Marker-Assisted Selection of Trees with MALE STERILITY 1 in Cryptomeria japonica D. Don

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotyping of Male Sterility and Single Nucleotide Polymorphism (SNP) Genotyping for Linkage Analysis
2.2. MAS of Trees with ms1
3. Results and Discussion
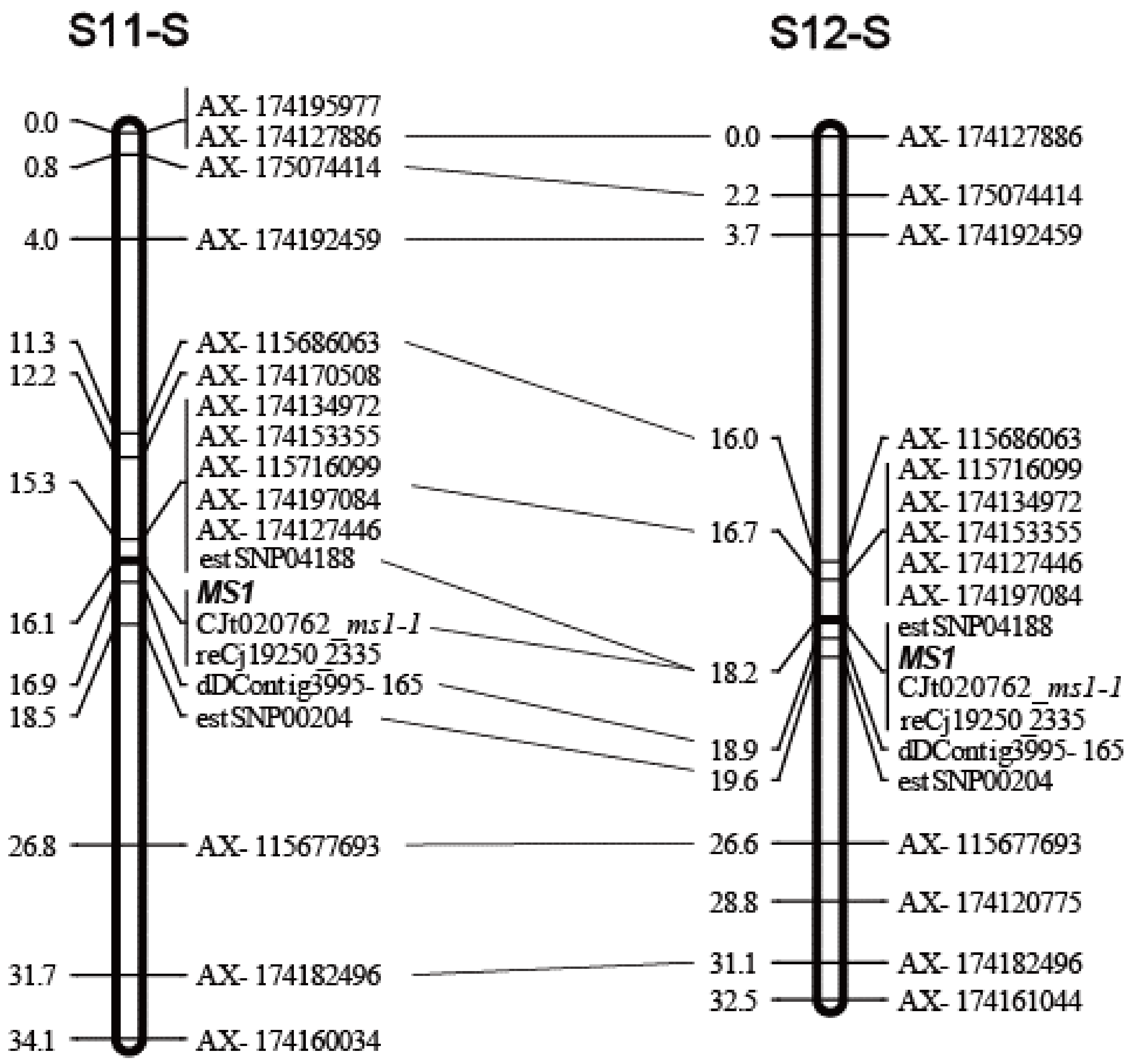
3.1. Linkage Maps of the MS1 Region
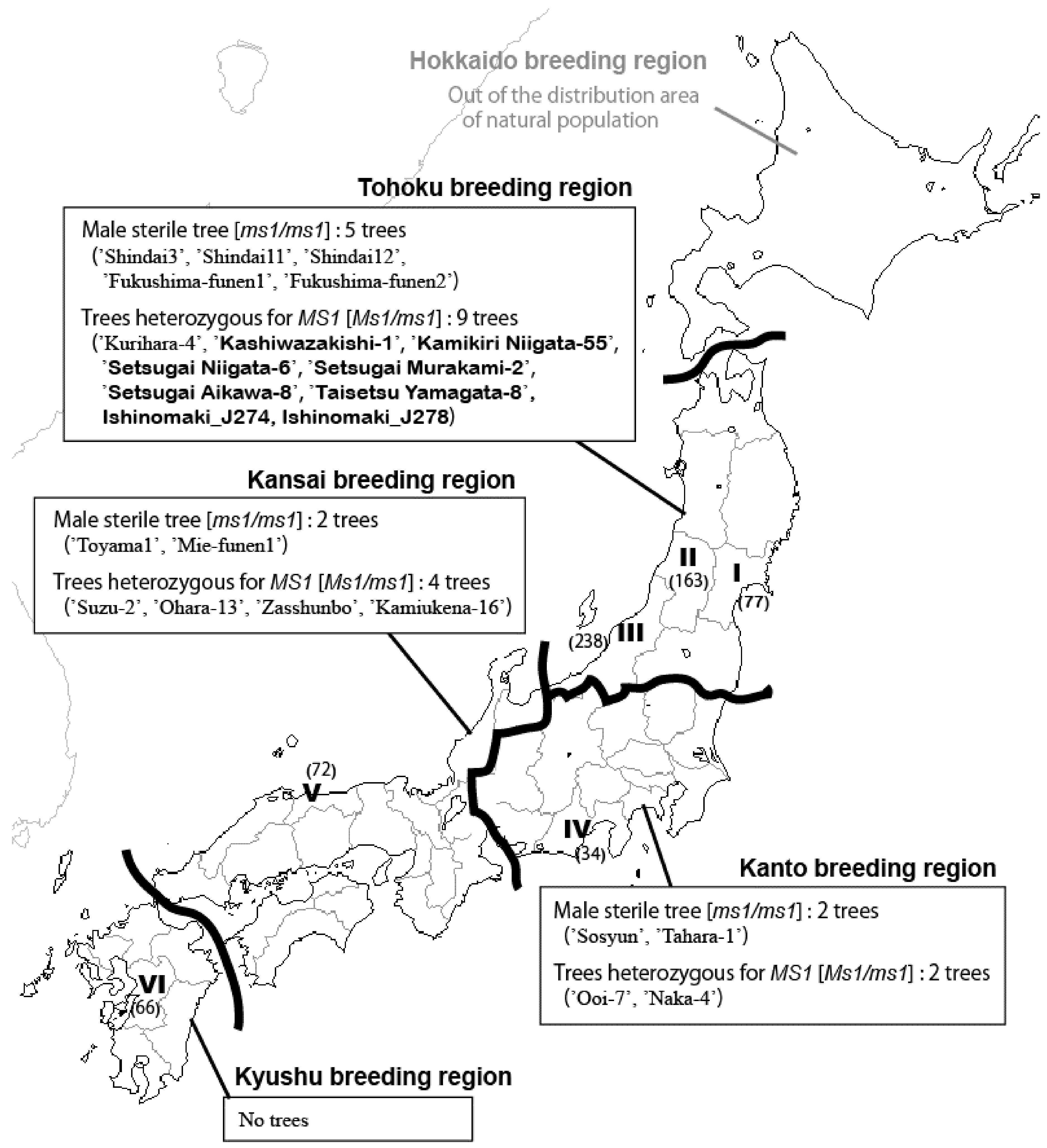
3.2. MAS of Trees with ms1
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. 2011, 12, 111–122. [Google Scholar] [CrossRef]
- Forestry Agency. Statistical Handbook of Forest and Forestry; Forestry Agency, Ministry of Agriculture, Forestry and Fisheries: Tokyo, Japan, 2014; pp. 8–9. (In Japanese) [Google Scholar]
- Baba, K.; Nakae, K. The national epidemiological survey of nasal allergy 2008 (compared with 1998) in otolaryngologists and their family members. Prog. Med. 2008, 28, 2001–2012. (In Japanese) [Google Scholar]
- Taira, H.; Teranishi, H.; Kenda, Y. A case study of male sterility in sugi (Cryptomeria japonica). J. Jpn. For. Soc. 1993, 75, 377–379, (In Japanese with English summary). [Google Scholar]
- Taira, H.; Saito, M.; Furuta, Y. Inheritance of the trait of male sterility in Cryptomeria japonica. J. For. Res. 1999, 4, 271–273. [Google Scholar] [CrossRef]
- Saito, M. Breeding strategy for the pollinosis preventive cultivars of Cryptomeria japonica D Don. J. Jpn. For. Soc. 2010, 92, 316–323, (In Japanese with English summary). [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, Y.; Ueno, S.; Wei, F.J.; Matsumoto, A.; Uchiyama, K.; Ujino-Ihara, T.; Hakamata, T.; Fujino, T.; Kasahara, M.; Bino, T.; et al. Identification and genetic diversity analysis of a male-sterile gene (MS1) in Japanese cedar (Cryptomeria japonica D. Don). Sci. Rep. 2020. under review. [Google Scholar] [CrossRef]
