Changes in Multi-Level Biodiversity and Soil Features in a Burned Beech Forest in the Southern Italian Coastal Mountain

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
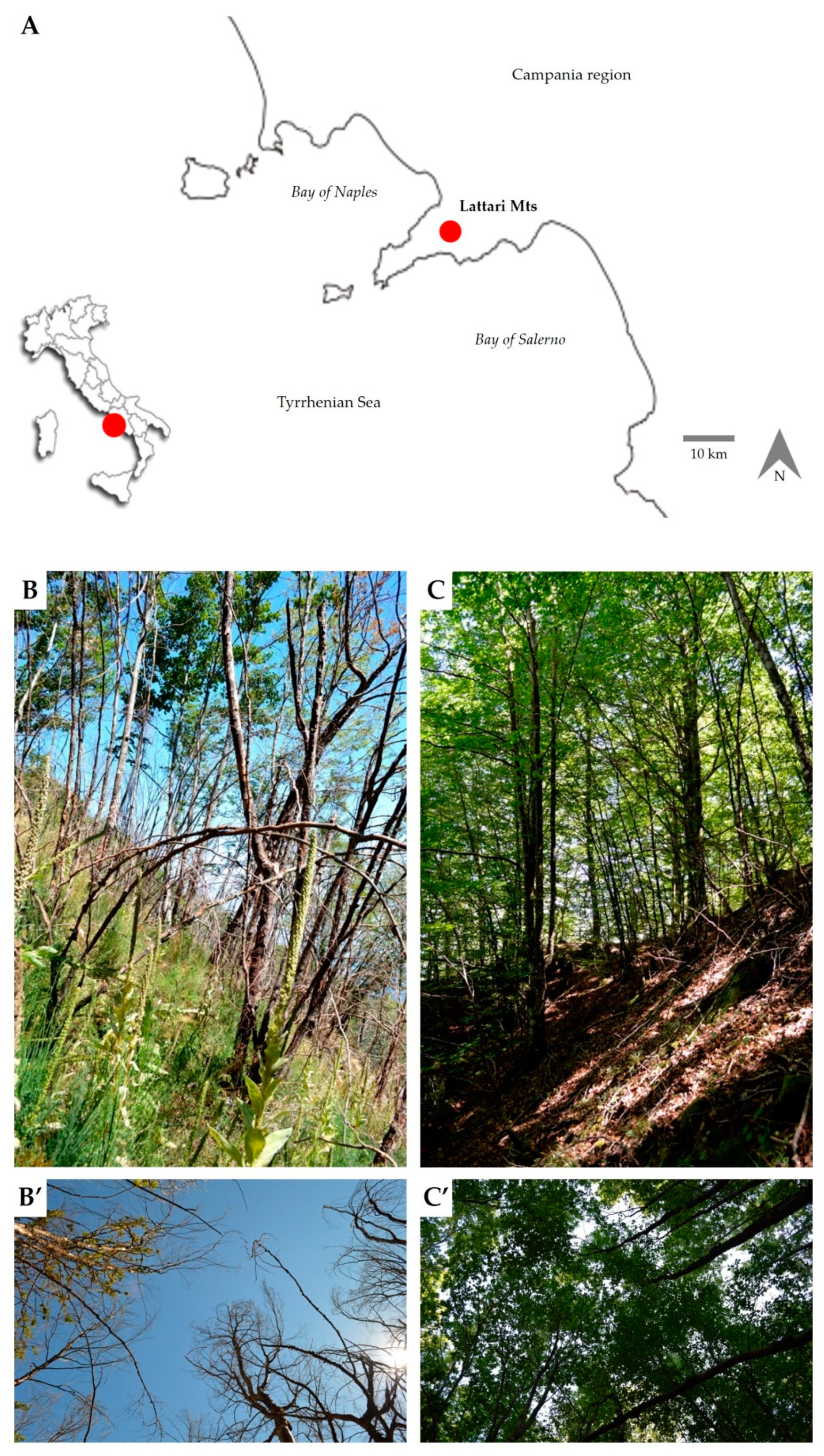
2.1. Study Area and Wildfire Event
2.2. Field Work and Laboratory Measurements
2.2.1. Sampling Design
2.2.2. Vegetation Sampling and Analyses
2.2.3. Soil Sampling and Analyses
2.2.4. Metagenomic Analysis
2.3. Bioinformatics and Statistics Analysis
3. Results
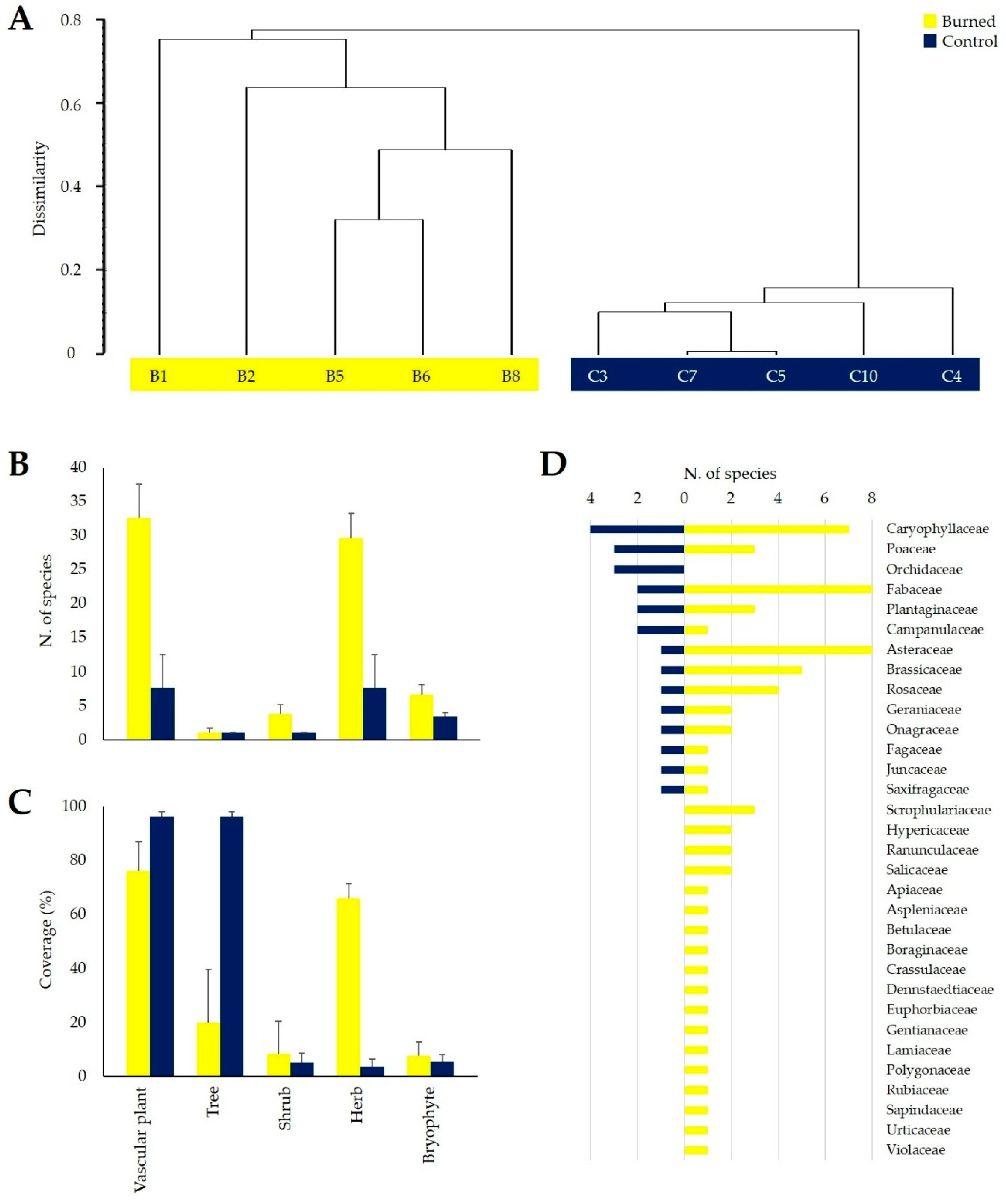
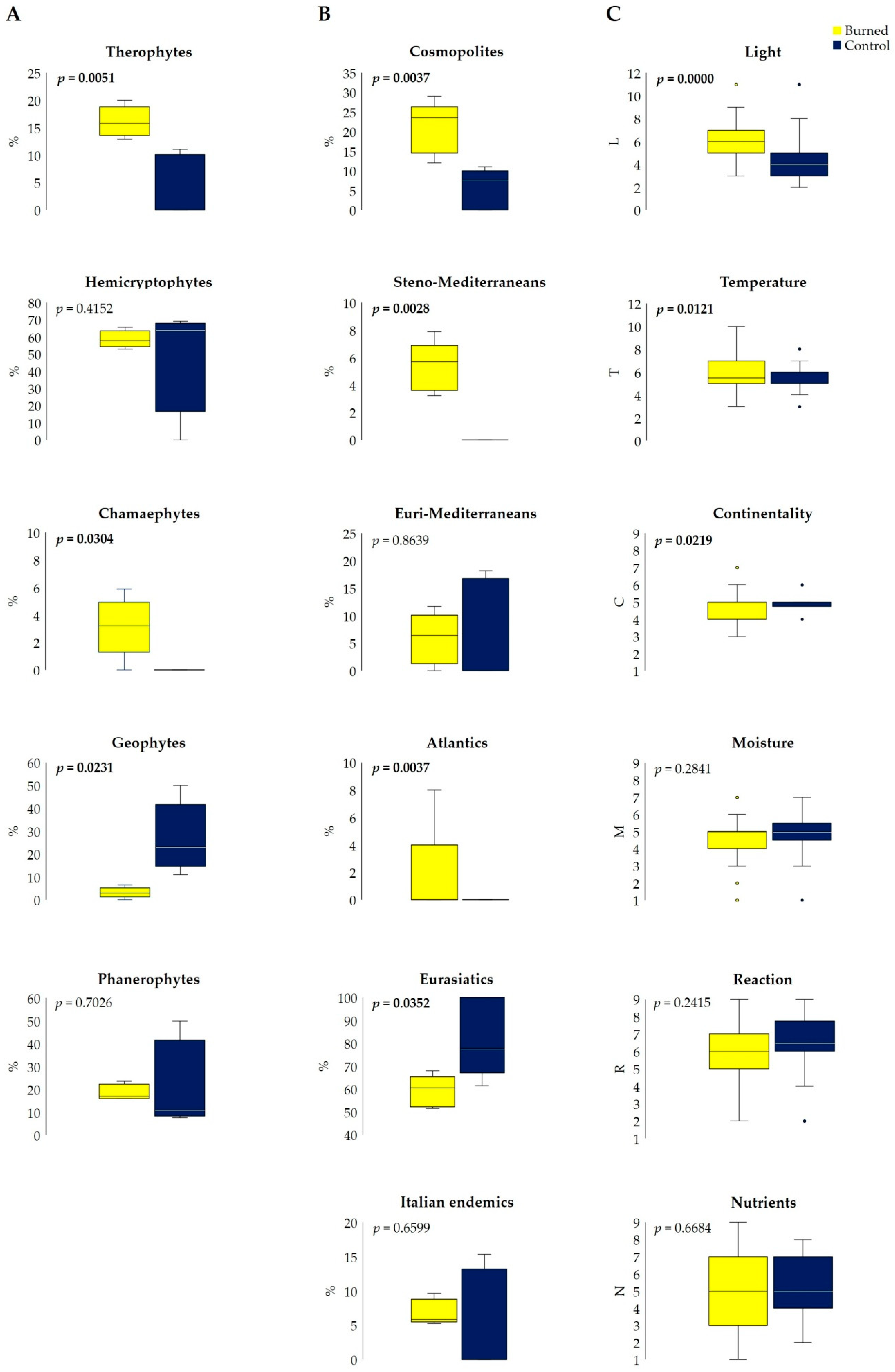
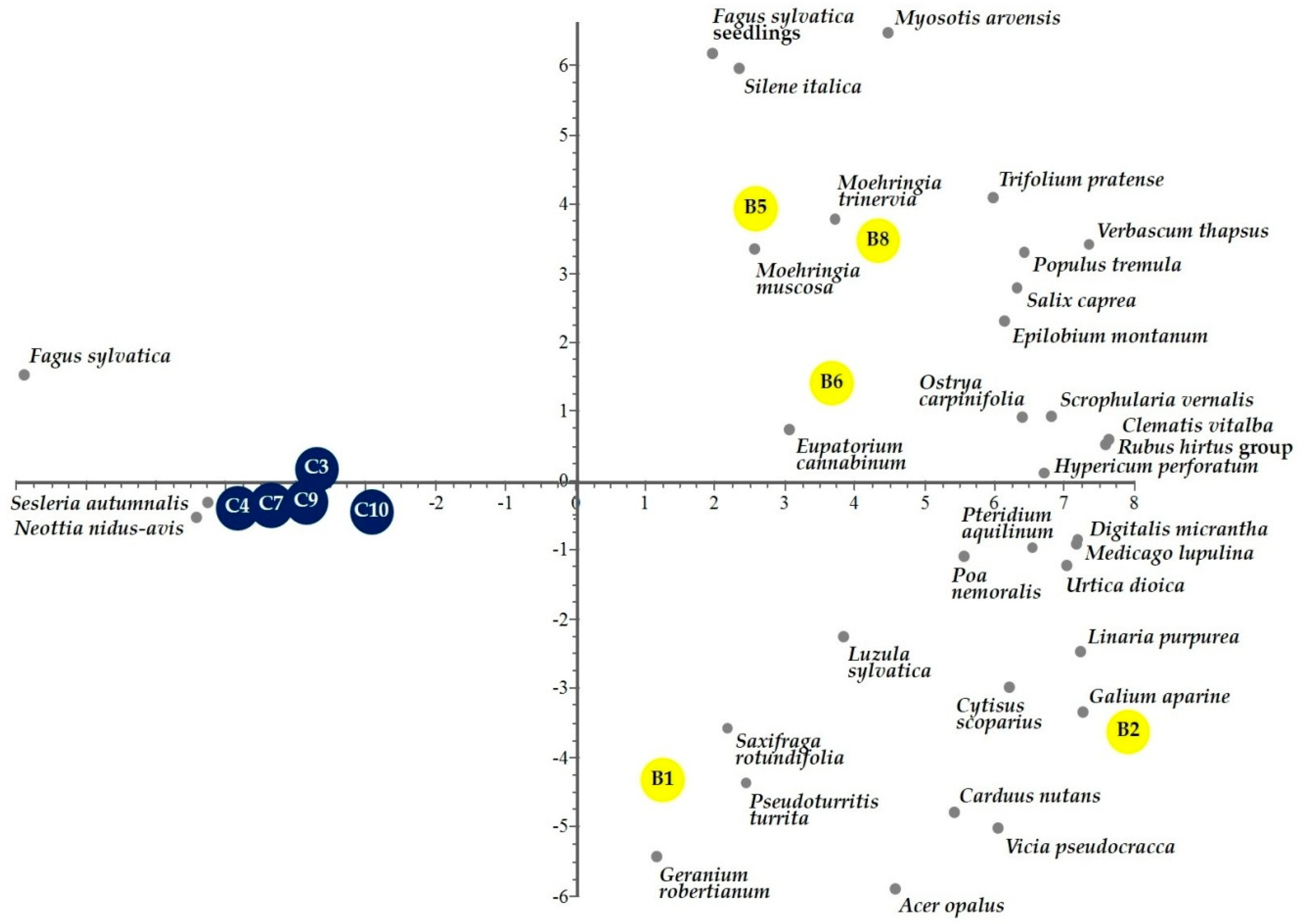
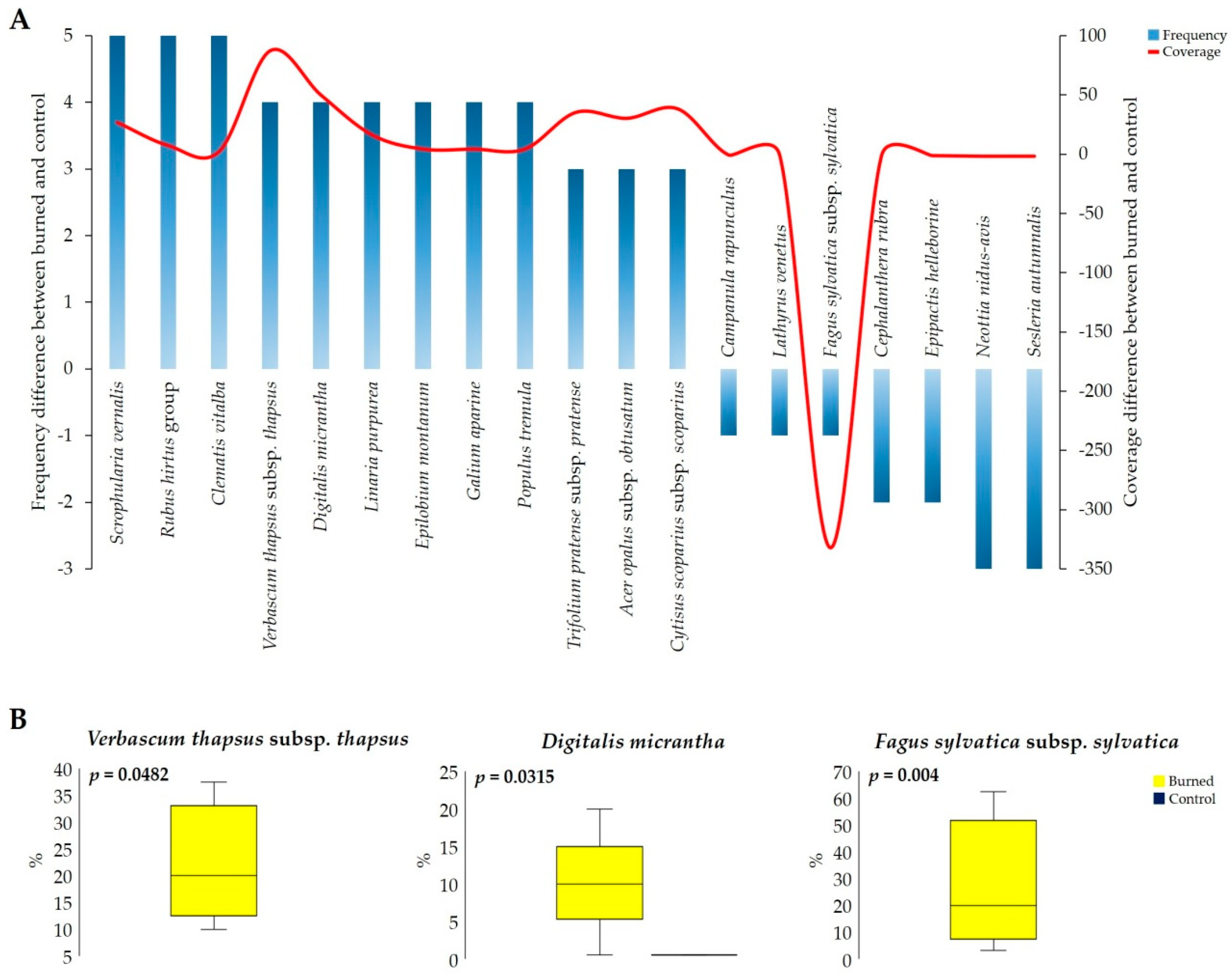
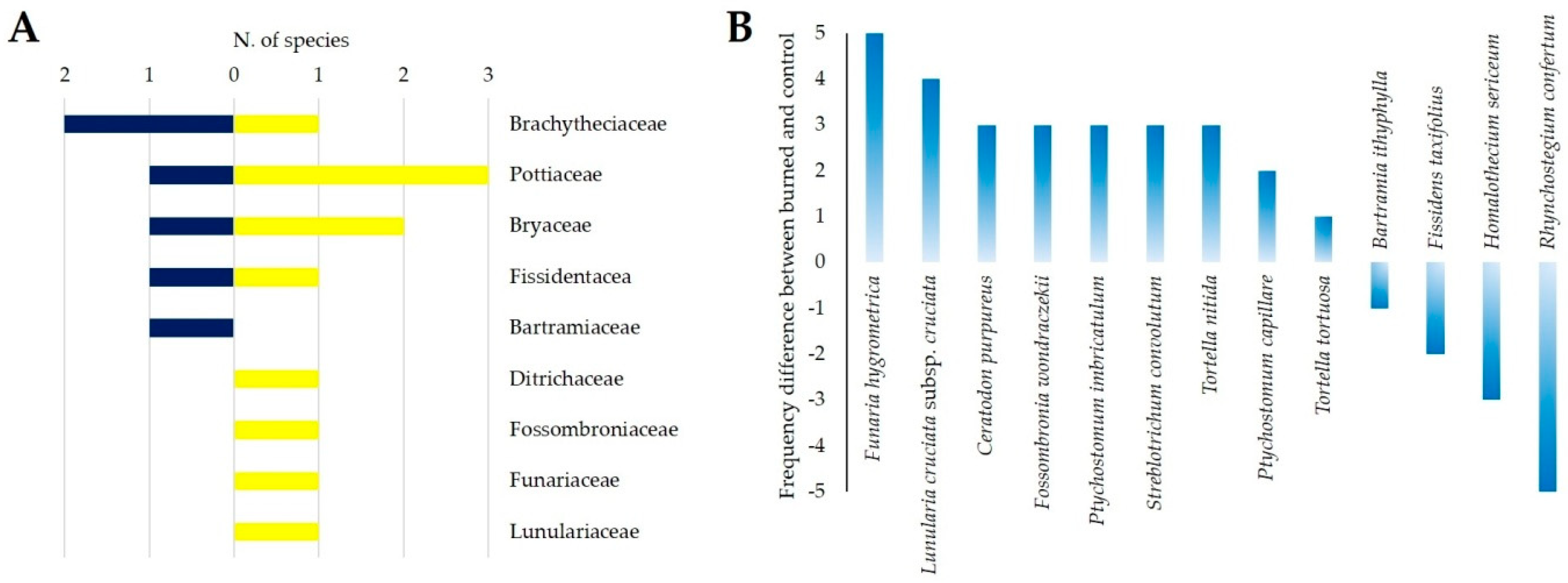
3.1. Fire Effects on Vascular Plants and Bryophyte Diversity
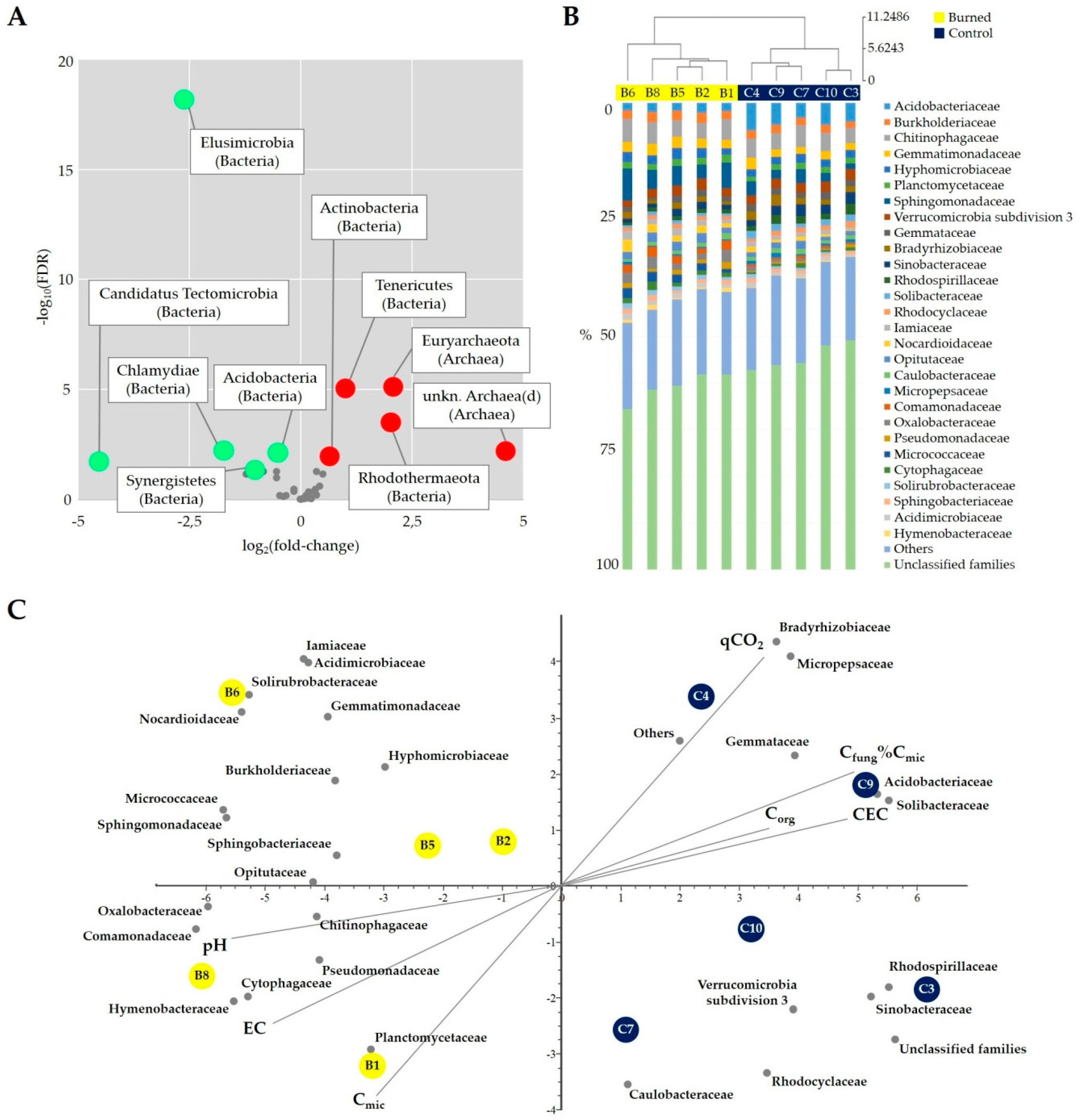
3.2. Fire Effect on Soil Chemical and Microbial Properties
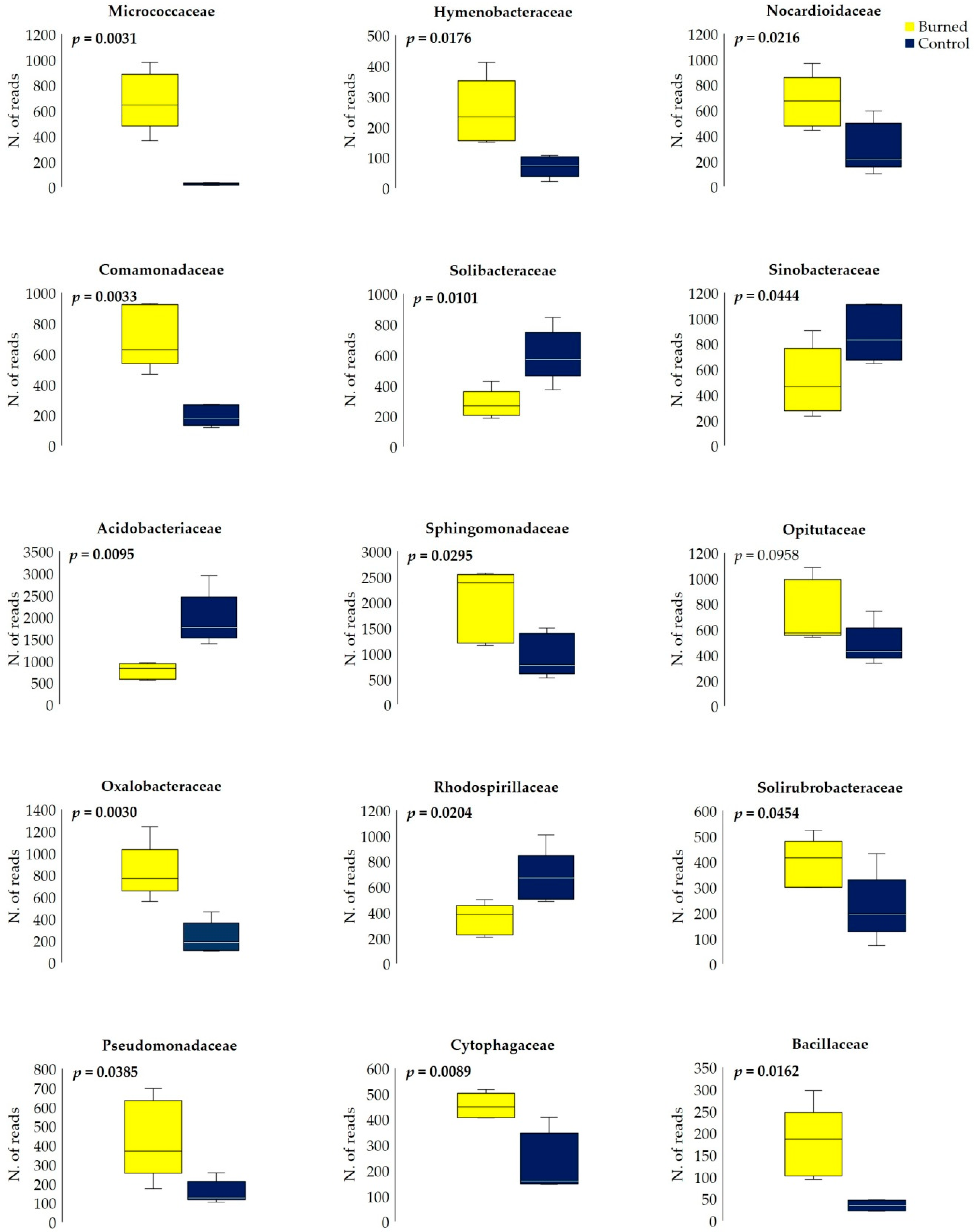
3.3. Fire Effect on Soil Bacterial Diversity
4. Discussion
4.1. Fire Effect on Vascular Plants and Bryophyte Diversity
4.2. Fire Effect on Soil Chemical and Microbial Properties
4.3. Fire Effect on Bacterial Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Van Mantgem, P.J.; Nesmith, J.C.B.; Keifer, M.; Knapp, E.E.; Flint, A.; Flint, L. Climatic stress increases forest fire severity across the western United States. Ecol. Lett. 2013, 16, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Diffenbaugh, N.S.; Swain, D.L.; Touma, D. Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA 2015, 112, 3931–3936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, S.; Reineking, B.; Sibold, J.; Bugmann, H. Modeling the impact of climate and vegetation on fire regimes in mountain landscapes. Landsc. Ecol. 2006, 21, 539–554. [Google Scholar] [CrossRef] [Green Version]
- Flannigan, M.D.; Stocks, B.J.; Wotton, B.M. Climate change and forest fires. Sci. Total Environ. 2000, 262, 221–229. [Google Scholar] [CrossRef]
- San-Miguel-Ayanz, J.; Durrant, T.; Boca, R.; Libertà, G.; Branco, A.; de Rigo, D.; Ferrari, D.; Maianti, P.; Artés Vivancos, T.; Costa, H.; et al. Forest Fires in Europe, Middle East and North Africa 2017; European Commission, JRC Technical Reports; Join Research Center: Ispra (Varese), Italy, 2018; p. 139. [Google Scholar]
- Ganteaume, A.; Camia, A.; Jappiot, M.; San-Miguel-Ayanz, J.; Long-Fournel, M.; Lampin, C. A Review of the Main Driving Factors of Forest Fire Ignition over Europe. Environ. Manag. 2013, 51, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Broncano, M.J.; Retana, J. Topography and forest composition affecting the variability in fire severity and post-fire regeneration occurring after a large fire in the Mediterranean basin. Int. J. Wildland Fire 2004, 13, 209–216. [Google Scholar] [CrossRef]
- Birch, D.S.; Morgan, P.; Kolden, C.A.; Abatzoglou, J.T.; Dillon, G.K.; Hudak, A.T.; Smith, A.M.S. Vegetation, topography and daily weather influenced burn severity in central Idaho and western Montana forests. Ecosphere 2015, 6, 1–23. [Google Scholar] [CrossRef]
- Rogers, B.M.; Soja, A.J.; Goulden, M.L.; Randerson, J.T. Influence of tree species on continental differences in boreal fires and climate feedbacks. Nat. Geosci. 2015, 8, 228–234. [Google Scholar] [CrossRef]
