Seasonal Variation of Soil Respiration in the Mongolian Oak (Quercus mongolica Fisch. Ex Ledeb.) Forests at the Cool Temperate Zone in Korea


Abstract
:1. Introduction
2. Materials and Methods
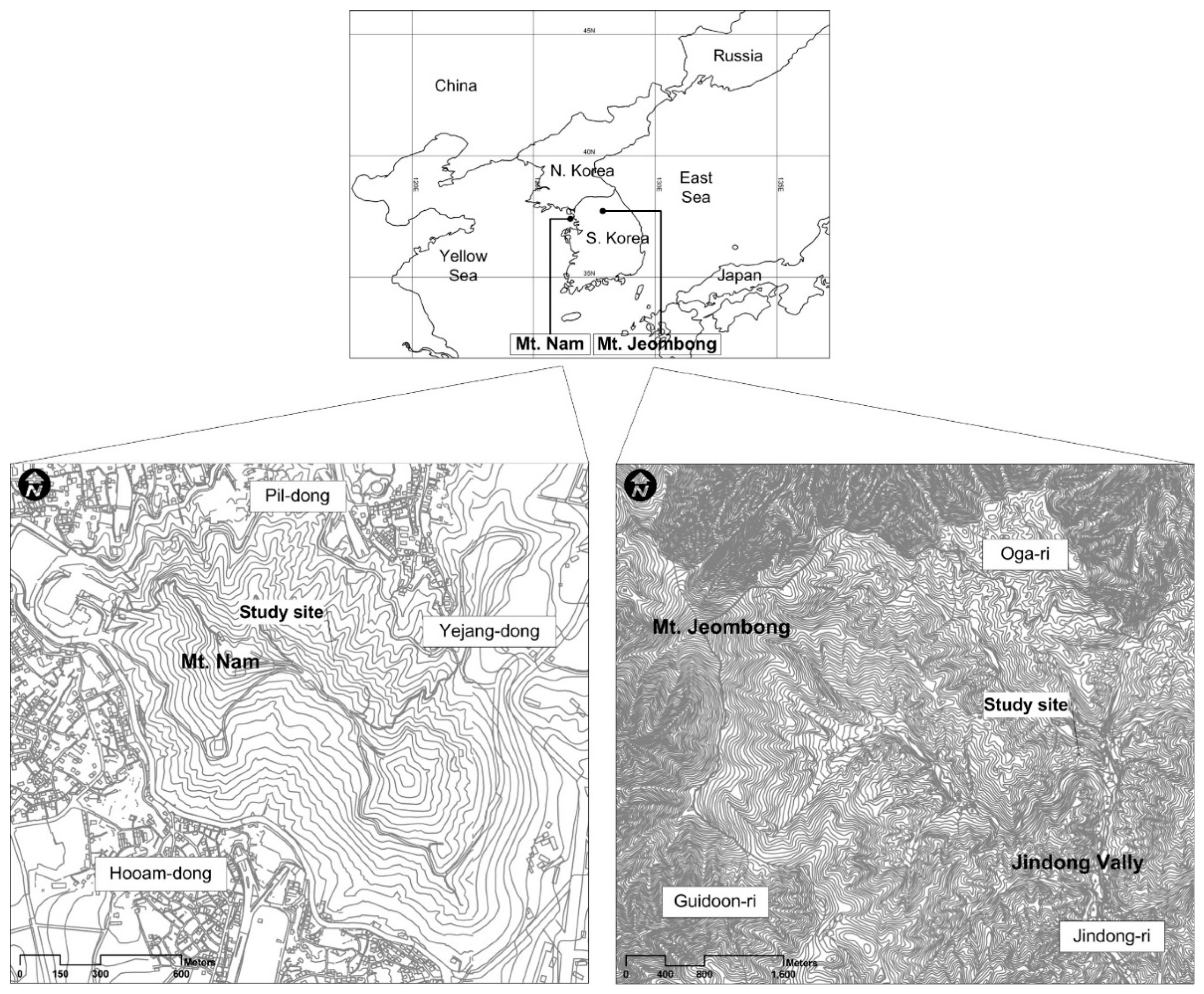
2.1. Study Site
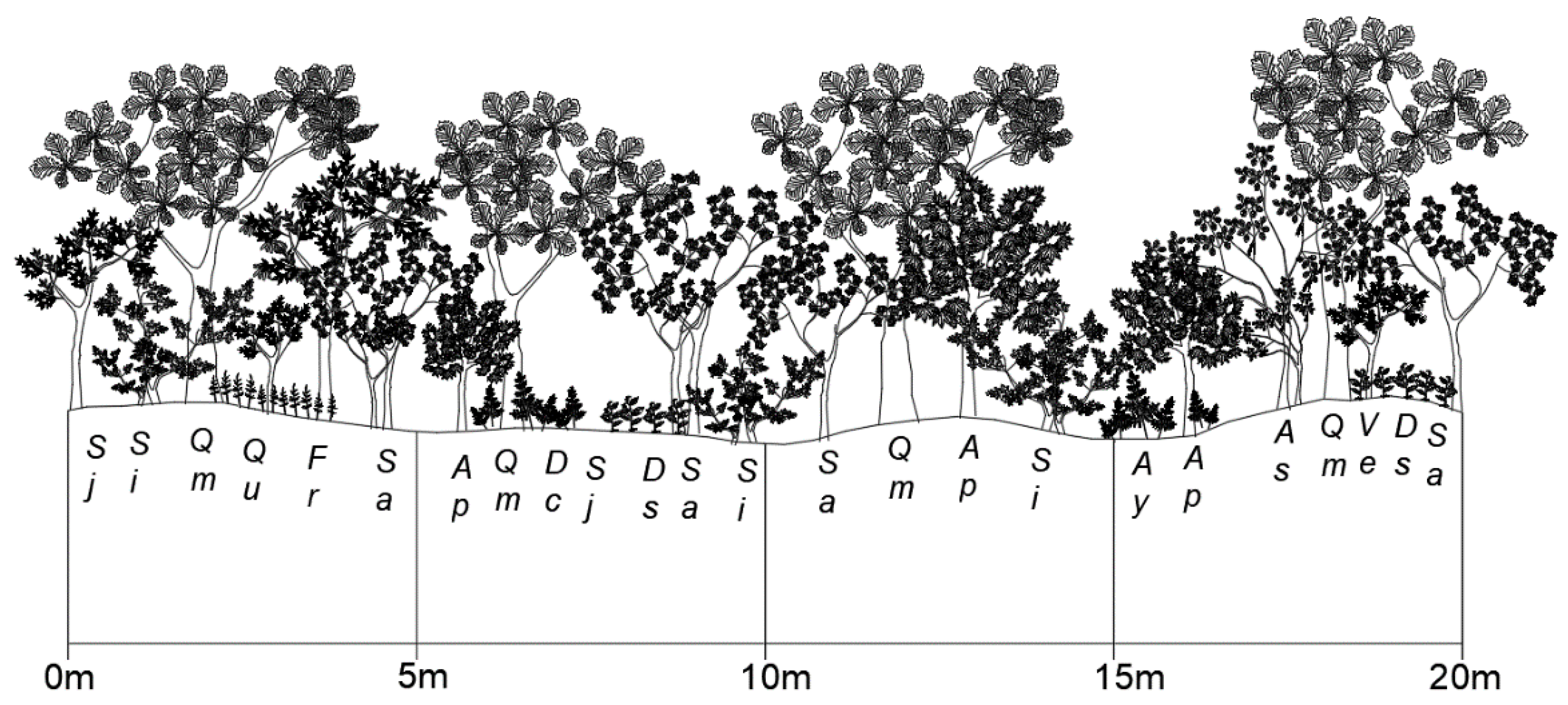