- Wilson, Z.A.; Morroll, S.M.; Dawson, J.; Swarup, R.; Tighe, P. The Arabidopsis MALE STERILITY1 (MS1) gene is a transcriptional regulator of male gametogenesis, with homology to the PHD-finger family of transcription factors. Plant J. 2001, 28, 27–39. [Google Scholar] [CrossRef]
- Igarashi, M.; Watanabe, J.; Ozawa, H.; Saito, Y.; Taira, H. The male sterile sugi (Cryptomeria japonica D. Don) was found in Fukushima Prefecture (I): Selection of the search ground and identification of the male sterility. Tohoku J. For. Sci. 2004, 9, 86–89, (In Japanese with English abstract). [Google Scholar]
- Takahashi, M.; Iwaizumi, M.G.; Hoshi, H.; Kubota, M.; Fukuda, Y.; Fukatsu, E.; Kondo, T. Survey of male sterility on sugi (Cryptomeria japonica D. Don) clones collected from Kanto Breeding Region and characteristics of two male-sterile clones. Bull. Natl. For. Tree Breed. Cent. 2007, 23, 11–36, (In Japanese with English abstract). [Google Scholar]
- Fujisawa, T.; Saito, H.; Fujimiya, T.; Taira, H.; Saito, M. Selection and practical application of male-fertile cedar from elite trees in Kanagawa prefecture. In Proceedings of the 120th Japanese Forest Society Meeting, Kyoto, Japan, 26 March 2009. Abstract Number Pb01-06 (In Japanese). [Google Scholar]
- Ueuma, H.; Yoshii, E.; Hosoo, Y.; Taira, H. Cytological study of a male-sterile Cryptomeria japonica that does not release microspores from tetrads. J. For. Res. 2009, 14, 123–126. [Google Scholar] [CrossRef]
- Yamada, H.; Yamaguchi, K. Survey of male sterility of sugi (Cryptomeria japonica D. Don) clones collected from Kansai Breeding Region and characteristics of the male-sterile clone. Appl. For. Sci. 2009, 8, 33–36. (In Japanese) [Google Scholar]
- Kawai, K.; Kubota, M.; Endo, K.; Isoda, K. Trial of efficient method for screening Cryptomeria japonica trees heterozygous for male-sterile gene by segregation of male-sterile seedlings derived from self-pollinated progeny. Bull. FFPRI 2017, 444, 265–266. (In Japanese) [Google Scholar]