- Fares, S.; Bajocco, S.; Salvati, L.; Camarretta, N.; Dupuv, J.L.; Xanthopoulos, G.; Guijarro, M.; Madrigal, J.; Hernando, C.; Corona, P. Characterizing potential wildland fire fuel in live vegetation in the Mediterranean region. Ann. For. Sci. 2017, 74, 1. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Yang, J.; White, M.; Liu, Z. Predicting Potential Fire Severity Using Vegetation, Topography and Surface Moisture Availability in a Eurasian Boreal Forest Landscape. Forests 2018, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G. Simulating Mediterranean landscape pattern and vegetation dynamics under different fire regimes. Plant Ecol. 2006, 187, 249–259. [Google Scholar] [CrossRef]
- He, T.; Lamont, B.B.; Pausas, J.G. Fire as a key driver of Earth’s biodiversity. Biol. Rev. 2019, 94, 1983–2010. [Google Scholar] [CrossRef]
- Ducrey, M.; Duhoux, F.; Huc, R.; Rigolot, E. The ecophysiological and growth responses of Aleppo pine (Pinus halepensis) to controlled heating applied to the base of the trunk. Can. J. For. Res. 1996, 26, 1366–1374. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.; Lin, X.; Zhang, H.; Zeng, J.; Chu, H. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Ramesh, A.; Sharma, M.P.; Joshi, O.P.; Govaerts, B.; Steenwerth, K.L.; Karlen, D.L. Microbial community structure and diversity as indicators for evaluating soil quality. In Biodiversity, Biofuels, Agroforestry and Conservation Agriculture; Lichtfouse, E., Ed.; Sustainable Agriculture Reviews; Springer: Amsterdam, The Netherlands, 2010; Volume 5, pp. 317–358. [Google Scholar]
- Dominati, E.; Patterson, M.; Mackay, A. A framework for classifying and quantifying the natural capital and ecosystem services of soils. Ecol. Econ. 2010, 69, 1858–1868. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Matei, G.M.; Matei, S.; Mocanu, V.; Dumitru, S. Microbiological characterization of suppressive forest soil from Enisala. Ann. Univ. Craiova-Agric. Montanol. Cadastre Ser. 2016, 46, 341–347. [Google Scholar]
- Weller, D.M.; Raaijmakers, J.M.; McSpadden Gardener, B.B.; Thomashow, L.S. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Gu, J.; Li, Y.; Qian, X.; Sun, W.; Wang, X.; Gao, H.; Zhen, L.; Lei, Y. Analyses of microbial biomass and community diversity in kiwifruit orchard soils of different planting ages. Acta Ecol. Sin. 2015, 35, 22–28. [Google Scholar] [CrossRef]
- Giuditta, E.; Marzaioli, R.; Esposito, A.; Ascoli, D.; Stinca, A.; Mazzoleni, S.; Rutigliano, F.A. Soil microbial diversity, biomass, and activity in two pine plantations of Southern Italy treated with prescribed burning. Forests 2020, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Hamman, S.T.; Burke, I.C.; Stromberger, M.E. Relationships between microbial community structure and soil environmental conditions in a recently burned system. Soil Biol. Biochem. 2007, 39, 1703–1711. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; De Marco, A.; D’Ascoli, R.; Castaldi, S.; Gentile, A.; Virzo De Santo, A. Impact of fire on fungal abundance and microbial efficiency in C assimilation and mineralisation in a Mediterranean maquis soil. Biol. Fertil. Soils 2007, 44, 377–381. [Google Scholar] [CrossRef]
