Biogeographic, Atmospheric, and Climatic Factors Influencing Tree Growth in Mediterranean Aleppo Pine Forests

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
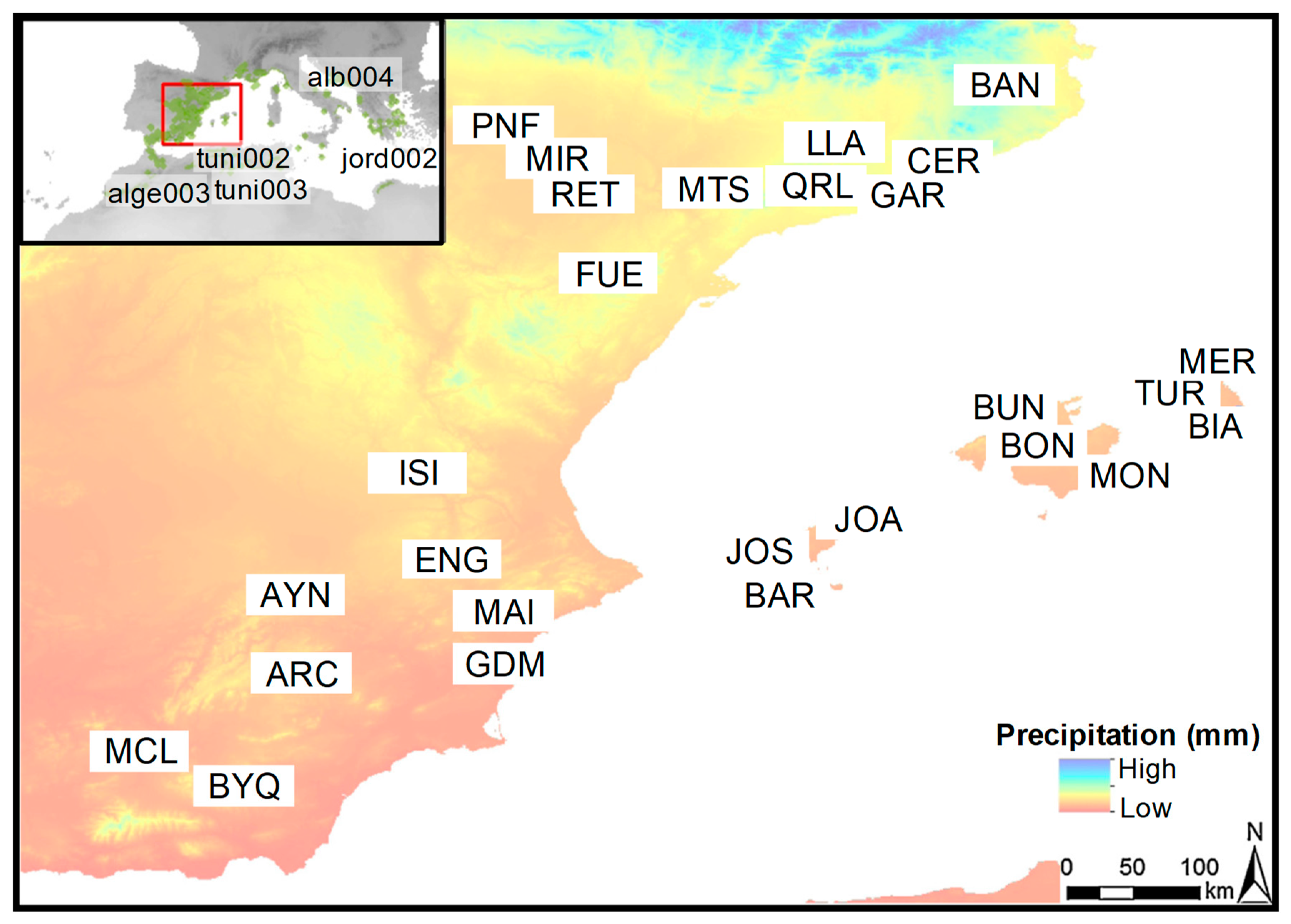
2.1. Study Region and Tree Species
2.2. Climate Data and Climate Indices Representing Major Atmospheric Patterns
2.3. Tree-Ring Width Data
2.4. Processing Tree-Ring Width and Climate Data
2.5. Statistical Analyses
2.6. VS-Lite Growth Model
3. Results
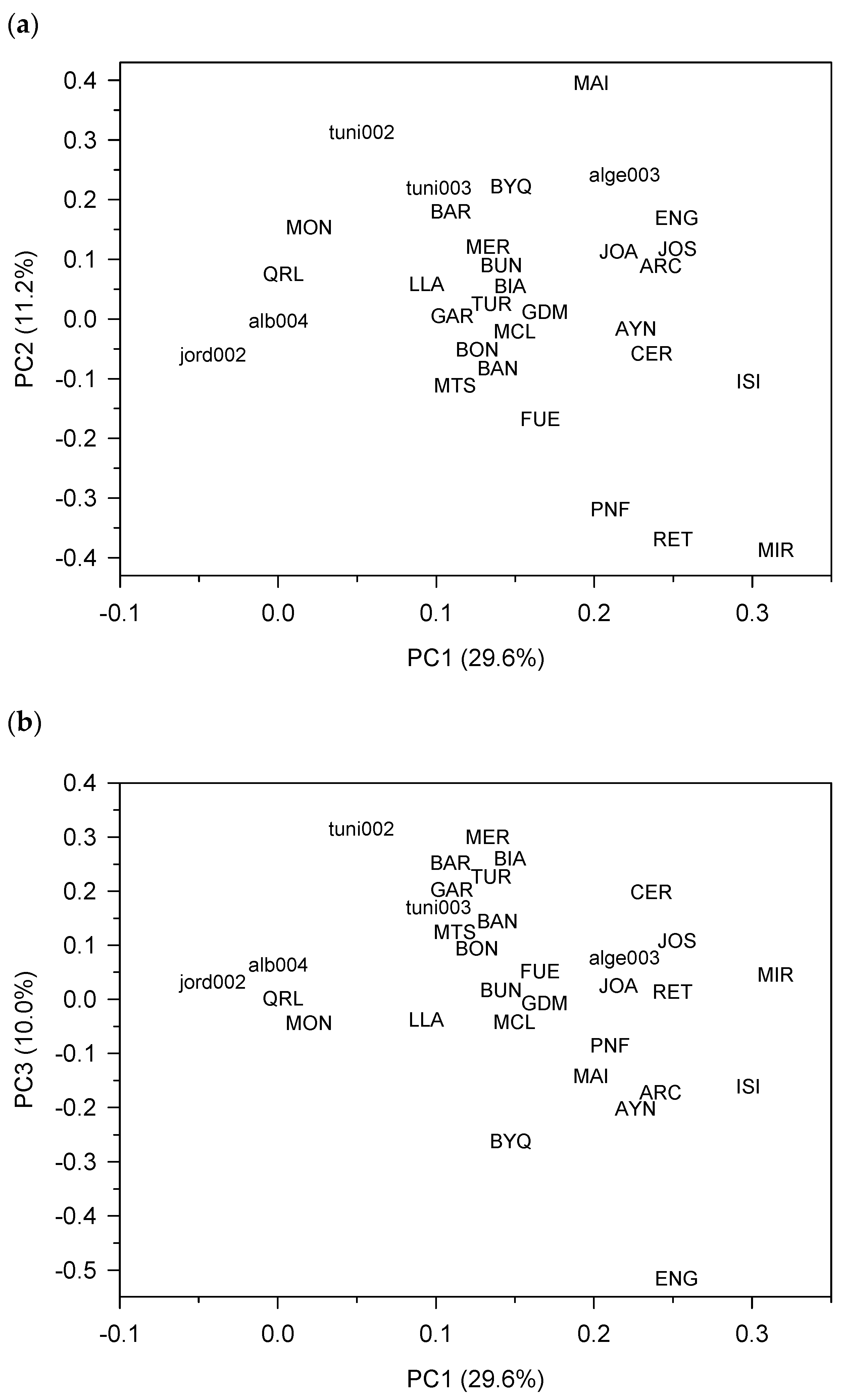
3.1. Tree-Ring Statistics and Sites Features
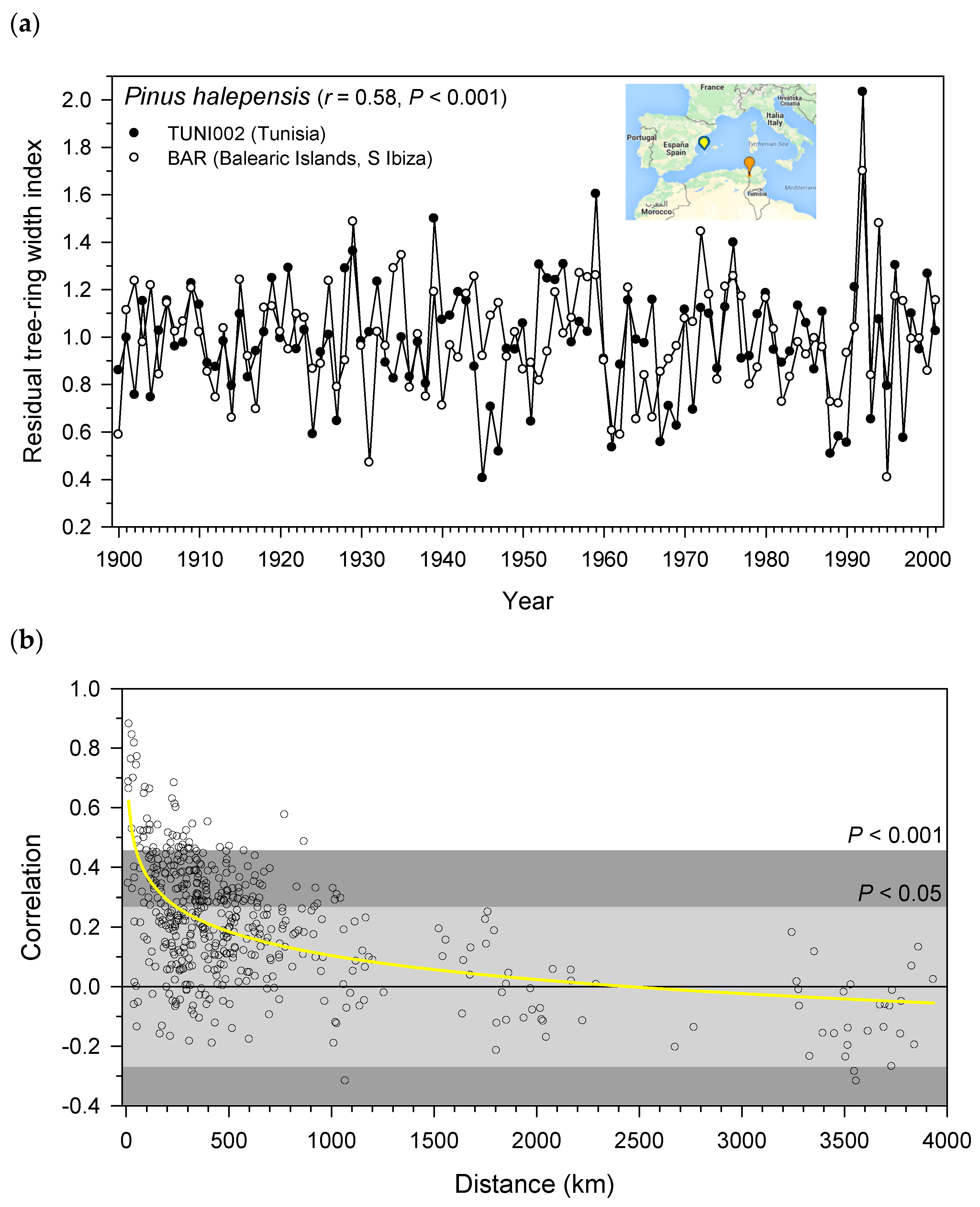
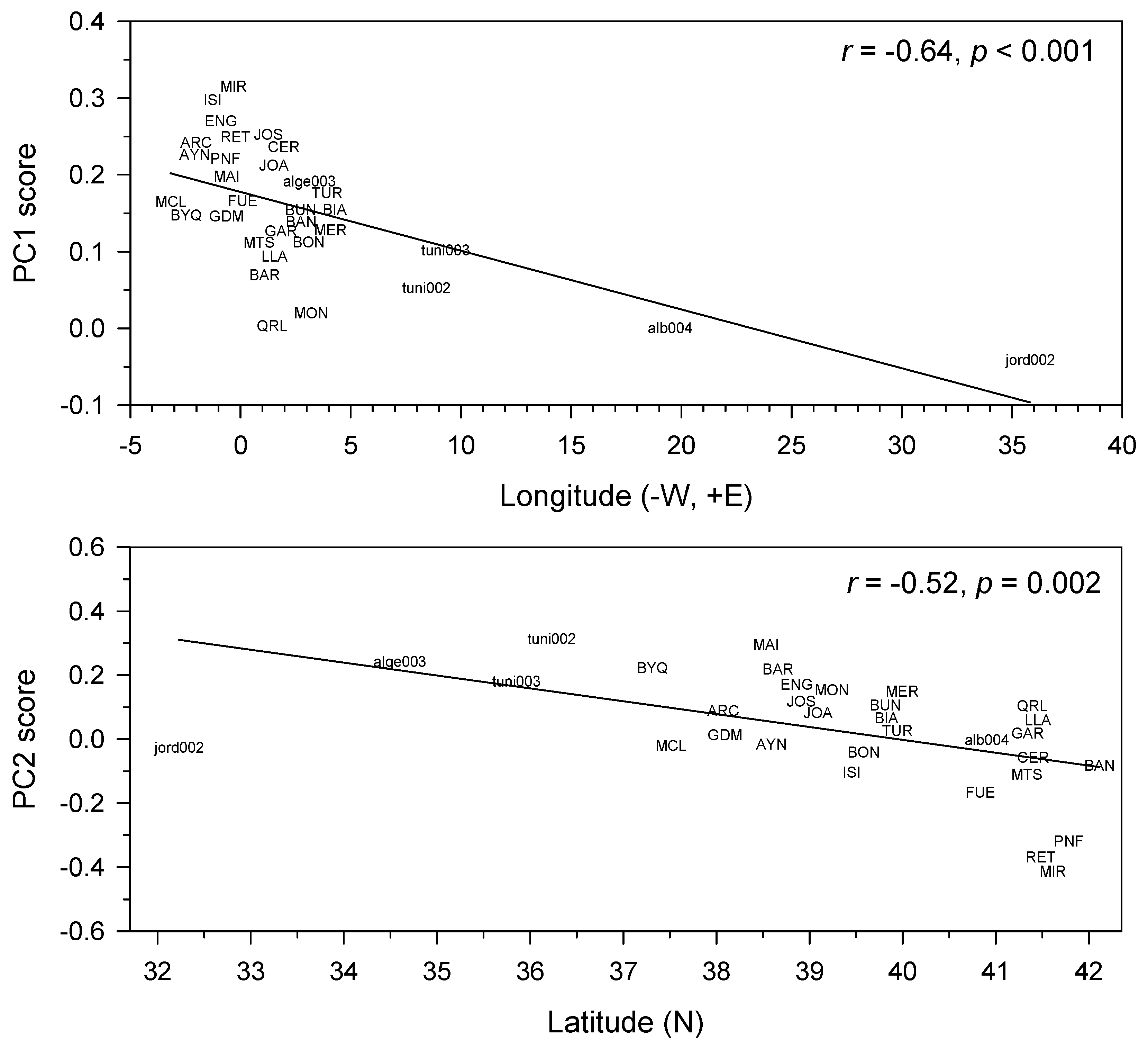
3.2. Inter-Site Distance and Common Growth Variability of Aleppo Pine Sites
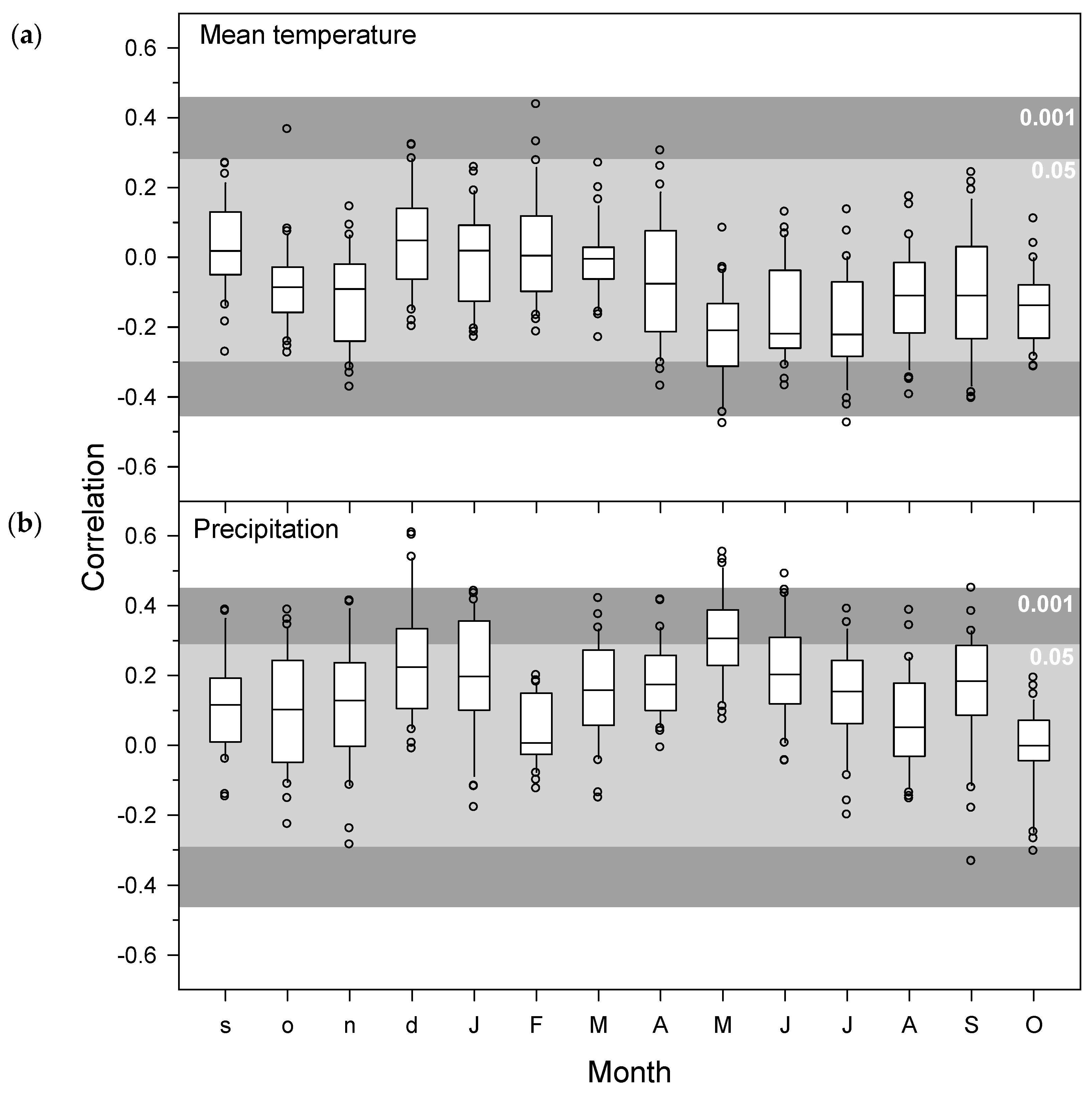
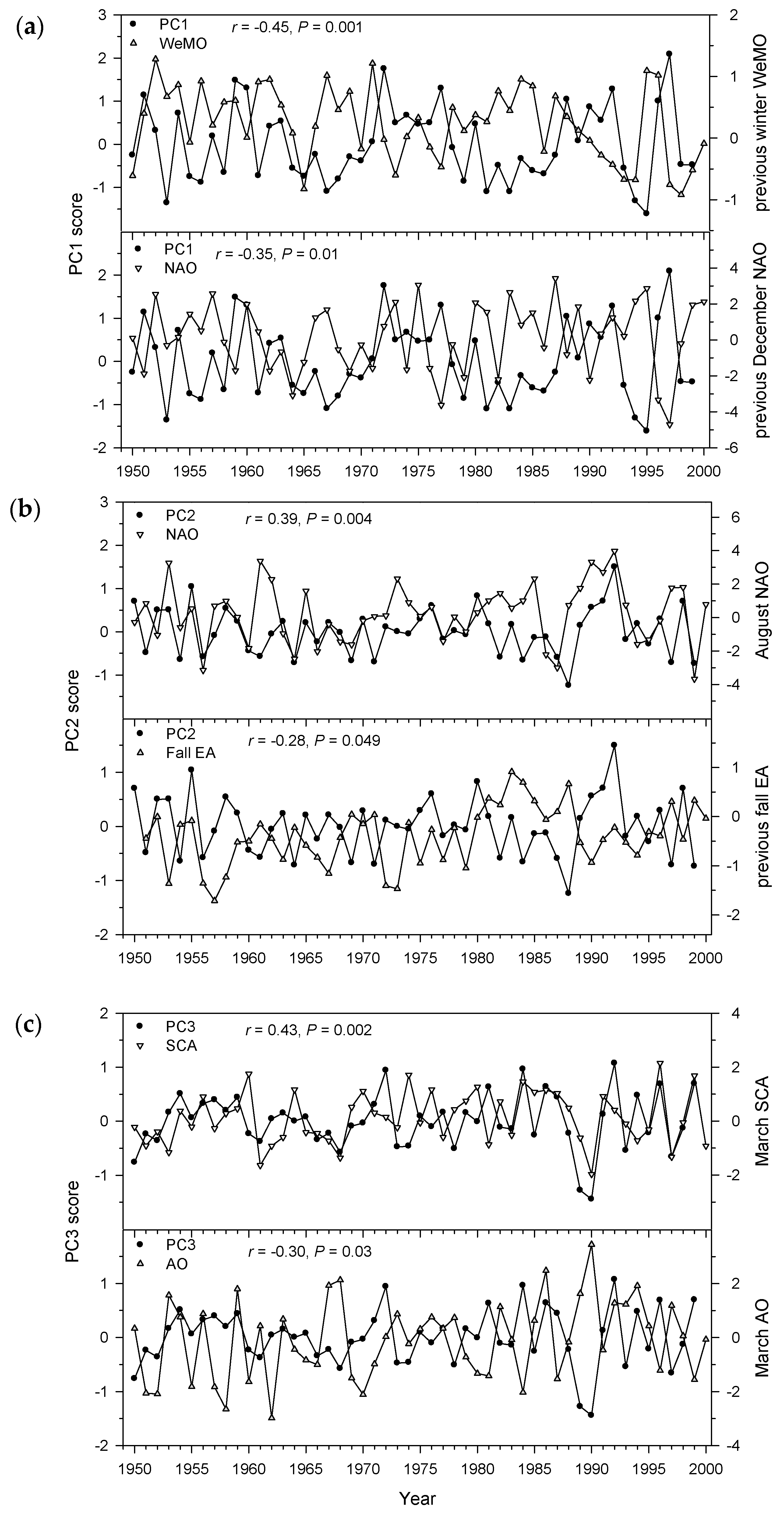
3.3. Associations of Aleppo Pine Growth versus Climate Variables and Atmospheric Patterns
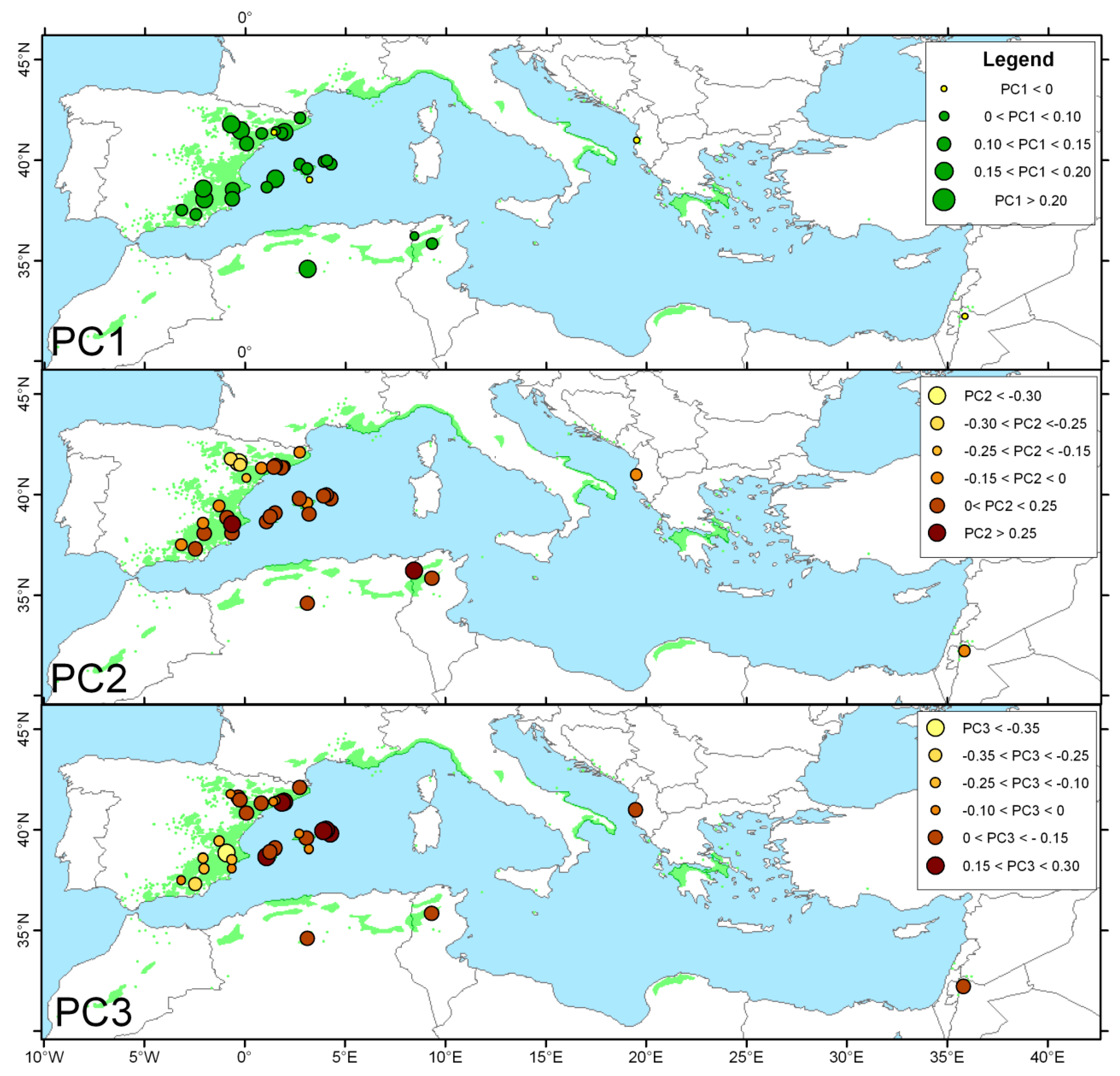
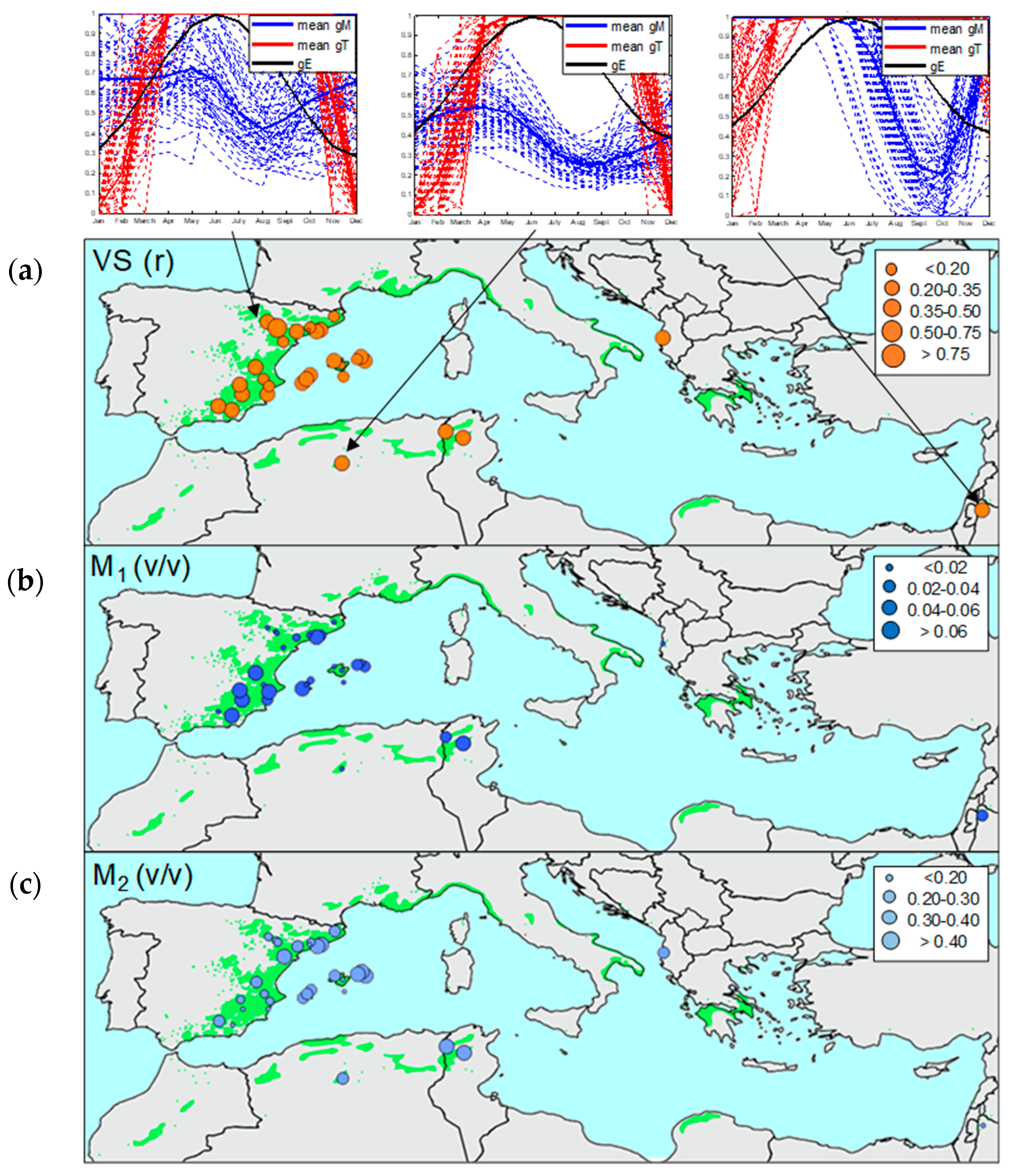
3.4. Biogeographical Climate–Growth Associations Inferred from the VS-Lite Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Touchan, R.; Anchukaitis, K.J.; Meko, D.M.; Attalah, S.; Baisan, C.; Aloui, A. Spatiotemporal drought variability in northwestern Africa over the last nine centuries. Clim. Dyn. 2011, 37, 237–252. [Google Scholar] [CrossRef]
- Dorado-Liñán, I.; Piovesan, G.; Martínez Sancho, E.; Gea-Izquierdo, G.; Zang, C.; Cañellas, I.; Castagneri, D.; Di Filippo, A.; Gutiérrez, E.; Ewald, J.; et al. Geographical adaptation prevails over species-specific determinism in trees’ vulnerability to climate change at Mediterranean rear-edge forests. Glob. Chang. Biol. 2019, 25, 1296–1314. [Google Scholar] [CrossRef] [PubMed]
- Xoplaki, E. Climate Variability over the Mediterranean. Ph.D. Thesis, University of Bern, Bern, Switzerland, 2002. [Google Scholar]
- Camarero, J.J. Direct and indirect effects of the North Atlantic Oscillation on tree growth and forest decline in northeastern Spain. In Hydrological, Socioeconomic and Ecological Impacts of the North Atlantic Oscillation in the Mediterranean Region; Advances in Global Change Research 46; Vicente-Serrano, S.M., Trigo, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 129–152. [Google Scholar]