- Yoshii, E.; Taira, H. Cytological and genetical studies on male sterile sugi (Cryptomeria japonica D. Don), Shindai 1 and Shindai 5. J. Jpn. For. Soc. 2007, 89, 26–30, (In Japanese with English summary). [Google Scholar] [CrossRef] [Green Version]
- Miyajima, D.; Yoshii, E.; Hosoo, Y.; Taira, H. Cytological and genetic studies on male sterility in Cryptomeria japonica D. Don (Shindai 8). J. Jpn. For. Soc. 2010, 92, 106–109, (In Japanese with English summary). [Google Scholar] [CrossRef] [Green Version]
- Saito, M.; Taira, H.; Furuta, Y. Cytological and genetical studies on male sterility in Cryptomeria japonica D. Don. J. For. Res. 1998, 3, 167–173. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Ujino-Ihara, T.; Uchiyama, K.; Futamura, N.; Saito, M.; Ueno, S.; Matsumoto, A.; Tani, N.; Taira, H.; Shinohara, K.; et al. The construction of a high-density linkage map for identifying SNP markers that are tightly linked to a nuclear-recessive major gene for male sterility in Cryptomeria japonica D. Don. BMC Genom. 2012, 13, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, Y.; Ueno, S.; Higuchi, Y.; Miyajima, D.; Itoo, S.; Futamura, N.; Shinohara, K.; Tsumura, Y. Establishment of a microsatellite panel covering the sugi (Cryptomeria japonica) genome, and its application for localization of a male sterile gene (ms-2). Mol. Breed. 2014, 33, 315–325. [Google Scholar] [CrossRef]
- Isoda, K.; Kawai, K.; Yamaguchi, K.; Kubota, M.; Yamada, H. Screening of trees heterozygous for male-sterile gene in Kansai breeding region. Ann. Rep. Tree Breed. Cent. 2013, 2013, 55–59. [Google Scholar]
- Saito, M.; Aiura, H.; Kato, A.; Matsuura, T. Characterization of the cutting cultivar of Cryptomeria japonica D. Don, Zasshunbo, with a heterozygous male-sterility gene in a heterozygous state, selected from Toyama Prefecture. Jpn. For. Genet. Tree Breed. 2015, 4, 45–51, (In Japanese with English abstract). [Google Scholar]
