4.1. Temperature Experiment
In the present study temperatures higher than used in standard routine were applied (40, 50, and 60 °C) during the initiation stage of
P. halepensis SE for different periods (4 h, 30 min, and 5 min, respectively). No statistically significant differences could be found between initiation, proliferation, maturation or germination rates obtained in the different treatments assayed during the induction of EMs. These results are in agreement with the ones obtained when the same temperatures and induction times were studied in
P. radiata [
43] where no significant differences were also found. Nonetheless, the control presented a lower initiation rate when compared to higher temperatures, in contrast with the results obtained in previous studies carried out in our laboratory [
29].
Also, the ECLs initiated under different temperatures did not show significant differences regarding their capacity to produce SES, corroborating the results found in
Pinus spp. previous studies [
29,
43]. However, samples induced at 60 °C for 5 min showed significantly higher number of somatic embryos, doubling the number of embryos produced in samples cultured at 40 or 50 °C. Similar results have been obtained in
P. radiata, where although not statistically significant differences were found, the highest embryo production was achieved at 60 °C [
43]. Similar experiments carried out in
P. pinaster [
44] showed that an increase of temperature up to 28 °C resulted on initiation rates similar to those observed in standard conditions but significantly higher number of SES were produced. This fact reinforce the idea that temperature exerts a selective pressure in the SE initial stages, that can result in an initiation decrease but higher rates for the forthcoming steps [
45,
46].
It seems that the temperature treatment that triggered a deeper effect on the efficiency of
P. halepensis SE, was 60 °C for 5 min. In spite of the association of stress with adaptation mechanisms and the induction of SES formation, cell responses to the applied stresses depends on its intensity [
47]. The different induction times for each temperature could in a way affect samples response to the different high temperatures applied.
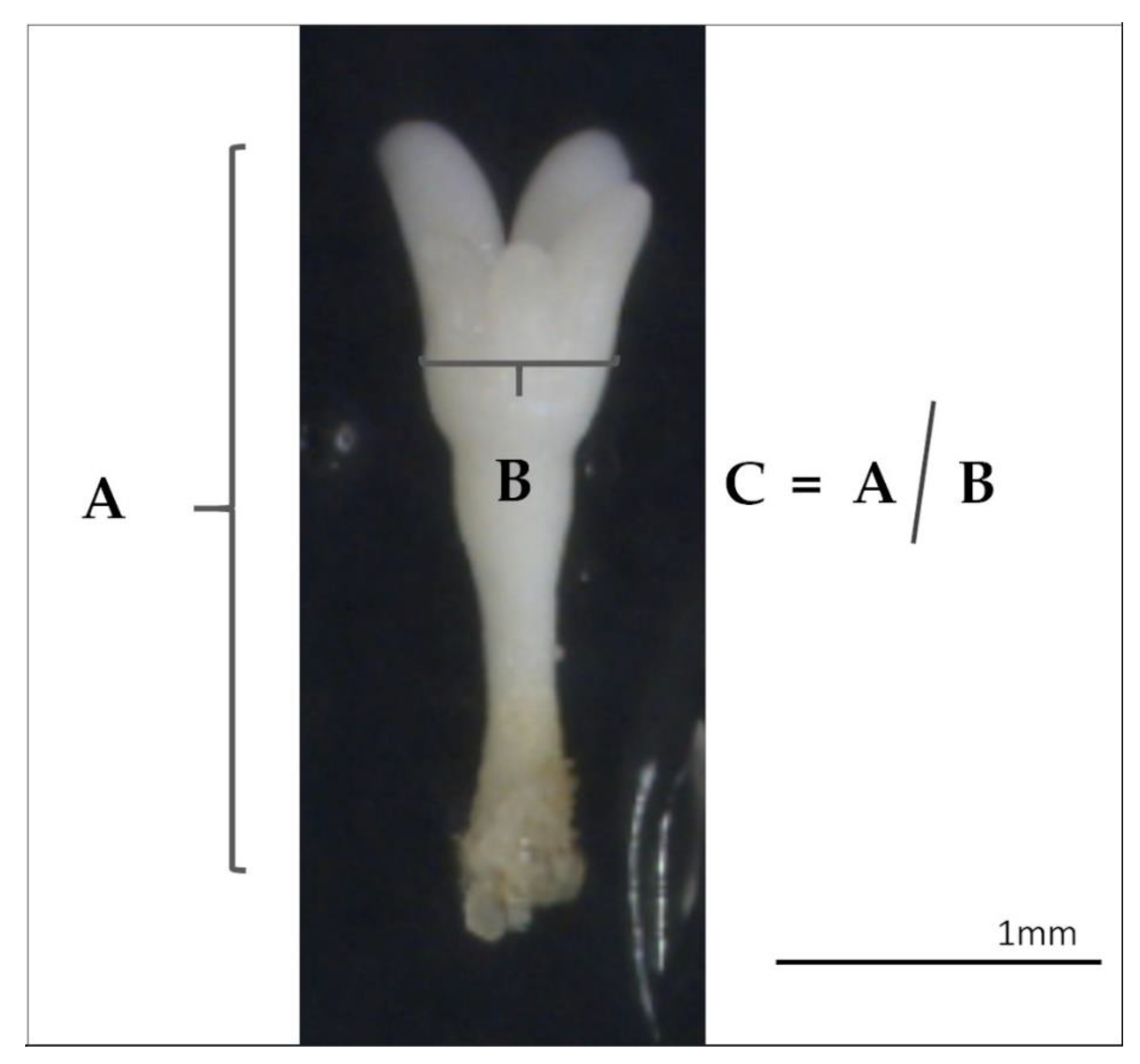
Regarding the morphological analysis of the SES obtained, more elongated embryos were obtained from samples induced at 60 °C for 5 min and from the control treatment, while the ones obtained from 40 °C for 4 h or 50 °C for 30 min were smaller and more barrel-shaped. Barrel-shaped form has been described as a symptom of poor quality embryos, which is usually accompanied by low germination rates in
P. taeda [
48],
P. Pinaster [
49], and
P. radiata [
43]. This is contrary to our results since no significant differences were found in the percentage of germinating somatic embryos, and, in fact, the highest percentage of germination was achieved in SES from ECLs initiated at 50 °C for 30 min. These results indicate that the shape of
P. halepensis SES may not be related with their ability to germinate as long as they are well formed.
4.2. Cytological Characterization of Embryogenic Cultures
Previous work on
Picea abies [