- Seim, A.; Treydte, K.; Trouet, V.; Frank, D.; Fonti, P.; Tegel, W.; Panayotov, M.; Fernández-Donado, L.; Krusic, P.; Büntgen, U. Climate sensitivity of Mediterranean pine growth reveals distinct east-west dipole. Int. J. Clim. 2015, 35, 2503–2513. [Google Scholar] [CrossRef]
- Dorado–Liñán, I.; Zorita, E.; Martínez–Sancho, E.; Gea–Izquierdo, G.; Di Filippo, A.; Gutiérrez, E.; Levanič, T.; Piovesan, G.; Vacchiano, G.; Zang, C.; et al. Large–scale atmospheric circulation enhances the Mediterranean East–West tree growth contrast at rear–edge deciduous forests. Agr. For. Meteorol. 2017, 239, 86–95. [Google Scholar] [CrossRef]
- Sangüesa–Barreda, G.; Camarero, J.J.; Sánchez-Salguero, R.; Gutiérrez, E.; Linares, J.C.; Génova, M.; Ribas, M.; Tíscar, P.A.; López-Sáez, J.A. Droughts and climate warming desynchronize Black pine growth across the Mediterranean Basin. Sci. Total Environ. 2019, 697, 133989. [Google Scholar] [CrossRef] [PubMed]
- Touchan, R.; Xoplaki, E.; Funkhouser, G.; Luterbacher, J.; Hughes, M.K.; Erkan, N.; Akkemik, U.; Stephan, J. Reconstructions of spring/summer precipitation for the Eastern Mediterranean from tree-ring widths and its connection to large-scale atmospheric circulation. Clim. Dyn. 2005, 25, 75–98. [Google Scholar] [CrossRef]
- Ribas, M. Dendroecología de Pinus Halepensis Mill. en el este de la Península Ibérica e islas Baleares: Sensibilidad y Grado de Adaptación a las Condicionesclimáticas. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 2006. [Google Scholar]
- De Luis, M.; Čufar, K.; Di Filippo, A.; Novak, K.; Papadopoulos, A.; Piovesan, G.; Rathgeber, C.B.; Raventós, J.; Saz, M.A.; Smith, K.T. Plasticity in dendroclimatic response across the distribution range of Aleppo pine (Pinus halepensis). PLoS ONE 2013, 8, e83550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazol, A.; Ribas, M.; Gutiérrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought–induced growth decline and partial recovery. Agr. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- Del Río, M.; Rodríguez-Alonso, J.; Bravo-Oviedo, A.; Ruíz-Peinado, R.; Cañellas, I.; Gutiérrez, E. Aleppo pine vulnerability to climate stress is independent of site productivity of forest stands in southeastern Spain. Trees 2014, 28, 1209–1224. [Google Scholar] [CrossRef]
- Touchan, R.; Shishov, V.V.; Meko, D.M.; Nouiri, I.; Grachev, A. Process based model sheds light on climate sensitivity of Mediterranean tree-ring width. Biogeosciences 2012, 9, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Salguero, R.; Camarero, J.J.; Gutiérrez, E.; González Rouco, F.; Gazol, A.; Sangüesa-Barreda, G.; Andreu-Hayles, L.; Linares, J.C.; Seftigen, K. Assessing forest vulnerability to climate warming using a process-based model of tree growth: Bad prospects for rear-edges. Glob. Chang. Biol. 2017, 23, 2705–2719. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Rozas, V.; Génova, M.; Olano, J.M.; Arzac, A.; Gazol, A.; Caminero, L.; Tejedor, E.; de Luis, M.; et al. Resist, recover or both? Growth plasticity in response to drought is geographically structured and linked to intra-specific variability in Pinus pinaster. J. Biogeogr. 2018, 45, 1126–1139. [Google Scholar]
- Vaganov, E.A.; Hughes, M.K.; Shashkin, A. Growth Dynamics of Conifer Tree Rings: Images of Past and Future Environments; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Gindel, I. Cambial activity as a function of the intensity of transpiration in Pinus halepensis Mill. Proc. XVI IUFRO Congr. München 1967, 4, 188–206. [Google Scholar]
- Serre-Bachet, F. Les rapports de la croissance et du climat chez le pin d’Alep. Acta Oecol./Oecol. Plant. 1976, 11, 143–171. [Google Scholar]
- Liphschitz, N.; Lev-Yadun, S.; Rosen, E.; Waisel, Y. The annual rhythm of activity of the lateral meristems (cambium and phellogen) in Pinus halepensis Mill. and Pinus pinea L. IAWA Bull. 1984, 5, 263–274. [Google Scholar] [CrossRef]