- Wang, Q.; Zhong, M.; Wang, S. A meta-analysis on the response of microbial biomass, dissolved organic matter, respiration, and N mineralization in mineral soil to fire in forest ecosystems. For. Ecol. Manag. 2012, 271, 91–97. [Google Scholar] [CrossRef]
- Pietikainen, J.; Fritze, H. Clear-cutting and prescribed burning in coniferous forest—Comparison of effects on soil fungal and total microbial biomass, respiration activity and nitrification. Soil Biol. Biochem. 1995, 27, 101–109. [Google Scholar] [CrossRef]
- Bárcenas-Moreno, G.; Bååth, E. Bacterial and fungal growth in soil heated at different temperatures to simulate a range of fire intensities. Soil Biol. Biochem. 2009, 41, 2517–2526. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire in Mediterranean climate ecosystems-A comparative overview. Isr. J. Ecol. Evol. 2012, 58, 123–135. [Google Scholar]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A.; et al. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef] [Green Version]
- Madrigal, J.; Souto-García, J.; Calama, R.; Guijarro, M.; Picos, J.; Hernando, C. Resistance of Pinus pinea L. bark to fire. Int. J. Wildland Fire 2019, 28, 342–353. [Google Scholar] [CrossRef]
- Van Gils, H.; Odoi, J.; Andrisano, T. From monospecific to mixed forest after fire? For. Ecol. Manag. 2010, 259, 433–439. [Google Scholar] [CrossRef]
- Walentowski, H.; Müller-Kroehling, S.; Bergmeier, E.; Bernhardt-Römermann, M.; Gossner, M.; Reif, A.; Schulze, E.; Bußler, H.; Strätz, C.; Adelmann, W. Faunal diversity of Fagus sylvatica forests: A regional and European perspective based on three indicator groups. Ann. For. Res. 2014, 57, 215–231. [Google Scholar]
- Wagner, S.; Collet, C.; Madsen, P.; Nakashizuka, T.; Nyland, R.D.; Sagheb-Talebi, K. Beech regeneration research: From ecological to silvicultural aspects. For. Ecol. Manag. 2010, 259, 2172–2182. [Google Scholar] [CrossRef]
- Bär, A.; Nardini, A.; Mayr, S. Post-fire effects in xylem hydraulics of Picea abies, Pinus sylvestris and Fagus sylvatica. New Phytol. 2018, 217, 1484–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packham, J.R.; Thomas, P.; Atkinson, M.; Degen, T. Biological flora of the British Isles: Fagus sylvatica. J. Ecol. 2012, 100, 1557–1608. [Google Scholar] [CrossRef]
- Maringer, J.; Wohlgemuth, T.; Neff, C.; Pezzatti, G.; Conedera, M. Postfire spread of alien plant species in a mixed broad-leaved forest of the Insubric region. Flora 2012, 1, 19–29. [Google Scholar] [CrossRef]
- Maringer, J.; Conedera, M.; Ascoli, D.; Schmatz, D.R.; Wohlgemuth, T. Resilience of European beech forests (Fagus sylvatica L.) after fire in a global change context. Int. J. Wildland Fire 2016, 25, 699–710. [Google Scholar] [CrossRef]
- Ascoli, D.; Castagneri, D.; Valsecchi, C.; Conedera, M.; Bovio, G. Postfire restoration of beech stands in the Southern Alps by natural regeneration. Ecol. Eng. 2013, 54, 210–217. [Google Scholar] [CrossRef]