50] showed that embryogenic cultures appeared in three distinct levels or organization: PEM I, PEM II, and PEM III, formed by three different cell types (ECs, TLCs, and SCs) attached together in clusters in different proportions.
In Aleppo pine three different types of cells (ECs, TLCs, and SCs) in embryogenic cultures from all different temperature treatments (23, 40, 50, and 60 °C) were also found. Nonetheless, only two of the organizational structures described by [
50] could be found, since no PEM III were present. In
Picea abies [
50] PEM III were described as enlarged clumps of densely cytoplasmic cells loosely attached to each other that do not present polarity while in the case of
P. radiata these cells formed compact clusters which showed a well-organized structure with a clear polarization in samples from the control and a higher disorganization at samples from higher temperatures treatments [
43]. Also, despite the fact that well organized and polarized PEMs could be observed from different treatments, the common scenario featured high cellular disorganization with a disproportionate number of TLCs and SCs and no apparent discrepancies in the analyzed samples could be found between treatments when it comes to these features. In
P. sylvestris [
51] a continuous loop of embryo degeneration and differentiation of new embryos during the initiation of embryogenic tissue was suggested. However, they could not find if the differentiation pattern during the initiation stage would give rise to differences in the embryos obtained. This can explain the higher number of TLCs and SCs found in our samples and the fact that all of the analyzed embryogenic cultures produced somatic embryos despite their disorganization
A large number of starch was found both in light microscopy and TEM analysis from all treatments. In fact, TEM analysis showed that starch grains were present both in ECs and TLCs and in higher number in samples from the control. Some authors support the idea that, in ECs, the accumulation of starch around the nucleus is a signal of loss of embryogenic competence and it is also linked with the appearance of dead cells in meristematic centers, leading to low initiation and proliferation rates [
43,
52]. This was not the case in
P. halepensis, since no differences were found for initiation or proliferation rates between treatments. Other authors pointed out that starch, as a primary source of energy for cell proliferation, is abundantly found at competent embryogenic cells that will later differentiate into PEMs. High metabolic activity of these cells will deplete this reserve in such a way that it will no longer be present when embryos start to organize [
53,
54]. In
Picea glauca a precise pattern of storage compound accumulation during SE was identified with the first accumulation of starch further followed by the formation of lipid bodies and protein vacuoles [
55]. In
P. halepensis lipid bodies were also found in all treatments, although more abundantly in the control. Numerous protein bodies of different sizes or lipid bodies were also observed in ECs of other species [
54,
56]. In the case of
P. halepensis, the presence of the above mentioned storage compounds could not be directly correlated with the progress of SE and their function could probably be related with house-keeping functions of the cells more than with a control of the embryogenic process.