- Lev-Yadun, S. Wood Structure and the Ecology of Annual Growth Ring Formation in Pinus halepensis and P. brutia. In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 67–78. [Google Scholar]
- Nicault, A.; Rathgeber, C.; Tessier, L.; Thomas, A. Observations sur la mise en place du cerne chez le pin d’Alep (Pinus halepensis Mill.): Confrontation entre les mesures de croissance radiale, de densité et les facteurs climatiques. Ann. For. Sci. 2001, 58, 769–784. [Google Scholar] [CrossRef] [Green Version]
- De Luis, M.; Gričar, J.; Čufar, K.; Raventós, R. Seasonal dynamics of wood formation in Pinus halepensis from dry and semi-arid ecosystems in Spain. IAWA J. 2007, 28, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- Pacheco, A.; Camarero, J.J.; Ribas, M.; Gazol, A.; Gutiérrez, E.; Carrer, M. Disentangling the climate-driven bimodal growth pattern in coastal and continental Mediterranean pine stands. Sci. Total Environ. 2018, 615, 1518–1526. [Google Scholar] [CrossRef]
- Tolwinski-Ward, S.E.; Evans, M.N.; Hughes, M.; Anchukaitis, K. An efficient forward model of the climate controls on interannual variation in tree-ring width. Clim. Dyn. 2011, 36, 2419–2439. [Google Scholar] [CrossRef]
- Allué, J.L. Atlas Fitoclimático de España; INIA-MAPA: Madrid, Spain, 1990. [Google Scholar]
- Quézel, P. Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Neeman, L., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherland, 2000; pp. 1–12. [Google Scholar]
- Schiller, G. Significance of bedrock as a site factor for Aleppo pine. For. Ecol. Manag. 1982, 4, 213–223. [Google Scholar] [CrossRef]
- Baquedano, F.J.; Valladares, F.; Castillo, F.J. Phenotypic plasticity blurs ecotypic divergence in the response of Quercus coccifera and Pinus halepensis to water stress. Eur. J. For. Res. 2008, 127, 495–506. [Google Scholar] [CrossRef]
