Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Occurrence Data
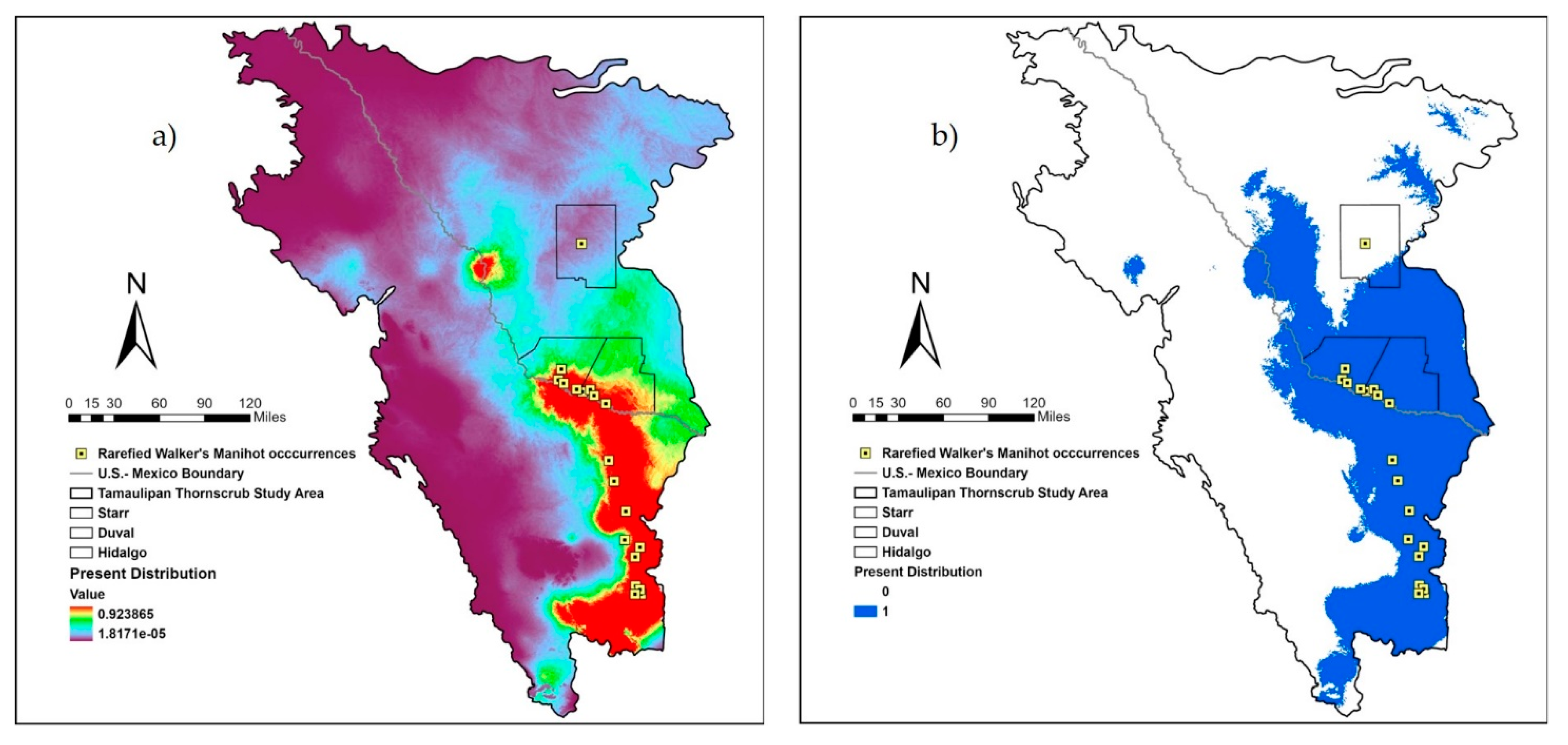
2.2. Study Area and Bioclimatic Variables
2.3. Running MaxEnt and Creating Consensus Models
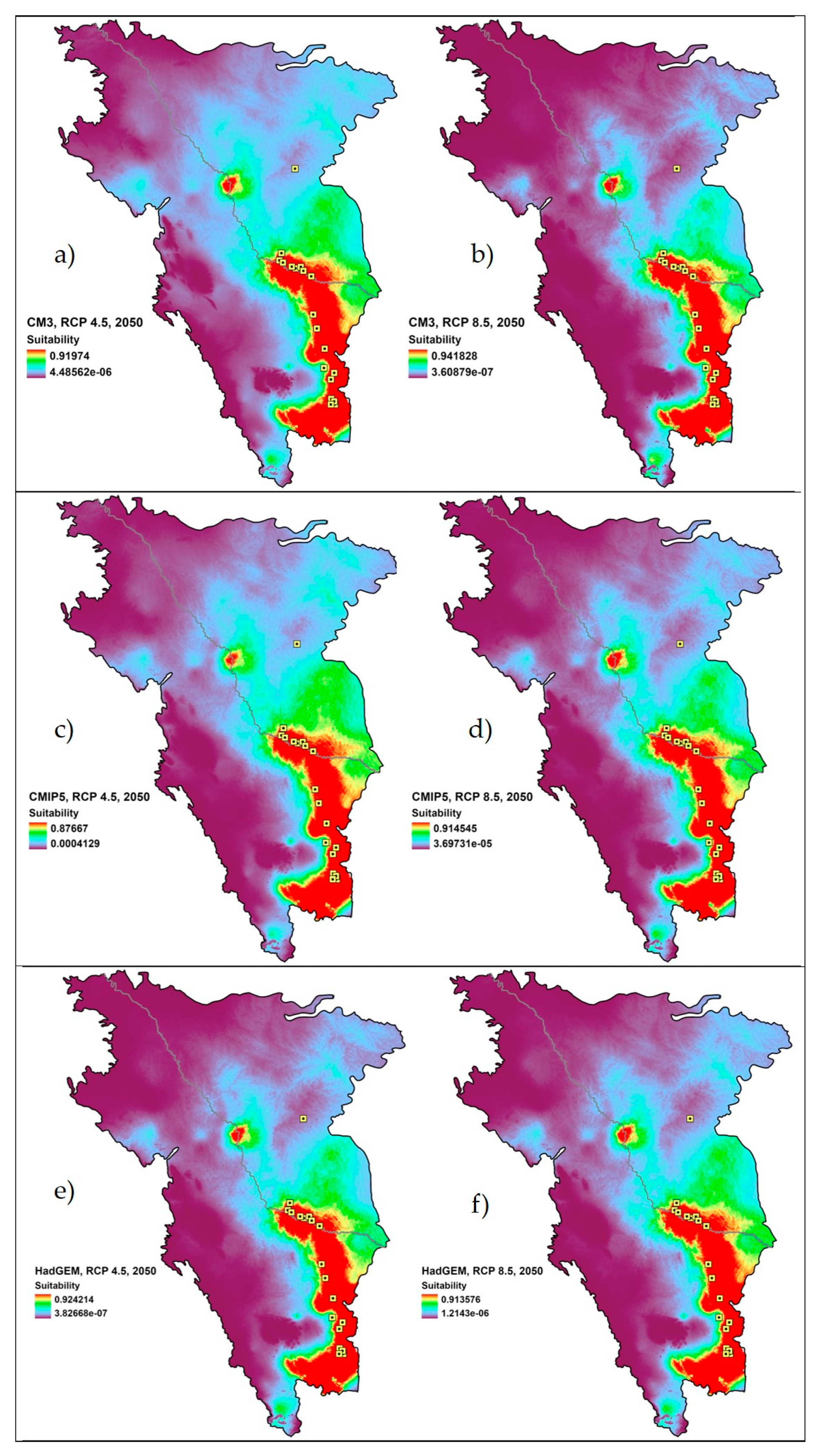
2.4. Calculating Percent Change of Geographic Distribution and Model Evaluation
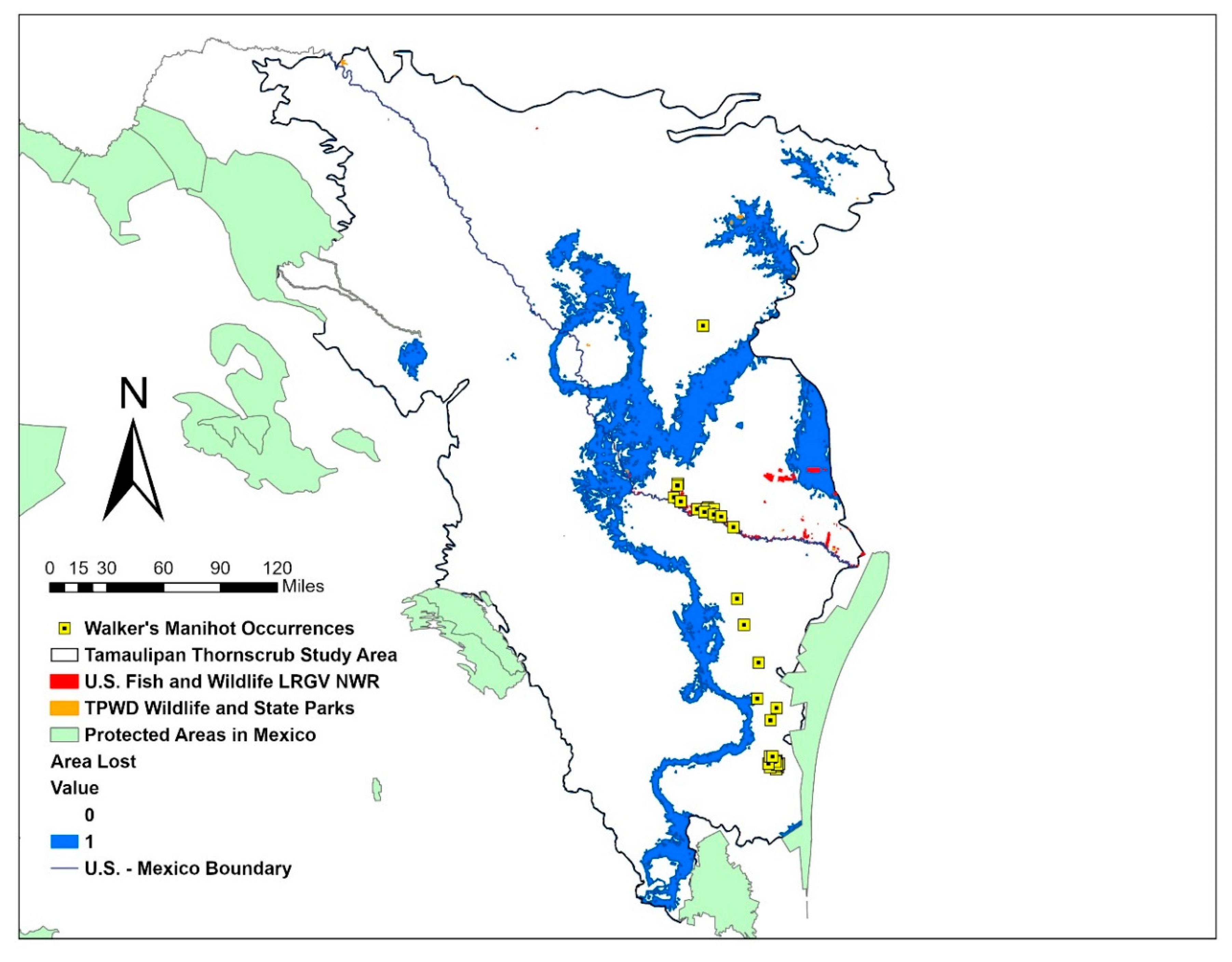
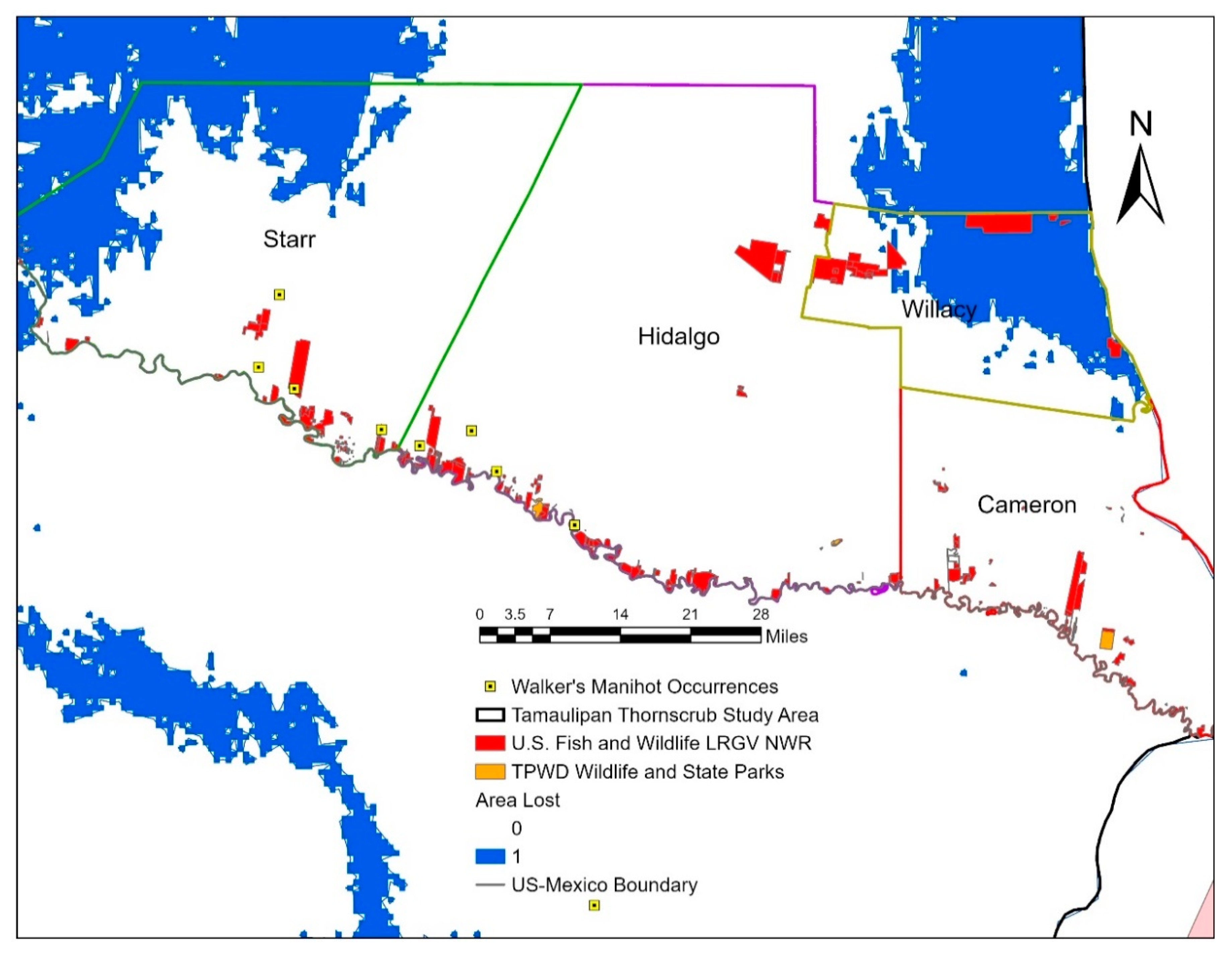
2.5. Protected Areas Maps
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paul, A.; Bharali, S.; Khan, M.L.; Tripathi, O.P. Anthropogenic disturbances led to risk of extinction of Taxus wallichiana Zuccarini, an endangered medicinal tree in Arunachal Himalaya. Nat. Areas J. 2013, 33, 447–454. [Google Scholar] [CrossRef]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The spread of invasive species and infectious disease as drivers of ecosystem change. Front. Ecol. Environ. 