod. 2015, 73, 759–773. [Google Scholar] [CrossRef]
- Shigo, A.L. Biology of decay and wood quality. In Biological Transformation of Wood by Microorganisms; Liese, W., Ed.; Springer: Berlin/Heidelberg, Germany, 1975; p. 15. [Google Scholar]
- Shigo, A.L.; Marx, H.G. Compartmentalization of Decay in Trees; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1977; p. 73. [Google Scholar]
- Green, D.J.; Shortle, W.C.; Shigo, A.L. Compartmentalization of discolored and decayed wood in red maple branch stubs. For. Sci. 1981, 27, 519–522. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Label 5 | Collection Number 6 |
|---|---|---|
| Diaporthe sp. Nitschke 1 | Ds | ZLVG 788 |
| Eutypa sp. Tul. & C. Tul. 1 | Es | ZLVG 790 |
| Eutypa maura (Fr.) Sacc. 1 | EM | ZLVG 789 |
| Eutypella parasitica R.W. Davidson & R.C. Lorenz | EP34 | ZLVG 34 |
| EP65 | ZLVG 65 | |
| EP67 | ZLVG 67 | |
| EPT | ZLVG 805 | |
| Fusarium sp. Link 1 | Fs | ZLVG 792 |
| Gloeophyllum trabeum (Pers.) Murrill 2 | GT | ZIM L017 |
| GTD | DSM 1398 | |
| Neocucurbitaria acerina Wanas., Camporesi, E.B.G. Jones & K.D. Hyde 1 | NA | ZLVG 794 |
| Neonectria faginata (M.L. Lohman, A.M.J. Watson & Ayers) Castl. & Rossman 3 | NF | ZLVG 807 |
| Neonectria punicea (J.C. Schmidt) Castl. & Rossman 3 | NP | ZLVG 806 |
| Trametes versicolor (L.) Lloyd 4 | TV | ZIM L057 |
| TVD | DSM 3086 |
| A. pseudoplatanus ΔMC (%) | A. platanoides ΔMC (%) | A. campestre ΔMC (%) | |
|---|---|---|---|
| Diaporthe sp. | 58.7 | 39.3 | 2.3 |
| Eutypa sp. | 53.8 | 13.9 | 35.4 |
| Eutypa maura | 65.2 | −4.3 | 2.8 |
| Eutypella parasitica1 | 35.7 | 21.5 | 38.1 |
| Fusarium sp. | 73.6 | 30.2 | 27.9 |
| Gloeophyllum trabeum1 | 71.0 | 78.8 | 86.5 |
| Neocucurbitaria acerina | 73.7 | 33.3 | 29.0 |
| Neonectria faginata | 60.0 | 20.0 | 23.7 |
| Neonectria punicea | 45.7 | 4.1 | 24.7 |
| Trametes versicolor1 | 125.0 | 129.8 | 171.6 |
| Average | 64.4 | 42.6 | 54.3 |
| Tree Species | Wood Density (kg/m3) | SD 1 | Statistic Group 2 |
|---|---|---|---|
| Acer pseudoplatanus | 616 | 18 | ab |
| Acer platanoides | 611 | 26 | b |
| Acer campestre | 620 | 29 | a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brglez, A.; Piškur, B.; Humar, M.; Gričar, J.; Ogris, N. The Effect of Eutypella parasitica on the Wood Decay of Three Maple Species. Forests 2020, 11, 671. https://doi.org/10.3390/f11060671
Brglez A, Piškur B, Humar M, Gričar J, Ogris N. The Effect of Eutypella parasitica on the Wood Decay of Three Maple Species. Forests. 2020; 11(6):671. https://doi.org/10.3390/f11060671
Chicago/Turabian StyleBrglez, Ana, Barbara Piškur, Miha Humar, Jožica Gričar, and Nikica Ogris. 2020. "The Effect of Eutypella parasitica on the Wood Decay of Three Maple Species" Forests 11, no. 6: 671. https://doi.org/10.3390/f11060671
APA StyleBrglez, A., Piškur, B., Humar, M., Gričar, J., & Ogris, N. (2020). The Effect of Eutypella parasitica on the Wood Decay of Three Maple Species. Forests, 11(6), 671. https://doi.org/10.3390/f11060671