Finally, PCs were found in cultures induced at the higher temperatures. These compounds started to accumulate in the internal face of membrane vacuoles and were particularly notorious when cultures were initiated at 50 °C. The accumulation of this type of secondary metabolites has been described as playing an essential role preventing oxidative damage caused by different types of stresses [
27,
57], explaining their appearance in samples coming from high temperature treatments and not in control ones. In
P. radiata differences were also seen among different treatments respect to PCs with larger accumulations at higher temperatures [
43]. Also, opposite to our results, a positive association between accumulation of phenolic compounds and somatic embryo formation has been indicated in
Acca sellowiana [
58].
4.3. Quantification of Endogenous Cytokinins
A significantly higher concentration of isoprenoid CKs is present at EMs induced at 50 °C when compared to the control. Samples from 40 and 60 °C treatments presented concentrations that could be similar to one or another, but were generally intermediate. This pattern was detected through the analysis of the endogenous content of total CKs, CK ribosides, CK nucleotides,
cZ,
tZ and DHZ types, and finally for
cZ,
tZ, iP,
cZRMP,
tZRMP, DHZRMP, and DHZR. The results obtained in
P. halepensis are in line with the concept that short-term or mild stresses stimulate CKs accumulation, while prolonged or more severe stresses are generally associated with downregulation of active CKs levels [
59,
60,
61]. Furthermore, the ribosylated form of the initial precursor of CKs biosynthesis, iPA, across the time of heat stress exposure in
P. radiata showed an increase as other author pointed out [
62]. Contrary, when the same temperatures and induction times were analyzed in
P. radiata, the results showed a decreasing tendency when applying high temperatures for isoprenoid CKs, especially for those applied for the longest periods of time (40 and 50 °C) [
63].
Although the physiological function of each individual CKs is not yet completely understood they can be classified into different functional groups: (1) Active forms (bases), (2) translocation forms (ribosides), (3) precursors (nucleotides), and (4) storage and inactivated forms usually bound to glycosides (N-glucosides (7-G/9-G) and
O-glucosides) [
22,
64]. Regarding the analysis of different isoprenoid functional groups, the results showed that the highest concentration was obtained for CK nucleotides. As reviewed by [
19], it has been demonstrated that exogenously applied free cytokinin bases are rapidly metabolized into the corresponding ribosides and nucleotides and that, in general, high concentrations of these are found in developing organs. In this sense, when the same induction conditions were studied in
P. radiata EMs a higher concentration of CKs bases was found [
63], while at mature SES of the same species CKs ribosides showed the highest concentration in general [
24].
The majority of N-glucosides (7-G/9-G) and
O-glucosides, both for isoprenoids and aromatic CKs, were below the limit of detection. In terms of physiological activity, they exhibit little or no activity in CK bioassays [
65]. These data are in agreement with those reported for
P. radiata when high temperatures were analyzed [
63], and opposite to those obtained when different water availability and temperatures were tested at maturation stage of
P. radiata SE, despite the low values obtained [
24], or when different CKs were examined on the induction of organogenesis [
66]. The latter authors hypothesized that this type of forms are normally used by the plant as a form of detoxification in the face of excess hormone. Following this hypothesis, we can say that, in our case, the endogenous content of the hormone is not such that it needs this detoxification mechanism. Data obtained with Norway spruce showed that CKs
O-glucosides are the most abundant group during all stages of the SE process, including at callus stage, whereas CKs N-glucosides were practically absent [
67]. This can be explained by the difference in the tissues and species analyzed, and the fact that these metabolites have distinctive importance at different stages of SE in these species.