- Ferrio, J.P.; Florit, A.; Vega, A.; Serrano, L.; Voltas, J. Isotopic signature of 13C and tree-ring width reflect different drought responses in Quercus ilex and Pinus halepensis. Oecologia 2003, 137, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Cohen, S.; Yakir, D. Hydraulic adaptations underlying drought resistance of Pinus halepensis. Tree Physiol. 2011, 31, 637–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voltas, J.; Shestakova, T.; Patsiou, T.; di Matteo, G.; Klein, T. Ecotypic variation and stability in growth performance of the thermophilic conifer Pinus halepensis across the Mediterranean basin. For. Ecol. Manag. 2018, 424, 205–215. [Google Scholar] [CrossRef]
- Esteban, L.G.; Martin, J.A.; de Palacios, P.; Fernández, F.G.; López, R. Adaptive anatomy of Pinus halepensis trees from different Mediterranean environments in Spain. Trees 2010, 24, 19–30. [Google Scholar] [CrossRef]
- Fréjaville, T.; Benito-Garzón, M. The EuMedClim database: Yearly climate data (1901–2014) of 1 km resolution grids for Europe and the Mediterranean Basin. Front. Ecol. Evol. 2018, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Tuhkanen, S. Climatic parameters and indices in plant geography. Acta Phyogeogr. Suecica 1980, 67, 1–105. [Google Scholar]
- Glueck, M.F.; Stockton, C.W. Reconstruction of the North Atlantic Oscillation, 1429–1983. Int. J. Climatol. 2001, 21, 1453. [Google Scholar] [CrossRef]
- Piovesan, G.; Schirone, B. Winter North Atlantic oscillation effects on the tree rings of the Italian beech (Fagus sylvatica L.). Int. J. Biometeorol. 2000, 44, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Yakir, D.; Lev-Yadum, S.; Zangvil, A. El Niño and tree growth near Jerusalem over the last 20 years. Glob. Chang. Biol. 1996, 2, 101–105. [Google Scholar]
- Rodó, X.; Baert, E.; Comin, F.A. Variations in seasonal rainfall in southern Europe during the present century: Relationships with the North Atlantic Oscillation and the El Niño Southern Oscillation. Clim. Dyn. 1997, 13, 275–284. [Google Scholar] [CrossRef]
- Martín-Vide, J.; López-Bustins, J.A. The Western Mediterranean Oscillation and rainfall in the Iberian Peninsula. Int. J. Climatol. 2006, 26, 1455–1475. [Google Scholar] [CrossRef]
- KNMI Climate Explorer. Available online: https://climexp.knmi.nl/ (accessed on 20 March 2020).