- Ascoli, D.; Vacchiano, G.; Maringer, J.; Bovio, G.; Conedera, M. The synchronicity of masting and intermediate severity fire effects favor beech recruitment. For. Ecol. Manag. 2015, 353, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Gimeno-Garcia, E.; Andreu, V.; Rubio, J.L. Influence of vegetation recovery on water erosion at short and medium-term after experimental fires in a Mediterranean shrubland. Catena 2007, 69, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Martínez, S.; Penas, Á.; Díaz, T.E. Bioclimatic & Biogeographic Maps of Europe. 2004. Available online: http://www.globalbioclimatics.org/form/maps.htm (accessed on 10 June 2020).
- Blasi, C.; Marignani, M.; Copiz, R.; Fipaldini, M.; Del Vico, E. (Eds.) Le Aree Importanti per le Piante Nelle Regioni d’Italia: Il Presente e il Futuro Della Conservazione del Nostro Patrimonio Botanico; Progetto Artiser: Roma, Italy, 2010; p. 224. [Google Scholar]
- CARG. Carta Geologica d’Italia. Scala 1:50.000. Foglio 466–485: Sorrento-Termini. Note Illustrative alla Carta Geologica d’Italia alla Scala 1:50.000. Fogli 466–485 Sorrento-Termini.ISPRA, CNR. 2014. Available online: https://www.isprambiente.gov.it/Media/carg/campania.html (accessed on 10 June 2020).
- Di Gennaro, A. I Sistemi di Terre della Campania; Risorsa: Firenze, Italy, 2002; p. 64. [Google Scholar]
- Zitti, S.; Frattaroli, A.R.; Carli, E.; Cutini, M. 9210 *Faggeti degli Appennini con Taxus e Ilex. In Manuali per il Monitoraggio di Specie e Habitat di Interesse Comunitario (Direttiva 92/43/CEE) in Italia: Habitat; Angelini, P., Casella, L., Grignetti, A., Genovesi, P., Eds.; Serie Manuali e Linee Guida 142/2016; ISPRA: Roma, Italy, 2016; pp. 234–235. [Google Scholar]
- Cancellieri, L.; Caneva, G.; Cutini, M. Phytosociology and ecology of the Mediterranean forests ecosystems in the Amalfi Coast (Monti Lattari, Italy). Rend. Lincei 2017, 28, 651–671. [Google Scholar] [CrossRef]
- Di Pietro, R.; Izco, J.; Blasi, C. Contribute to the nomenclatural knowledge of the beech-woodland syntaxa of southern Italy. Plant Biosyst. 2004, 138, 27–36. [Google Scholar] [CrossRef]
- Peruzzi, L.; Domina, G.; Bartolucci, F.; Galasso, G.; Peccenini, S.; Raimondo, F.M.; Albano, A.; Alessandrini, A.; Banfi, E.; Barberis, G.; et al. An inventory of the names of vascular plants endemic to Italy, their loci classici and types. Phytotaxa 2015, 196, 1–217. [Google Scholar] [CrossRef] [Green Version]
- Stinca, A. Lonicera stabiana Guss. & Pasq. In La Flora Endemica Minacciata delle Montagne Italiane; Conti, F., Bartolucci, F., Di Martino, L., Manzi, A., Eds.; Club Alpino Italiano: Milano, Italy, 2019; pp. 340–342. [Google Scholar]
- Orsenigo, S.; Montagnani, C.; Fenu, G.; Gargano, D.; Peruzzi, L.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Bovio, M.; et al. Red Listing plants under full national responsibility: Extinction risk and threats in the vascular flora endemic to Italy. Biol. Conserv. 2018, 224, 213–222. [Google Scholar] [CrossRef]
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M.; et al. Red list of threatened vascular plant species in Italy. Plant Biosyst. 2020. [Google Scholar] [CrossRef]