When endogenous concentrations of aromatic CKs were analyzed, a different pattern was obtained when compared to isoprenoid CKs. In K types, despite the fact that samples from 50 °C showed the higher concentrations, 40 °C showed a significantly lower concentration and, in BA types, samples from 40 °C showed significantly lower concentrations when compared with samples from the control. It seems that, when it comes to BA and K types, 40 °C for 4 h acted more as a severe stress, contrary to its effect on isoprenoids CKs [
59,
60,
61]. Also, our results show that the different temperature treatments did not lead to a clear accumulation of these hormones. Reports in apple trees [
68], where concentration of Kn tended to reduce with the intensity of water deficit, and in soybean [
69], where its growth under salt stress was significantly promoted by elevated Kn levels that effectively improved the quantities of isoflavones presented a more clear function of Kn in response to stress.
Kinetin showed to be the most abundant cytokinin in our samples which might be related to its exogenous application in the culture media. Despite the fact that the majority of naturally occurring CKs in plants are the N
6-isopentenyl conjugated adenine derivatives, in addition to a small amount of N
6-aromatic CK species [
64,
70], exogenous addition of CKs to culture media is known to have an effect by altering and increasing the endogenous CKs pool [
66,
71]. Similarly, reports in
Solanum tuberosum [
72],
Pinus pinea [
73], and
Pinus radiata [
66] found that aromatic CKs accounted for more than 90% of the total endogenous CK pool although isoprenoid forms are generally the most abundant CKs in plant materials that have not been hormonally treated.
High temperatures application overlap with higher endogenous concentrations of
oT,
cZ types and higher ratios between
cZ types and
tZ types. Within the CKs identified in response to mistletoe infestation in
P. sylvestris, the levels of biologically highly active (
tZ, iP) and less active (
cZ, DHZ) CKs were also found to be generally enhanced [
74]. It should be noted that, despite of the fact that there were no significant differences, the ratio between iP types and Z types also decreased as the temperatures applied rose. Z-type cytokinins are derived from iP-type compounds and not vice versa [
22,
60,
64] and both cell fate and organ formation have been associated with local concentration gradients of these hormones [
75]. Despite the fact that
oT is present in inferior concentrations comparing to
mT (
Figure 8;
Table 7) it seems to have a stronger effect in stress response as well as in cellular activity since
mT did not presented significant variations. Contrary, the results found in
Pinus pinea [
73] suggest that a higher cellular activity that lead to a stronger callogenesis response was related to higher concentrations of
mT as well as BAR and KR that also did not presented significant differences in our case.
Apart from their influence on stress response
per se, differences obtained in CKs profiles can explain some of the behaviors seen throughout different SE stages between the different treatments applied. After all, CKs are known to be responsible for the regulation of a number of aspects of plant growth and development, such as cell differentiation and growth [
25].
Although there were also no statistically significant differences at proliferation and germination stages, high concentrations of
tZ types,
tZRMP and iPR of samples from 50 and 60 °C treatments overlays with the highest germination rate achieved during SE, at the same time that the treatments with lower concentrations of these hormones presented lower proliferation rates. In coffee it was reported that the transition to the callus stage was accompanied by significant decrease of Z-type (
cis- and
trans-) [
76]. Experiments carried out in
P. pinea [
77] indicated that
tZR,
tZ, and DHZ had a relevant role in the callogenic process. Also, in
P. radiata [
62] ZR seems to be a key element to further acclimation and the recovery mechanism in plants while iPR and DHZR seems to be highly implied, across time, at the heat stress response.
It must be stressed out that samples from 60 °C and control treatment (23 °C) had a similar endogenous low hormonal concentration of
cZ, iP and DHZR, and these are the treatments that were able to induce a significantly higher number of somatic embryos. It was observed in [
78] that high concentrations of
Z and active CKs in general are important during the initial cell division phase of somatic embryogenesis, but not for the later stages of embryo development and maturation. However, as reported in
P. radiata, low levels of iP types leads to higher success in the SE process [
24,
63]. Additionally, the only CK N-glucoside found in our samples, K9G, presented a significantly higher concentration at 60 °C, the treatment that produced more somatic embryos. In
Cocos nucifera [
79] the isoprenoid cytokinin profiles showed a predominant pattern of 9-conjugation as a major metabolism route and the detection of high levels of inactive metabolic products has been cited as evidence of high CK turnover.



 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}