- WeMO. Available online: http://www.ub.edu/gc/wemo/ (accessed on 20 March 2020).
- ITRDB. Available online: https://www.ncdc.noaa.gov/data-access/paleoclimatology-data/datasets/tree-ring (accessed on 20 March 2020).
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1990, 21, 414–416. [Google Scholar] [CrossRef]
- Touchan, R.; Hughes, M. Dendrochronology in Jordan. J. Arid Environ. 1999, 42, 291–303. [Google Scholar] [CrossRef]
- Touchan, R.; Anchukaitis, K.J.; Meko, D.M.; Attalah, S.; Baisan, C.; Aloui, A. Long term context for recent drought in northwestern Africa. Geophys. Res. Lett. 2008, 35, L13705. [Google Scholar] [CrossRef]
- Toromani, E.; Pasho, E.; Alla, A.Q.; Mine, V.; Çollaku, N. Radial growth responses of Pinus halepensis Mill. and Pinus pinea L. forests to climate variability in Western Albania. Geochronometria 2015, 42, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Krusic, P. A Tree-Ring Standardization Program Based on Detrending and Autoregressive Time Series Modeling, with Interactive Graphics; Tree-Ring Laboratory, Lamont Doherty Earth Observatory, Columbia University: New York, NY, USA, 2005. [Google Scholar]
- Touchan, R.; Meko, D.M.; Aloui, A. Precipitation reconstruction for Northwestern Tunisia from tree rings. J. Arid Environ. 2008, 72, 1887–1896. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree-Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Briffa, K.R.; Jones, P.D. Basic Chronology Statistics and Assessment. In Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer Academic Publishers: Berlin, Germany, 1990; pp. 137–152. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Rolland, C. Decreasing teleconnections with inter-site distance in monthly climatic data and tree-ring width networks in a mountainous Alpine area. Theor. Appl. Climatol. 2002, 71, 63–75. [Google Scholar] [CrossRef]
- Tolwinski-Ward, S.E.; Anchukaitis, K.J.; Evans, M.N. Bayesian parameter estimation and interpretation for an intermediate model of tree-ring width. Clim. Past 2013, 9, 1481–1493. [Google Scholar] [CrossRef] [Green Version]
- Touchan, R.; Anchukaitis, K.J.; Meko, D.M.; Kerchouche, D.; Slimani, S.; Ilmen, R.; Hasnaoui, F.; Guibal, F.; Camarero, J.J.; Sánchez-Salguero, R.; et al. Climate controls on tree growth in the Western Mediterranean. Holocene 2017, 27, 1429–1442. [Google Scholar] [CrossRef]
- Novak, K.; De Luis, M.; Raventos, J.; Čufar, K. Climatic signals in tree–ring widths and wood structure of Pinus halepensis in contrasted environmental conditions. Trees 2013, 27, 927–936. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Serré-Bachet, F.; Tessier, L. Tree ring to climate relationships of Aleppo pine (Pinus halepensis Mill.) in Greece. Ecol. Mediterr. 2001, 27, 89–98. [Google Scholar] [CrossRef]
- Olivar, J.; Bogino, S.; Spiecker, H.; Bravo, F. Climate impact on growth dynamic and intra-annual density fluctuations in Aleppo Pine (Pinus halepensis) trees of different crown classes. Dendrochronologia 2012, 30, 35–47. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente–Serrano, S.M. Spatial variability in large–scale and regional atmospheric drivers of Pinus halepensis growth in eastern Spain. Agr. For. Meteorol. 2011, 151, 1106–1119. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; Vicente-Serrano, S.M. Climatic impacts and drought control of radial growth and seasonal wood formation in Pinus halepensis. Trees 2012, 26, 1875–1886. [Google Scholar] [CrossRef]
- Choury, Z.; Shestakova, T.A.; Himrane, H.; Touchan, R.; Kerchouche, D.; Camarero, J.J.; Voltas, J. Quarantining the Sahara desert: Growth and water–use efficiency of Aleppo pine in the Algerian Green Barrier. Eur. J. For. Res. 2017, 136, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Oppenheimer, H.R. Cambial wood production in stems of Pinus halepensis. Palest. J. Bot. 1945, 5, 22–51. [Google Scholar]
- Attolini, M.R.; Calvani, F.; Galli, M.; Nanni, T.; Ruggiero, L.; Schaer, E.; Zuanni, F. The relationship between climatic variables and wood structure in Pinus halepensis Mill. Theor. Appl. Climatol. 1990, 41, 121–127. [Google Scholar] [CrossRef]
- Serre-Bachet, F. Les enseignements écologiques de la variation de l’épaisseur du cerne chez le pin d’Alep. Forêt Méditerranéenne 1992, 8, 171–176. [Google Scholar]
- Mitrakos, K. A theory for Mediterranean plant life. Oecol. Plant. 1980, 1, 245–252. [Google Scholar]
- Maseyk, K.S.; Lin, T.; Rotenberg, E.; Grünzweig, J.M.; Schwartz, A.; Yakir, D. Physiology–phenology interactions in a productive semi-arid pine forest. New Phytol. 2008, 178, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Hoch, G.; Yakir, D.; Körner, C. Drought stress, growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest. Tree Physiol. 2014, 34, 981–992. [Google Scholar] [CrossRef] [Green Version]
- Touchan, R.; Kherchouche, D.; Oudjehih, B.; Touchan, H.; Slimani, S.; Meko, D.M. Dendroclimatology and wheat production in Algeria. J. Arid Environ. 2016, 124, 102–110. [Google Scholar] [CrossRef]
- Touchan, R.; Shishov, V.V.; Tychkov, I.I.; Sivrikaya, F.; Attieh, J.; Ketmen, M.; Stephan, J.; Mitsopoulos, I.; Christou, A.; Meko, D.M. Elevation-layered dendroclimatic signal in eastern Mediterranean tree rings. Environ. Res. Lett. 2016, 11, 044020. [Google Scholar] [CrossRef] [Green Version]
- Rathgeber, C.; Nicault, A.; Guiot, J.; Keller, T.; Guibal, F.; Roche, P. Simulated responses of Pinus halepensis forest productivity to climatic change and CO2 increase using a statistical model. Glob. Planet. Chang. 2000, 26, 405–421. [Google Scholar] [CrossRef]