- Battipaglia, G.; Tognetti, R.; Valese, E.; Ascoli, D.; De Luca, P.F.; Basile, S.; Ottaviano, M.; Mazzoleni, S.; Marchetti, M.; Esposito, A. Incendi 2017: Un’importante lezione. For. J. Silvic. For. Ecol. 2017, 14, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria; New York, NY, USA, 1964; p. 865. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, Volume 2; Cambridge University Press: Cambridge, UK, 1968; p. 465. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, Volume 3; Cambridge University Press: Cambridge, UK, 1972; p. 381. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, Volume 4; Cambridge University Press: Cambridge, UK, 1976; p. 515. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, Volume 5; Cambridge University Press: Cambridge, UK, 1980; p. 463. [Google Scholar]
- Tutin, T.G.; Burges, N.A.; Chater, A.O.; Edmondson, J.R.; Heywood, V.H.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. (Eds.) Flora Europaea, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993; Volume 1, p. 581. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, Volume 1; Edagricole: Bologna, Italy, 2017; p. 1064. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, Volume 2; Edagricole: Bologna, Italy, 2017; p. 1178. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, Volume 3; Edagricole: Bologna, Italy, 2018; p. 1288. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, Volume 4; Edagricole: Bologna, Italy, 2019; p. 1054. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Smith, A.J.E. The Liverworts of Britain and Ireland; Cambridge University Press: Cambridge, UK, 1991; p. 378. [Google Scholar]
- Smith, A.J.E. The Moss Flora of Britain and Ireland; Cambridge University Press: Cambridge, UK, 2010; p. 1026. [Google Scholar]
- Cortini Pedrotti, C. Flora dei Muschi d’Italia. Sphagnopsida. Andreaeopsida. Bryopsida (I Parte); Antonio Delfino Editore: Roma, Italy, 2001; p. 817. [Google Scholar]
- Cortini Pedrotti, C. Flora dei muschi d’Italia. Bryopsida (II Parte); Antonio Delfino Editore: Roma, Italy, 2006; pp. 818–1235. [Google Scholar]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Valori di indicazione delle piante vascolari della Flora d’Italia. Braun-Blanquetia 2005, 9, 1–97. [Google Scholar]
- Vacchiano, G.; Lonati, M.; Berretti, R.; Motta, R. Drivers of Pinus sylvestris L. regeneration following small, high-severity fire in a dry, inner-alpine valley. Plant Biosyst. 2015, 149, 354–363. [Google Scholar] [CrossRef]
- Khapugin, A.A.; Vargot, E.V.; Chugunov, G.G. Vegetation recovery in fire damaged forests: A case study at the southern boundary of the taiga zone. For. Stud. 2016, 64, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, V.; Carranza, M.L.; Evangelista, A.; Marchetti, M.; Stinca, A.; Stanisci, A. Long term changes in the composition, ecology and structure of Pinus mugo scrubs in the Apennines (Italy). Diversity 2018, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Mei, G.; Pesaresi, S.; Corti, G.; Cocco, S.; Colpi, C.; Taffetani, F. Changes in vascular plant species composition, top-soil and seed-bank along coppice rotation in an Ostrya carpinifolia forest. Plant Biosyst. 2020, 154, 259–268. [Google Scholar] [CrossRef]
- USDA-NRCS. Soil Survey Laboratory Methods Manual; Soil Survey Investigations Report No. 42, Version 4.0; Burc, R., Ed.; National Soil Survey Center: Lincoln, NE, USA, 2004; p. 487.