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Penuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- IŞIK, K. Rare and endemic species: Why are they prone to extinction? Turk. J. Bot. 2011, 35, 411–417. [Google Scholar] [CrossRef]
- Pulliam, H.R. On the relationship between niche and distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Richardson, D.M.; Midgley, G.F. Will Climate Change Promote Alien Plant Invasions? Springer: Berlin/Heidelberg, Germany, 2008; pp. 197–211. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Reisinger, A. Climate Change 2007: Synthesis Report. Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2007; p. 104. [Google Scholar]
- Brook, B.W.; Sodhi, N.S.; Bradshaw, C.J. Synergies among extinction drivers under global change. Trends Ecol. Evol. 2008, 23, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Leimu, R.; Vergeer, P.; Angeloni, F.; Ouborg, N.J. Habitat fragmentation, climate change, and inbreeding in plants. Ann. N. Y. Acad. Sci. 2010, 1195, 84–98. [Google Scholar] [CrossRef]
- El-Keblawy, A. Impact of climate change on biodiversity loss and extinction of endemic plants of arid land mountains. J. Biodiver. Endang. Spp. 2014, 2, 2. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Strayer, D.L. Usefulness of bioclimatic models for studying climate change and invasive species. Ann. N. Y. Acad. Sci. 2008, 1134, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Abolmaali, S.M.; Tarkesh, M.; Bashari, H. MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inform. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecologica 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Vieilledent, G.; Cornu, C.; Sanchez, A.C.; Pock-Tsy, J.M.; Danthu, P. Vulnerability of baobab species to climate change and effectiveness of the protected area network in Madagascar: Towards new conservation priorities. Biol. Conserv. 2013, 166, 11–22. [Google Scholar] [CrossRef]
- Yu, J.; Wang, C.; Wan, J.; Han, S.; Wang, Q.; Nie, S. A model-based method to evaluate the ability of nature reserves to protect endangered tree species in the context of climate change. For. Ecol. Manag. 2014, 327, 48–54. [Google Scholar] [CrossRef]
- Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modelling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Ardestani, E.G.; Tarkesh, M.; Bassiri, M.; Vahabi, M.R. Potential habitat modeling for reintroduction of three native plant species in central Iran. J. Arid Land. 2015, 7, 381–390. [Google Scholar] [CrossRef]
- Hole, D.G.; Huntley, B.; Arinaitwe, J.; Butchart, S.H.; Collingham, Y.C.; Fishpool, L.D.; Pain, D.J.; Willis, S.G. Toward a management framework for networks of protected areas in the face of climate change. Conserv. Biol. 2011, 25, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Clayton, P.W. Walker’s Manioc Manihot Walkerae Recovery Plan; U.S. Fish and Wildlife Service, Region 2: Albuquerque, NM, USA, 1993. [Google Scholar]
- Best, C.; Miller, A.; Cobb, R. Walker’s Manioc (Manihot walkerae) 5-Year Review: Summary and Evaluation; U.S. Fish and Wildlife Service: Albuquerque, NM, USA, 2009. [Google Scholar]
- Vera-Sánchez, K.S.; Nassar, N. Manihot Walkerae. The IUCN Red List of Threatened Species. 2019. Available online: https://www.iucnredlist.org/species/20755842/20756066 (accessed on 7 April 2020).