- Moriguchi, Y.; Uchiyama, K.; Ueno, S.; Ujino-Ihara, T.; Matsumoto, A.; Iwai, J.; Miyajima, D.; Saito, M.; Sato, M.; Tsumura, Y. A high-density linkage map with 2,560 markers and its application for the localization of the male-sterile genes ms3 and ms4 in Cryptomeria japonica D. Don. Tree Genet. Genomes 2016, 12, 57. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Ueno, S.; Matsumoto, A.; Ujino-Ihara, T.; Uchiyama, K.; Totsuka, S.; Iwai, J.; Hakamata, T.; Moriguchi, Y. Fine mapping of the male-sterile genes (MS1, MS2, MS3, and MS4) and development of SNP markers for marker-assisted selection in Japanese cedar (Cryptomeria japonica D. Don). PLoS ONE 2018, 13, e0206695. [Google Scholar] [CrossRef] [PubMed]
- Mishima, K.; Hirao, T.; Tsubomura, M.; Tamura, M.; Kurita, M.; Nose, M.; Hanaoka, S.; Takahashi, M.; Watanabe, A. Identification of novel putative causative genes and genetic marker for male sterility in Japanese cedar (Cryptomeria japonica D. Don). BMC Genom. 2018, 19, 277. [Google Scholar] [CrossRef] [PubMed]
- Ueno, S.; Uchiyama, K.; Moriguchi, Y.; Ujino-Ihara, T.; Matsumoto, A.; Fu-Jin, W.; Saito, M.; Higuchi, Y.; Futamura, N.; Kanamori, H.; et al. Scanning RNA-Seq and RAD-Seq approach to develop SNP markers closely linked to MALE STERILITY 1 (MS1) in Cryptomeria japonica D. Don. Breed. Sci. 2019, 69, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, Y.; Ueno, S.; Fu-Jin, W.; Matsumoto, A.; Ujino-Ihara, T.; Uchiyama, K.; Moriguchi, Y.; Kasahara, M.; Fujino, T.; Shigenobu, S.; et al. Development of diagnostic PCR and LAMP markers for MALE STERILITY 1 (MS1) in Cryptomeria japonica D Don. BMC Res. Note 2020. under review. [Google Scholar] [CrossRef]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- Moriguchi, Y.; Ueno, S.; Saito, M.; Higuchi, Y.; Miyajima, D.; Itoo, S.; Tsumura, Y. A simple allele-specific PCR marker for identifying male-sterile trees: Towards DNA marker-assisted selection Cryptomeria japonica breeding program. Tree Genet. Genomes 2014, 10, 1069–1077. [Google Scholar] [CrossRef]