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publications: Oxford, UK, 1989; p. 368. [Google Scholar]
- Springer, U.; Klee, J. Prüfung der Leistungsfähigkeit von einigen wichtigeren Verfahren zur Bestimmung des Kohlemstoffs mittels Chromschwefelsäure sowie Vorschlag einer neuen Schnellmethode. Z. Pflanz. Düngung Bodenkd. 1954, 64, 1–26. [Google Scholar] [CrossRef]
- Sleutel, S.; De Neve, S.; Singier, B.; Hofman, G. Quantification of Organic Carbon in Soils: A Comparison of Methodologies and Assessment of the Carbon Content of Organic Matter. Commun. Soil Sci. Plant Anal. 2007, 38, 2647–2657. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Sundman, V.; Sivelä, S. A comment on the membrane filter technique for estimation of length of fungal hyphae in soil. Soil Biol. Biochem. 1978, 10, 399–401. [Google Scholar] [CrossRef]
- Olson, F.C.W. Quantitative estimates of filamentous algae. Trans. Am. Microsc. Soc. 1959, 69, 272–279. [Google Scholar] [CrossRef]
- Berg, B.; Söderström, B. Fungal biomass and nitrogen in decomposing Scots pine needle litter. Soil Biol. Biochem. 1979, 11, 339–341. [Google Scholar] [CrossRef]
- ISO 16072. Soil Quality—Laboratory Methods for Determination of Microbial Soil Respiration; International Organization for Standardization: Geneva, Switzerland, 2002. [Google Scholar]
- Pell, M.; Stenström, J.; Granhall, U. Monitoring and evaluating soil quality. In Microbiological Methods for Assessing Soil Quality; Bloem, J., Benedetti, A., Hopkins, D.W., Eds.; CAB International: Wallingford, UK, 2006; pp. 117–127. [Google Scholar]
- Riffaldi, R.; Saviozzi, A.; Levi-Minzi, R. Carbon mineralization kinetics as influenced by soil properties. Biol. Fertil. Soils 1996, 22, 293–298. [Google Scholar] [CrossRef]
- D’Ascoli, R.; Rutigliano, F.A.; De Pascale, R.A.; Gentile, A.; Virzo De Santo, A. Functional diversity of the microbial community in Mediterranean maquis soils as affected by fires. Int. J. Wildland Fire 2005, 14, 355–363. [Google Scholar] [CrossRef]
- Killham, K. Soil Ecology; Cambridge University Press: Cambridge, UK, 1994; p. 242. [Google Scholar]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Blackwell Scientific Publications: Oxford, UK, 1979; p. 372. [Google Scholar]
- Anderson, T.H.; Domsch, K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Canullo, R.; Starlinger, F.; Granke, O.; Fischer, R.; Aamlid, D. Part VI.1: Assessment of Ground Vegetation. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-Ordinating Centre, Ed.; Thunen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 15. Available online: http://icp-forests.net/page/icp-forests-manual (accessed on 15 June 2020).
- Knapp, E.E.; Ritchie, M.W. Response of understory vegetation to salvage logging following a high-severity wildfire. Ecosphere 2016, 7, e01550. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. ACM SIGMOBILE Mob. Comput. Commun. Rev. 1948, 27, 379–423. [Google Scholar]
- Sokal, R.R.; Rholf, F.J. Biometry: The Principles and Practices of Statistic in Biological Research, 3rd ed.; W.H. Freeman & Company: New York, NY, USA, 1995; p. 887. [Google Scholar]
- Podani, J. SYN-TAX 2000. Computer Programs for Data Analysis in Ecology and Systematics; User’s Manual; Scientia Publishing: Budapest, Hungary, 2001; p. 452. [Google Scholar]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Paytuví, A.; Battista, E.; Scippacercola, F.; Aiese Cigliano, R.; Sanseverino, W. GAIA: An integrated metagenomics suite. bioRxiv 2019. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Mcmurdie, P.J.; Holmes, S.; Watson, M. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purdie, R.W.; Slatyer, R.O. Vegetation succession after fire in sclerophyll woodland communities in south-eastern Australia. Aust. J. Ecol. 1976, 1, 223–236. [Google Scholar] [CrossRef]
- Trabaud, L. Dynamics after fire of sclerophyllous plant communities in the mediterranean basin. Ecol. Mediterr. 1987, 13, 25–37. [Google Scholar] [CrossRef]
- Guo, Q. Early post-fire succession in California chaparral: Changes in diversity, density, cover and biomass. Ecol. Res. 2001, 16, 471–485. [Google Scholar] [CrossRef]
- Harvey, B.J.; Holzman, B.A. Divergent successional pathways of stand development following fire in a Californian closed-cone pine forest. J. Veg. Sci. 2014, 25, 88–99. [Google Scholar] [CrossRef]
- Naveh, Z. The Role of Fire and Its Management in the Conservation of Mediterranean Ecosystems and Landscapes. In The Role of Fire in Mediterranean-Type Ecosystems; Moreno, J.M., Oechel, W.C., Eds.; Springer: New York, NY, USA, 1995; pp. 163–185. [Google Scholar]
- Capitanio, R.; Carcaillet, C. Post-fire Mediterranean vegetation dynamics and diversity: A discussion of succession models. For. Ecol. Manag. 2008, 255, 431–439. [Google Scholar] [CrossRef]
- Herranz, J.M.; Martinez-Sanchez, J.J.; De Las Heras, J.; Ferrandis, P. Stages of plant succession in Fagus sylvatica L. and Pinus sylvestris L. forest of Tejera Negra National Park (Central Spain), three years after fire. Isr. J. Plant Sci. 1996, 44, 347–358. [Google Scholar] [CrossRef]
- Christopoulou, A.; Kazanis, D.; Fyllas, N.M.; Arianoutsou, M. Post-fire recovery of Abies cephalonica forest communities: The case of Mt Parnitha National Park, Attica, Greece. iFor.-Biogeosci. For. 2018, 11, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Kazanis, D.; Arianoutsou, M. Vegetation composition in a post-fire successional gradient of Pinus halepensis forests in Attica, Greece. Int. J. Wildland Fire 1996, 6, 83–91. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Neeman, G. Post-fire regeneration of natural Pinus halepensis forests in the East Mediterranean Basin. In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Ne’Eman, G., Trabaud, L., Eds.; Backhuys: Leiden, The Netherlands, 2000; pp. 269–290. [Google Scholar]
- Ganatsas, P.; Zagas, T.; Tsakaldimi, M.; Tsitsoni, T. Post-fire regeneration dynamics in a Mediterranean type ecosystem in Sithonia, northern Greece: Ten years after the fire. In Proceedings of the 10th MEDECOS Conference, Rhodes, Greece, 25 April–1 May 2004; Arianoutsou, M., Papanastasis, V., Eds.; Millpress: Rotterdam, The Netherlands, 2004; pp. 1–9. [Google Scholar]
- Ocak, A.; Kurt, L.; Oz, M.; Tug, N. Floristic al and ecological studies on burned black pine (Pinus nigra Arn. subsp. pallasiana (Lamb) Holboe) forest area at Central Anatolia. Asian J. Plant Sci. 2007, 6, 892–905. [Google Scholar]
- Nagel, T.A.; Svoboda, M.; Diaci, J. Regeneration patterns after intermediate wind disturbance in an old-growth Fagus–Abies forest in southeastern Slovenia. For. Ecol. Manag. 2006, 226, 268–278. [Google Scholar] [CrossRef]
- Bonanomi, G.; Incerti, G.; Abd El-Gawad, A.M.; Sarker, T.C.; Stinca, A.; Motti, R.; Cesarano, G.; Teobaldelli, M.; Saulino, L.; Cona, F.; et al. Windstorm disturbance triggers multiple species invasion in an urban Mediterranean forest. iFor.-Biogeosci. For. 2018, 11, 64–71. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; Fierro, A.; De Pascale, R.A.; De Marco, A.; Virzo De Santo, A. Role of fire on soil organic matter turnover and microbial activity in a mediterranean burned area. Dev. Soil Sci. 2002, 28B, 205–215. [Google Scholar]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Auld, T.D.; O’Connell, M.A. Predicting patterns of post-fire germination in 35 eastern Australian Fabaceae. Aust. J. Ecol. 1991, 16, 53–70. [Google Scholar] [CrossRef]
- Herranz, J.M.; Ferrandis, P.; Martinez-Sanchez, J.J. Influence of heat on seed germination of seven Mediterranean Leguminosae species. Plant Ecol. 1998, 136, 95–103. [Google Scholar] [CrossRef]
- Baeza, M.J.; Vallejo, V.R. Ecological mechanisms involved in dormancy breakage in Ulex parviflorus seeds. Plant Ecol. 2006, 183, 191–205. [Google Scholar] [CrossRef]