- Sabaté, S.; Gracia, C.A.; Sánchez, A. Likely effects of climate change on growth of Quercus ilex, Pinus halepensis, Pinus pinaster, Pinus sylvestris and Fagus sylvatica forests in the Mediterranean region. For. Ecol. Manag. 2002, 162, 23–37. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Tardif, J.C.; Conciatori, F. Attributing forest responses to global-change drivers: Limited evidence of a CO2-fertilization effect in Iberian pine growth. J. Biogeogr. 2015, 42, 2220–2233. [Google Scholar] [CrossRef] [Green Version]
- Shestakova, T.A.; Camarero, J.J.; Ferrio, J.P.; Knorre, A.A.; Gutiérrez, E.; Voltas, J. Increasing drought effects on five European pines modulate Δ13C-growth coupling along a Mediterranean altitudinal gradient. Funct. Ecol. 2017, 31, 1359–1370. [Google Scholar] [CrossRef] [Green Version]
- Vennetier, M.; Vila, B.; Liang, E.Y.; Guibal, F.; Thabeet, A.; Gadbin-Henry, C. Impact of climate change on pine forest productivity and on the shift of a bioclimatic limit in a Mediterranean area. Options Méditerranéenne Séries A 2007, 75, 189–197. [Google Scholar]
- Hevia, A.; Campelo, F.; Chambel, R.; Vieira, J.; Alía, R.; Majada, J.; Sánchez-Salguero, R. Which matters more for wood traits in Pinus halepensis Mill., provenance or climate? Ann. For. Sci. 2020. [Google Scholar] [CrossRef]
- Alfaro Sánchez, R.; Camarero, J.J.; López-Serrano, F.R.; Sánchez Salguero, R.; Moya, D.; de las Heras, J. Positive coupling between growth and reproduction in young post-fire Aleppo pines depends on climate and site conditions. Int. J. Wildland Fire 2015, 24, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Preisler, Y.; Tatarinov, F.; Grünzweig, J.M.; Bert, D.; Ogée, J.; Wingate, L.; Rotenberg, E.; Rohatyn, S.; Her, N.; Moshe, I.; et al. Mortality versus survival in drought-affected Aleppo pine forest depends on the extent of rock cover and soil stoniness. Funct. Ecol. 2019, 33, 901–912. [Google Scholar] [CrossRef]
- Borghetti, M.; Cinnirella, S.; Magnani, F.; Saracino, A. Impact of long-term drought on xylem embolism and growth in Pinus halepensis Mill. Trees 1998, 12, 187–195. [Google Scholar] [CrossRef]
- Oliveras, I.; Martínez-Vilalta, J.; Jimenez-Ortiz, T.; Lledó, M.J.; Escarré, A.; Piñol, J. Hydraulic properties of Pinus halepensis, Pinus pinea and Tetraclinis articulata in a dune ecosystem of Eastern Spain. Plant Ecol. 2003, 169, 131–141. [Google Scholar] [CrossRef]
- Girard, F.; Vennetier, M.; Guibal, F.; Corona, C.; Ouarmim, S.; Herrero, A. Pinus halepensis Mill. crown development and fruiting declined with repeated drought in Mediterranean France. Eur. J. Forest. Res. 2012, 131, 919–931. [Google Scholar] [CrossRef] [Green Version]
- Sarris, D.; Christodoulakis, D.; Körner, C. Recent decline in precipitation and tree growth in the eastern Mediterranean. Glob. Chang. Biol. 2007, 13, 1187–1200. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerillo, R.M.; Camarero, J.J.; Fernández-Cancio, A. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early-warning signals of dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef] [Green Version]
- De la Serrana, R.G.; Vilagrosa, A.; Alloza, J.A. Pine mortality in southeast Spain after an extreme dry and warm year: Interactions among drought stress, carbohydrates and bark beetle attack. Trees 2015, 29, 1791–1804. [Google Scholar] [CrossRef]
- De Luis, M.; Novak, K.; Čufar, K.; Raventós, J. Size mediated climate–growth relationships in Pinus halepensis and Pinus pinea. Trees 2009, 23, 1065–1073. [Google Scholar] [CrossRef]
- Dorman, M.; Perevolotsky, A.; Sarris, D.; Svoray, T. The effect of rainfall and competition intensity on forest response to drought: Lessons learned from a dry extreme. Oecologia 2015, 177, 1025–1038. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (Country) | Code | Latitude N (°) | Longitude −W/+E (°) | Elevation (m a.s.l) | MAT (°C) | MAP (mm) | AI |
|---|---|---|---|---|---|---|---|
| Sierra de Archivel (Spain) | ARC | 32.092 | −2.033 | 1086 | 13.6 | 361 | 15.3 |
| Ayna Los Luisos (Spain) | AYN | 38.594 | −2.079 | 1000 | 14.5 | 466 | 19.0 |
| Banyoles (Spain) | BAN | 42.107 | 2.739 | 260 | 14.8 | 804 | 32.4 |
| Es Cap de Barbaria (Spain) | BAR | 38.657 | 1.077 | 80 | 18.8 | 364 | 12.6 |
| Cala Biniancolla (Spain) | BIA | 39.816 | 4.261 | 50 | 16.9 | 601 | 22.3 |
| Santuari de Bonany (Spain) | BON | 39.582 | 3.076 | 200 | 17.5 | 594 | 21.6 |
| Comuna de Bunyola (Spain) | BUN | 39.808 | 2.705 | 500 | 17.4 | 937 | 34.2 |
| Pinar de Bayarque (Spain) | BYQ | 37.313 | −2.465 | 1280 | 12.6 | 347 | 15.4 |
| Sta. Maria de Cervelló (Spain) | CER | 41.397 | 1.958 | 300 | 13.1 | 664 | 28.7 |
| Serra d’Enguera (Spain) | ENG | 38.857 | −0.897 | 800 | 16.1 | 562 | 21.5 |
| Fuentespalda (Spain) | FUE | 40.825 | 0.083 | 850 | 12.8 | 639 | 28.0 |
| P.N. del Garraf (Spain) | GAR | 41.341 | 1.844 | 300 | 13.2 | 673 | 29.0 |
| Guardamar del Segura (Spain) | GDM | 38.092 | −0.649 | 10 | 17.7 | 293 | 10.6 |
| Los Isidros (Spain) | ISI | 39.450 | −1.275 | 635 | 13.1 | 450 | 19.5 |
| Cala Xarraca (Spain) | JOA | 39.099 | 1.511 | 175 | 17.9 | 462 | 16.6 |
| Sa Talaia de St. Josep (Spain) | JOS | 38.909 | 1.256 | 360 | 17.8 | 412 | 14.8 |
| Serra d’Ancosa (Spain) | LLA | 41.450 | 1.533 | 730 | 12.3 | 607 | 27.2 |
| Serra de Maigmó (Spain) | MAI | 38.525 | −0.636 | 900 | 14.9 | 351 | 14.1 |
| El Mencal (Spain) | MCL | 37.514 | −3.173 | 1150 | 14.3 | 357 | 14.7 |
| Es Mercadal (Spain) | MER | 39.999 | 4.063 | 200 | 17.5 | 606 | 22.0 |
| Miramón (Spain) | MIR | 41.615 | −0.331 | 500 | 14.7 | 376 | 15.2 |