- Van Ooijen, J.W.; Voorrips, R.E. JoinMap, Version 3.0: Software for the Calculation of Genetic Linkage Maps; University and Research Center: Wageningen, The Netherlands, 2001. [Google Scholar]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1944, 12, 172–175. [Google Scholar] [CrossRef]
- Doğan, G.; Tadama, T.; Kohama, H.; Matsumoto, A.; Moriguchi, Y. Evidence of clonal propagation in Cryptomeria japonica D. Don distributed on Pacific Ocean side in Japan. Silvae Genet. 2017, 66, 43–46. [Google Scholar] [CrossRef] [Green Version]
- Miura, S.; Nameta, M.; Yamamoto, T.; Igarashi, M.; Taira, H. Mechanisms of male sterility in four Cryptomeria japonica individuals with obvious visible abnormality at the tetrad stage. J. Jpn. For. Soc. 2011, 93, 1–7, (In Japanese with English summary). [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriguchi, Y.; Ueno, S.; Hasegawa, Y.; Tadama, T.; Watanabe, M.; Saito, R.; Hirayama, S.; Iwai, J.; Konno, Y. Marker-Assisted Selection of Trees with MALE STERILITY 1 in Cryptomeria japonica D. Don. Forests 2020, 11, 734. https://doi.org/10.3390/f11070734
Moriguchi Y, Ueno S, Hasegawa Y, Tadama T, Watanabe M, Saito R, Hirayama S, Iwai J, Konno Y. Marker-Assisted Selection of Trees with MALE STERILITY 1 in Cryptomeria japonica D. Don. Forests. 2020; 11(7):734. https://doi.org/10.3390/f11070734
Chicago/Turabian StyleMoriguchi, Yoshinari, Saneyoshi Ueno, Yoichi Hasegawa, Takumi Tadama, Masahiro Watanabe, Ryunosuke Saito, Satoko Hirayama, Junji Iwai, and Yukinori Konno. 2020. "Marker-Assisted Selection of Trees with MALE STERILITY 1 in Cryptomeria japonica D. Don" Forests 11, no. 7: 734. https://doi.org/10.3390/f11070734
APA StyleMoriguchi, Y., Ueno, S., Hasegawa, Y., Tadama, T., Watanabe, M., Saito, R., Hirayama, S., Iwai, J., & Konno, Y. (2020). Marker-Assisted Selection of Trees with MALE STERILITY 1 in Cryptomeria japonica D. Don. Forests, 11(7), 734. https://doi.org/10.3390/f11070734