- Bekdouche, F.; Sahnoune, M.; Krouchi, F.; Achour, S.; Guemati, N.; Derridj, A. The contribution of legumes to post-fire regeneration of Quercus suber and Pinus halepensis forests in northeastern Algeria. Rev. D’écol. 2011, 66, 29–42. [Google Scholar]
- Johnson, D.W.; Susfalk, R.B.; Caldwell, T.G.; Murphy, J.F.; Miller, W.W.; Walker, R.F. Fire effects on carbon and nitrogen budgets in forests. In Biogeochemical Investigations of Terrestrial, Freshwater, and Wetland Ecosystems across the Globe; Kelman Wieder, R., Novák, M., Vile, M.A., Eds.; Reprinted from Water, Air, & Soil Pollution: Focus 4; Springer: Dordrecht, The Netherlands, 2004; pp. 263–275. [Google Scholar]
- Goergen, E.M.; Chambers, J.C. Influence of a native legume on soil N and plant response following prescribed fire in sagebrush steppe. Int. J. Wildland Fire 2009, 18, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Pancer-Koteja, E.; Szwagrzyk, J.; Bodziarczyk, J. Small scale spatial pattern and size structure of Rubus hirtus in a canopy gap. J. Veg. Sci. 1998, 9, 755–762. [Google Scholar] [CrossRef]
- Gazda, A.; Szwagrzyk, J.; Nybom, H.; Werlemark, G. Morphological and genetical variability of Rubus hirtus (Waldst. & Kitt.) plants under partly open forest canopy. Pol. J. Ecol. 2007, 55, 49–55. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Elsevier, Academic Press: London, UK, 2008; p. 800. [Google Scholar]
- Zagory, D.; Parmeter, J.R. Fungitoxicity of smoke. Phytopathology 1984, 74, 1027–1031. [Google Scholar] [CrossRef]
- Lin, H.-L.; Chumpookam, J.; Shiesh, C.-C.; Chung, W.-H. Smoke-water controls Pythium damping-off in papaya seedling. HortScience 2012, 47, 1453–1456. [Google Scholar] [CrossRef] [Green Version]
- Stinca, A. Le Orchidee della Penisola Sorrentina e dei Monti Lattari; Regione Campania, Assessorato all’Ecologia e alla Tutela dell’Ambiente; Nicola Longobardi Editore: Castellammare di Stabia (Napoli), Italy, 2014; p. 127. [Google Scholar]
- Gehring, E.; Conedera, M.; Maringer, J.; Giadrossich, F.; Guastini, E.; Schwarz, M. Shallow landslide disposition in burnt European beech (Fagus sylvatica L.) forests. Sci. Rep. 2019, 9, 8638. [Google Scholar] [CrossRef] [PubMed]
- Catling, P.M.; Sinclair, A.; Cuddy, D. Plant community composition and relationship of disturbed and undisturbed alvar woodland. Can. Field Nat. 2002, 116, 571–579. [Google Scholar]
- Bataineh, A.L.; Oswald, B.P.; Bataineh, M.M.; Williams, H.M.; Coble, D.W. Changes in understory vegetation of a ponderosa pine forest in northern Arizona 30 years after a wildfire. For. Ecol. Manag. 2006, 235, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.E.; Omi, P.N. Seed supply of native and cultivated grasses in pine forests of the southwestern United States and the potential for vegetation recovery following wildfire. Plant Ecol. 2006, 183, 1–8. [Google Scholar] [CrossRef]
- Stark, K.E.; Arsenault, A.; Bradfield, G.E. Soil seed banks and plant community assembly following disturbance by fire and logging in interior Douglas-fir forests of south-central British Columbia. Can. J. Bot. 2006, 84, 1548–1560. [Google Scholar] [CrossRef]
- Gross, K.L.; Werner, P.A. Biology of Canadian Weeds. 28. Verbascum Thapsus L. and Verbascum Blattaria L. Can. J. Plant Sci. 1978, 58, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Mazzoleni, S.; Esposito, A. Vegetation regrowth after fire and cutting of Mediterranean macchia species. In Fire in Mediterranean Ecosystems; Trabaud, L., Prodod, R., Eds.; ECSC-EEC-EAEC: Brussels, Belgium; Luxembourg, 1993; pp. 87–99. [Google Scholar]
- Esposito, A.; Strumia, S.; Buonanno, M.; Castaldo-Cobianchi, R.; Mazzoleni, S. Analysis of bryophyte dynamics after fires of Pine woodlands and Mediterranean macchia, southern Italy. In Fire Management and Landscape Ecology; Trabaud, L., Ed.; International Association of Wildland Fire: Washington, DC, USA, 1998; pp. 77–87. [Google Scholar]
- Esposito, A.; Mazzoleni, S.; Strumia, S. Post-fire bryophyte dynamics in Mediterranean vegetation. J. Veg. Sci. 1999, 10, 261–268. [Google Scholar] [CrossRef]
- Hoffman, G.R. Ecological Studies of Funaria hygrometrica Hedw. In Eastern Washington and Northern Idaho. Ecol. Monogr. 1966, 36, 157–180. [Google Scholar] [CrossRef]
- Brasell, H.M.; Mattay, J.P. Colonization by Bryophytes of Burned Eucalyptus Forest in Tasmania, Australia: Changes in Biomass and Element Content. Bryologist 1984, 87, 302–307. [Google Scholar] [CrossRef]
- Smith, R.J.; Abella, S.R.; Stark, L.R. Post-fire recovery of desert bryophyte communities: Effects of fires and propagule soil banks. J. Veg. Sci. 2014, 25, 447–456. [Google Scholar] [CrossRef]
- Zeibig, A.; Diaci, J.; Wagner, S. Gap disturbance patterns of a Fagus sylvatica virgin forest remnant in the mountain vegetation belt of Slovenia. For. Snow Landsc. Res. 2005, 79, 69–80. [Google Scholar]
- Collet, C.; Piboule, A.; Leroy, O.; Frochot, H. Advance Fagus sylvatica and Acer pseudoplatanus seedlings dominate tree regeneration in a mixed broadleaved former coppice-with-standards forest. Forestry 2008, 81, 135–150. [Google Scholar] [CrossRef] [Green Version]
- Mazzoleni, S.; Bonanomi, G.; Incerti, G.; Chiusano, M.L.; Termolino, P.; Mingo, A.; Senatore, M.; Giannino, F.; Cartenì, F.; Rietkerk, M.; et al. Inhibitory and toxic effects of extracellular self-DNA in litter: A mechanism for negative plant–soil feedbacks? New Phytol. 2015, 205, 1195–1210. [Google Scholar] [CrossRef] [Green Version]
- Szwagrzyk, J.; Szewczyk, J.; Bodziarczyk, J. Dynamics of seedling banks in beech forest: Results of a 10-year study on germination, growth and survival. For. Ecol. Manag. 2001, 141, 237–250. [Google Scholar] [CrossRef]
- Alberti, G.; Boscutti, F.; Pirotti, F.; Bertacco, C.; De Simon, G.; Sigura, M.; Cazorzi, F.; Bonfanti, P. A LiDAR-based approach for a multi-purpose characterization of Alpine forests: An Italian case study. iFor.-Biogeosci. For. 2013, 6, 156–170. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire Management Impacts on Invasive Plants in the Western United States. Conserv. Biol. 2006, 20, 375–384. [Google Scholar] [CrossRef]
- Stinca, A.; Motti, R. Alien plant invasions in Astroni crater, a decades-long unmanaged forest in southern Italy. Atti della Soc. Toscana Sci. Nat. Mem. Ser. B 2017, 124, 101–108. [Google Scholar]
- Becker, T.; Dietz, H.; Billeter, R.; Buschmann, H.; Edwards, P.J. Altitudinal distribution of alien plant species in the Swiss Alps. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 173–183. [Google Scholar] [CrossRef]
- Alexander, J.M.; Kueffer, C.; Daehler, C.C.; Edwards, P.J.; Pauchard, A.; Seipel, T.; MIREN Consortium. Assembly of nonnative floras along elevational gradients explained by directional ecological filtering. Proc. Natl. Acad. Sci. USA 2011, 108, 656–661. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Fei, S.; Shen, Z.; Iannone, B.V.; Knott, J.; Chown, S.L. A global analysis of elevational distribution of non-native versus native plants. J. Biogeogr. 2018, 45, 793–803. [Google Scholar] [CrossRef]
- Aronson, M.F.; Handel, S.N.; La Puma, I.P.; Clemants, S.E. Urbanization promotes non-native woody species and diverse plant assemblages in the New York metropolitan region. Urban Ecosyst. 2015, 18, 31–45. [Google Scholar] [CrossRef]
- McDaniel, P.A.; Lowe, D.J.; Arnalds, O.; Ping, C.-L. Andisols. In Handbook of Soil Sciences, Vol. 1: Properties and Processes, 2nd ed.; Huang, P.M., Li, Y., Sumner, M.E., Eds.; CRC Press (Taylor & Francis): Boca Raton, FL, USA, 2012; pp. 33.29–33.48. [Google Scholar]
- Soil Science Division Staff. Soil Survey Manual; USDA, Agriculture Handbook No. 18; Ditzler, C., Scheffe, K., Monger, H.C., Eds.; Government Printing Office: Washington, DC, USA, 2017; p. 603.