- Leslie, D.M., Jr. An International Borderland of Concern: Conservation of Biodiversity in the Lower Rio Grande Valley; No. 2016-5078; U.S. Geological Survey: Reston, VA, USA, 2016. [Google Scholar]
- Jahrsdoerfer, S.E.; Leslie, D.M., Jr. Tamaulipan brushland of the Lower Rio Grande Valley of South Texas: Description, human impacts, and management options. U.S. Fish Wildl. Serv. Biol. Rep. 1988, 88, 1–63. [Google Scholar]
- Cook, T.; Adams, J.; Valero, A.; Schipper, J.; Allnutt, T. Southern North America: Southern United States into Northeastern Mexico. World Wildlife Fund. Available online: https://www.worldwildlife.org/ecoregions/na1312 (accessed on 24 April 2020).
- Marshall, V.M.; Lewis, M.M.; Ostendorf, B. Buffel grass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: A review. J. Arid Environ. 2012, 78, 1–2. [Google Scholar] [CrossRef]
- Saravanan, R.A.; Ravi, V.; Stephen, R.; Thajudhin, S.H.; George, J. Post-harvest physiological deterioration of cassava (Manihot esculenta)-A review. Indian J. Ag. Sci. 2016, 86, 1383–1390. [Google Scholar]
- Zainuddin, I.M.; Fathoni, A.; Sudarmonowati, E.; Beeching, J.R.; Gruissem, W.; Vanderschuren, H. Cassava post-harvest physiological deterioration: From triggers to symptoms. Postharvest Biol. Technol. 2018, 142, 115–123. [Google Scholar] [CrossRef]
- Morante, N.; Sánchez, T.; Ceballos, H.; Calle, F.; Pérez, J.C.; Egesi, C.; Cuambe, C.E.; Escobar, A.F.; Ortiz, D.; Chávez, A.L.; et al. Tolerance to postharvest physiological deterioration in cassava roots. Crop Sci. 2010, 50, 1333–1338. [Google Scholar] [CrossRef]
- Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Khoury, C.K.; Müller, J.V.; Toll, J. Adapting agriculture to climate change: A global initiative to collect, conserve, and use crop wild relatives. Agroecol. Sustain. Food Syst. 2014, 38, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Kurpis, J.; Serrato-Cruz, M.A.; Arroyo, T.P. Modeling the effects of climate change on the distribution of Tagetes lucida Cav. (Asteraceae). Glob. Ecol. Conser. 2019, 20, e00747. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- ESRI 2020. ArcGIS Pro 2.5.1; Environmental Systems Research Institute: Redlands, CA, USA, 2020. [Google Scholar]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2008, 15, 59–69. [Google Scholar] [CrossRef]
- Norris, D. Model thresholds are more important than presence location type: Understanding the distribution of lowland tapir (Tapirus terrestris) in a continuous Atlantic forest of southeast Brazil. Trop. Conserv. Sci. 2014, 7, 529–547. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Osorio-Olvera, L.; Barve, V.; Barve, N.; Soberón, J.; Falconi, M. Niche Toolbox: From Getting Biodiversity Data to Evaluating Species Distribution Models in a Friendly GUI Environment. R Package Version 0.2.5.4. 2018. Available online: http://shiny.conabio.gob.mx:3838/nichetoolb2/ (accessed on 13 April 2020).