| P.N. de Mondragó (Spain) | MON | 39.035 | 3.191 | 60 | 16.4 | 439 | 16.6 |
| Serra de Montsant (Spain) | MTS | 41.329 | 0.820 | 775 | 13.2 | 670 | 28.9 |
| Vedado de Peñaflor (Spain) | PNF | 41.779 | −0.709 | 350 | 14.0 | 416 | 17.3 |
| Querol (Spain) | QRL | 41.387 | 1.433 | 770 | 13.4 | 493 | 21.1 |
| La Retuerta de Pina (Spain) | RET | 41.480 | −0.242 | 386 | 15.8 | 428 | 16.6 |
| Cala Turqueta (Spain) | TUR | 39.938 | 3.916 | 63 | 17.5 | 585 | 21.3 |
| Tobji * (Algeria) | alge003 | 34.600 | 3.117 | 1380 | 13.4 | 338 | 14.1 |
| Dahllia * (Tunisia) | tuni002 | 36.233 | 8.433 | 950 | 18.0 | 434 | 15.5 |
| Jebnoun * (Tunisia) | tuni003 | 35.510 | 9.180 | 800 | 17.3 | 222 | 8.1 |
| Divjakë * (Albania) | alb004 | 41.000 | 19.483 | 3 | 15.9 | 1027 | 39.6 |
| Dibeen * (Jordan) | jord002 | 32.140 | 35.490 | 800 | 16.6 | 350 | 13.2 |
| Tree-Ring Width Data | Statistics of the VS-Lite Model | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | No. Trees/No. Cores | Mean Width ± SD (mm) | Best-Replicated Period | MS | VP1 (%) | SNR | T1 (°C) | T2 (°C) | M1 (v/v) | M2 (v/v) | r |
| ARC | 15/31 | 0.42 ± 0.39 | 1889–2001 | 0.41 | 72.7 | 28.5 | 5.60 | 13.83 | 0.09 | 0.17 | 0.62 |
| AYN | 15/30 | 0.53 ± 0.27 | 1862–1999 | 0.29 | 53.9 | 14.6 | 5.11 | 13.03 | 0.09 | 0.30 | 0.63 |
| BAN | 15/30 | 2.51 ± 0.73 | 1932–1999 | 0.34 | 59.1 | 15.0 | 8.47 | 12.16 | 0.01 | 0.39 | 0.45 |
| BAR | 15/30 | 1.43 ± 0.72 | 1873–2001 | 0.26 | 61.0 | 16.2 | 3.93 | 10.82 | 0.07 | 0.38 | 0.52 |
| BIA | 12/24 | 1.57 ± 0.74 | 1940–2001 | 0.27 | 61.8 | 13.9 | 3.91 | 15.77 | 0.06 | 0.46 | 0.67 |
| BON | 13/26 | 0.78 ± 0.24 | 1911–2003 | 0.37 | 52.6 | 8.6 | 7.58 | 11.85 | 0.00 | 0.13 | 0.12 |
| BUN | 15/25 | 1.27 ± 0.32 | 1864–2003 | 0.29 | 56.8 | 16.5 | 7.07 | 22.07 | 0.01 | 0.34 | 0.65 |
| BYQ | 15/29 | 0.99 ± 0.42 | 1902–1999 | 0.38 | 73.3 | 37.6 | 7.33 | 11.96 | 0.07 | 0.17 | 0.51 |
| CER | 13/28 | 1.43 ± 0.64 | 1916–2001 | 0.38 | 58.2 | 13.1 | 3.86 | 11.33 | 0.06 | 0.50 | 0.68 |
| ENG | 14/28 | 0.60 ± 0.36 | 1860–2000 | 0.49 | 62.3 | 17.2 | 7.35 | 21.71 | 0.01 | 0.28 | 0.46 |
| FUE | 16/31 | 0.91 ± 0.40 | 1907–1999 | 0.29 | 57.2 | 17.9 | 8.09 | 15.71 | 0.01 | 0.42 | 0.50 |
| GAR | 22/43 | 0.95 ± 0.25 | 1938–2003 | 0.28 | 55.1 | 20.7 | 3.94 | 22.03 | 0.07 | 0.44 | 0.65 |
| GDM | 15/28 | 0.73 ± 0.38 | 1915–2000 | 0.24 | 46.5 | 10.8 | 5.45 | 19.59 | 0.05 | 0.15 | 0.59 |
| ISI | 15/30 | 1.05 ± 0.40 | 1920–1999 | 0.32 | 62.5 | 19.0 | 7.34 | 11.19 | 0.08 | 0.40 | 0.74 |
| JOA | 16/31 | 1.60 ± 0.77 | 1915–2001 | 0.35 | 65.4 | 21.4 | 8.16 | 12.07 | 0.03 | 0.35 | 0.70 |
| JOS | 15/28 | 1.57 ± 0.65 | 1920–2001 | 0.36 | 68.2 | 22.6 | 8.26 | 13.49 | 0.01 | 0.36 | 0.72 |
| LLA | 16/27 | 1.70 ± 0.52 | 1949–1999 | 0.21 | 48.2 | 11.8 | 6.00 | 13.22 | 0.00 | 0.27 | 0.43 |
| MAI | 13/30 | 0.79 ± 0.38 | 1895–1999 | 0.45 | 59.8 | 14.1 | 5.01 | 21.18 | 0.08 | 0.26 | 0.37 |
| MCL | 13/24 | 1.58 ± 0.58 | 1893–1999 | 0.31 | 62.9 | 17.6 | 5.66 | 10.76 | 0.00 | 0.35 | 0.55 |
| MER | 11/22 | 2.86 ± 1.30 | 1926–2001 | 0.29 | 66.8 | 13.8 | 7.01 | 12.34 | 0.04 | 0.47 | 0.52 |
| MIR | 19/38 | 0.68 ± 0.44 | 1840–2004 | 0.41 | 54.9 | 18.4 | 6.90 | 11.66 | 0.00 | 0.24 | 0.75 |
| MON | 15/30 | 1.57 ± 0.60 | 1909–2003 | 0.32 | 52.1 | 11.5 | 6.51 | 19.29 | 0.02 | 0.19 | 0.34 |
| MTS | 15/30 | 1.15 ± 0.31 | 1919–1999 | 0.24 | 48.8 | 10.2 | 8.45 | 13.29 | 0.03 | 0.34 | 0.60 |
| PNF | 23/43 | 0.74 ± 0.34 | 1860–2004 | 0.44 | 69.8 | 35.3 | 6.61 | 11.14 | 0.01 | 0.27 | 0.67 |
| QRL | 15/30 | 2.04 ± 0.60 | 1948–1999 | 0.17 | 48.0 | 11.8 | 5.84 | 11.34 | 0.01 | 0.14 | 0.16 |
| RET | 12/43 | 0.85 ± 0.36 | 1857–2004 | 0.39 | 70.2 | 26.0 | 8.07 | 12.07 | 0.02 | 0.25 | 0.76 |
| TUR | 10/21 | 1.53 ± 0.67 | 1909–2001 | 0.25 | 50.1 | 10.0 | 4.58 | 15.99 | 0.06 | 0.41 | 0.44 |
| alge003 | 20/39 | 0.99 ± 0.70 | 1887–2006 | 0.44 | 71.3 | 44.4 | 7.23 | 12.65 | 0.00 | 0.40 | 0.74 |
| tuni002 | 11/19 | 1.60 ± 0.99 | 1912–2001 | 0.37 | 55.8 | 8.8 | 4.35 | 13.82 | 0.06 | 0.44 | 0.70 |
| tuni003 | 14/25 | 1.35 ± 0.81 | 1933–2001 | 0.38 | 59.6 | 17.0 | 2.77 | 11.82 | 0.07 | 0.44 | 0.67 |
| alb004 | 20/40 | 3.99 ± 1.81 | 1968–2008 | 0.23 | 32.9 | 7.5 | 4.44 | 16.75 | 0.01 | 0.38 | 0.52 |
| jord002 | 14/14 | 1.92 ± 1.16 | 1940–1994 | 0.35 | 45.7 | 7.0 | 6.40 | 10.93 | 0.05 | 0.13 | 0.73 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camarero, J.J.; Sánchez-Salguero, R.; Ribas, M.; Touchan, R.; Andreu-Hayles, L.; Dorado-Liñán, I.; Meko, D.M.; Gutiérrez, E. Biogeographic, Atmospheric, and Climatic Factors Influencing Tree Growth in Mediterranean Aleppo Pine Forests. Forests 2020, 11, 736. https://doi.org/10.3390/f11070736
Camarero JJ, Sánchez-Salguero R, Ribas M, Touchan R, Andreu-Hayles L, Dorado-Liñán I, Meko DM, Gutiérrez E. Biogeographic, Atmospheric, and Climatic Factors Influencing Tree Growth in Mediterranean Aleppo Pine Forests. Forests. 2020; 11(7):736. https://doi.org/10.3390/f11070736
Chicago/Turabian StyleCamarero, J. Julio, Raúl Sánchez-Salguero, Montserrat Ribas, Ramzi Touchan, Laia Andreu-Hayles, Isabel Dorado-Liñán, David M. Meko, and Emilia Gutiérrez. 2020. "Biogeographic, Atmospheric, and Climatic Factors Influencing Tree Growth in Mediterranean Aleppo Pine Forests" Forests 11, no. 7: 736. https://doi.org/10.3390/f11070736
APA StyleCamarero, J. J., Sánchez-Salguero, R., Ribas, M., Touchan, R., Andreu-Hayles, L., Dorado-Liñán, I., Meko, D. M., & Gutiérrez, E. (2020). Biogeographic, Atmospheric, and Climatic Factors Influencing Tree Growth in Mediterranean Aleppo Pine Forests. Forests, 11(7), 736. https://doi.org/10.3390/f11070736