- Certini, G.; Nocentini, C.; Knicker, H.; Arfaioli, P.; Rumpe, C. Wildfire effects on soil organic matter quantity and quality in two fire-prone Mediterranean pine forests. Geoderma 2011, 167–170, 148–155. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Shrestha, B.M. Stand age, fire and clearcutting affect soil organic carbon and aggregation of mineral soils in boreal forests. Soil Biol. Biochem. 2012, 50, 149–157. [Google Scholar] [CrossRef]
- Shen, J.P.; Chen, C.R.; Lewis, T. Long term repeated fire disturbance alters soil bacterial diversity but not the abundance in an Australian wet sclerophyll forest. Sci. Rep. 2016, 6, 19639. [Google Scholar] [CrossRef] [Green Version]
- Prendergast-Miller, M.T.; de Menezes, A.B.; Macdonald, L.M.; Toscas, P.; Bissett, A.; Baker, G.; Farrell, M.; Richardson, A.E.; Wark, T.; Thrall, P.H. Wildfire impact: Natural experiment reveals differential short-term changes in soil microbial communities. Soil Biol. Biochem. 2017, 109, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Knelman, J.E.; Schmidt, S.K.; Garayburu-Caruso, V.; Kumar, S.; Graham, E.B. Multiple, compounding disturbances in a forest ecosystem: Fire increases susceptibility of soil edaphic properties, bacterial community structure, and function to change with extreme precipitation event. Soil Syst. 2019, 3, 40. [Google Scholar] [CrossRef] [Green Version]
- Ulery, A.L.; Graham, R.C.; Amrhein, C. Wood-ash composition and soil pH following intense burning. Soil Sci. 1993, 156, 358–364. [Google Scholar] [CrossRef]
- Badía, D.; Martí, C. Effect of Simulated Fire on Organic Matter and Selected Microbiological Properties of Two Contrasting Soils. Arid Land Res. Manag. 2003, 17, 55–69. [Google Scholar] [CrossRef]
- Bogunović, I.; Kisić, I.; Jurišić, A. Influence of wildfire and fire suppression by seawater on soil properties. Appl. Ecol. Environ. Res. 2015, 13, 1157–1169. [Google Scholar] [CrossRef]
- Park, J.S.; Koo, K.-S.; Lee, E.J. The changes of soil salinity in the Pinus densiflora forest after seawater spread using a fire-fight helicopter. J. Ecol. Environ. 2015, 38, 443–450. [Google Scholar] [CrossRef]
- Santín, C.; Stefan, H.; Doerr, S.H.; Preston, C.M.; González-Rodríguez, G. Pyrogenic organic matter production from wildfires: A missing sink in the global carbon cycle. Glob. Chang. Biol. 2014, 21, 1621–1633. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.C.; DeLuca, T.H.; Newman, G.S.; MacKenzie, M.D.; Boyle, S.I. Post-fire vegetative dynamics as drivers of microbial community structure and function in forest soils. For. Ecol. Manag. 2005, 220, 166–184. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; Migliorini, M.; Maggi, O.; D’Ascoli, R.; Fanciulli, P.P.; Persiani, A.M. Dynamics of fungi and fungivorous microarthropods in a Mediterranean maquis soil affected by experimental fire. Eur. J. Soil Biol. 2013, 56, 33–34. [Google Scholar] [CrossRef]
- De Marco, A.; Gentile, A.E.; Arena, C.; Virzo De Santo, A. Organic matter, nutrient content and biological activity in burned and unburned soils of a Mediterranean maquis area of southern Italy. Int. J. Wildland Fire 2005, 14, 367–377. [Google Scholar] [CrossRef]
- Wardle, D.A.; Ghani, A. A critique of the microbial metabolic quotient (qCO2) as a bioindicator of disturbance and ecosystem development. Soil Biol. Biochem. 1995, 27, 1601–1610. [Google Scholar] [CrossRef]
- Fierro, A.; Rutigliano, F.A.; De Marco, A.; Castaldi, S.; Virzo De Santo, A. Post-fire stimulation of soil biogenic emission of CO2 in a sandy soil of a Mediterranean shrubland. Int. J. Wildland Fire 2007, 16, 573–583. [Google Scholar] [CrossRef]
- Li, W.; Niu, S.; Liu, X.; Wang, J. Short-term response of the soil bacterial community to differing wildfire severity in Pinus tabulaeformis stands. Sci. Rep. 2019, 9, 1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáenz de Miera, L.E.; Pinto, R.; Gutierrez-Gonzalez, J.J.; Calvo, L.; Ansola, G. Wildfire effects on diversity and composition in soil bacterial communities. Sci. Total Environ. 2020, 726, 138636. [Google Scholar] [CrossRef] [PubMed]
- Andreolli, M.; Lampis, S.; Brignoli, P.; Vallini, G. Bioaugmentation and biostimulation as strategies for the bioremediation of a burned woodland soil contaminated by toxic hydrocarbons: A comparative study. J. Environ. Manag. 2015, 153, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.-A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Burned | Control |
|---|---|---|
| pH | 6.5 (±0.2) b | 6.0 (±0.2) a |
| EC (dS m−1) | 0.40 (±0.06) b | 0.30 (±0.06) a |
| CEC (cmol kg−1 d.w.) | 27.1 (±9.2) | 37.0 (±3.3) |
| Corg (g kg−1 d.w.) | 86.4 (±21.7) | 94.0 (±31.2) |
| Cext (g kg−1 d.w.) | 0.9 (±0.5) | 1.2 (±0.3) |
| Mineralizable C (g CO2-C kg−1 d.w.) | 1.8 (±0.4) | 1.9 (±0.2) |
| Cmic (mg g−1 d.w.) | 1.9 (±1.2) | 1.2 (±0.8) |
| FM (mg g−1 d.w.) | 0.9 (±0.3) | 1.1 (±0.3) |
| Cfung%Cmic | 22.4 (±13.1) | 38.9 (±15.4) |
| Respiration (mg CO2-C g−1 d.w. d−1) | 0.15 (±0.03) | 0.15 (±0.01) |
| qM (CO2-C % Corg) | 2.2 (±0.4) | 2.2 (±0.8) |
| qCO2 (mg CO2-C g−1 Cmic d−1) | 107.7 (±61.2) | 168.0 (±92.5) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stinca, A.; Ravo, M.; Marzaioli, R.; Marchese, G.; Cordella, A.; Rutigliano, F.A.; Esposito, A. Changes in Multi-Level Biodiversity and Soil Features in a Burned Beech Forest in the Southern Italian Coastal Mountain. Forests 2020, 11, 983. https://doi.org/10.3390/f11090983
Stinca A, Ravo M, Marzaioli R, Marchese G, Cordella A, Rutigliano FA, Esposito A. Changes in Multi-Level Biodiversity and Soil Features in a Burned Beech Forest in the Southern Italian Coastal Mountain. Forests. 2020; 11(9):983. https://doi.org/10.3390/f11090983
Chicago/Turabian StyleStinca, Adriano, Maria Ravo, Rossana Marzaioli, Giovanna Marchese, Angela Cordella, Flora A. Rutigliano, and Assunta Esposito. 2020. "Changes in Multi-Level Biodiversity and Soil Features in a Burned Beech Forest in the Southern Italian Coastal Mountain" Forests 11, no. 9: 983. https://doi.org/10.3390/f11090983
APA StyleStinca, A., Ravo, M., Marzaioli, R., Marchese, G., Cordella, A., Rutigliano, F. A., & Esposito, A. (2020). Changes in Multi-Level Biodiversity and Soil Features in a Burned Beech Forest in the Southern Italian Coastal Mountain. Forests, 11(9), 983. https://doi.org/10.3390/f11090983