- Sosa, V.; Loera, I.; Angulo, D.F.; Vásquez-Cruz, M.; Gándara, E. Climate change and conservation in a warm North American desert: Effect in shrubby plants. PeerJ 2019, 7, e6572. [Google Scholar] [CrossRef]
- Borzée, A.; Andersen, D.; Groffen, J.; Kim, H.T.; Bae, Y.; Jang, Y. Climate change-based models predict range shifts in the distribution of the only Asian plethodontid salamander: Karsenia koreana. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Stanton, J.C.; Pearson, R.G.; Horning, N.; Ersts, P.; Reşit Akçakaya, H. Combining static and dynamic variables in species distribution models under climate change. Methods Ecol. Evol. 2012, 3, 349–357. [Google Scholar] [CrossRef]
- Téllez-Valdés, O.; D¡Vila-Aranda, P. Protected areas and climate change: A case study of the Cacti in the Tehuacán-Cuicatlán Biosphere Reserve, México. Conserv. Biol. 2003, 17, 846–853. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Iglesius, A.; Yang, X.B.; Epstein, P.R.; Chivian, E. Climate change and extreme weather events-Implications for food production, plant diseases, and pests. Glob. Chang. Hum. Health 2001, 2, 90–104. [Google Scholar] [CrossRef]
- Greenwald, N.; Segee, B.; Curry, T.; Bradley, C. A Wall in the Wild: The Disastrous Impacts of Trump’s Border Wall on Wildlife; Center for Biological Diversity: Tucson, AZ, USA, 2017. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Explanation | % Contribution |
|---|---|---|
| BIO1 | Annual Mean Temperature | 37.1 |
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp-min temp)) | 0.3 |
| BIO3 | Isothermality (BIO2/BIO7) × 100 | 1 |
| BIO4 | Temperature Seasonality (standard deviation × 100) | |
| BIO5 | Max Temperature of Warmest Month | |
| BIO6 | Min Temperature of Coldest Month | |
| BIO7 | Temperature Annual Range (BIO5-BIO6) | 20.3 |
| BIO8 | Mean Temperature of Wettest Quarter | |
| BIO9 | Mean Temperature of Driest Quarter | |
| BIO10 | Mean Temperature of Warmest Quarter | 0 |
| BIO11 | Mean Temperature of Coldest Quarter | |
| BIO12 | Annual Precipitation | 7.1 |
| BIO13 | Precipitation of Wettest Month | 0 |
| BIO14 | Precipitation of Driest Month | 2.1 |
| BIO15 | Precipitation of Seasonality (Coefficient of Variation) | 13.7 |
| BIO16 | Precipitation of Wettest Quarter | |
| BIO17 | Precipitation of Driest Quarter | |
| BIO18 | Precipitation of Warmest Quarter | |
| BIO19 | Precipitation of Coldest Quarter | 18.3 |
| Present | 2050 | ||||||
| RCP 4.5 | RCP 8.5 | ||||||
| CM3 | CMIP5 | HadGEM | CM3 | CMIP5 | HadGEM | ||
| Suitable Area km2 | 75,901 | 88,186 | 88,571 | 66,345 | 59,657 | 72,352 | 73,870 |
| % Change | +7.20 | +7.42 | −5.60 | −9.52 | −2.08 | −1.19 | |
| Present | 2070 | ||||||
| RCP 4.5 | RCP 8.5 | ||||||
| CM3 | CMIP5 | HadGEM | CM3 | CMIP5 | HadGEM | ||
| Suitable Area km2 | 75,901 | 63,995 | 51,398 | 71,449 | 52,653 | 65,156 | 68,845 |
| % Change | −6.98 | −14.37 | −2.61 | −13.63 | −6.30 | −4.13 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garza, G.; Rivera, A.; Venegas Barrera, C.S.; Martinez-Ávalos, J.G.; Dale, J.; Feria Arroyo, T.P. Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae. Forests 2020, 11, 689. https://doi.org/10.3390/f11060689
Garza G, Rivera A, Venegas Barrera CS, Martinez-Ávalos JG, Dale J, Feria Arroyo TP. Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae. Forests. 2020; 11(6):689. https://doi.org/10.3390/f11060689
Chicago/Turabian StyleGarza, Gisel, Armida Rivera, Crystian Sadiel Venegas Barrera, José Guadalupe Martinez-Ávalos, Jon Dale, and Teresa Patricia Feria Arroyo. 2020. "Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae" Forests 11, no. 6: 689. https://doi.org/10.3390/f11060689
APA StyleGarza, G., Rivera, A., Venegas Barrera, C. S., Martinez-Ávalos, J. G., Dale, J., & Feria Arroyo, T. P. (2020). Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae. Forests, 11(6), 689. https://doi.org/10.3390/f11060689