2.1.1. Mt. Nam
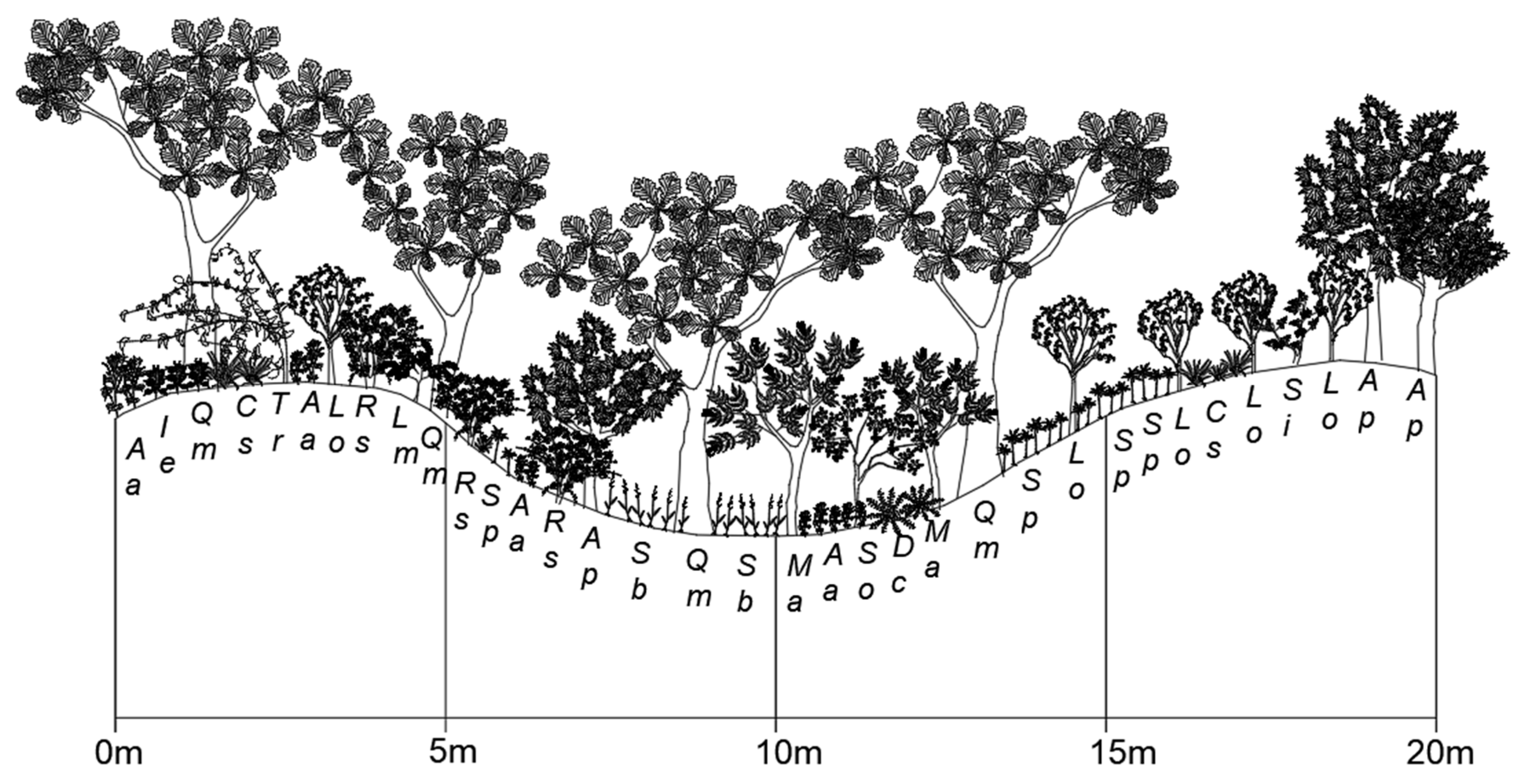
2.1.2. Mt. Jeombong
2.2. Measuring Soil Respiration with an Automated Chamber System
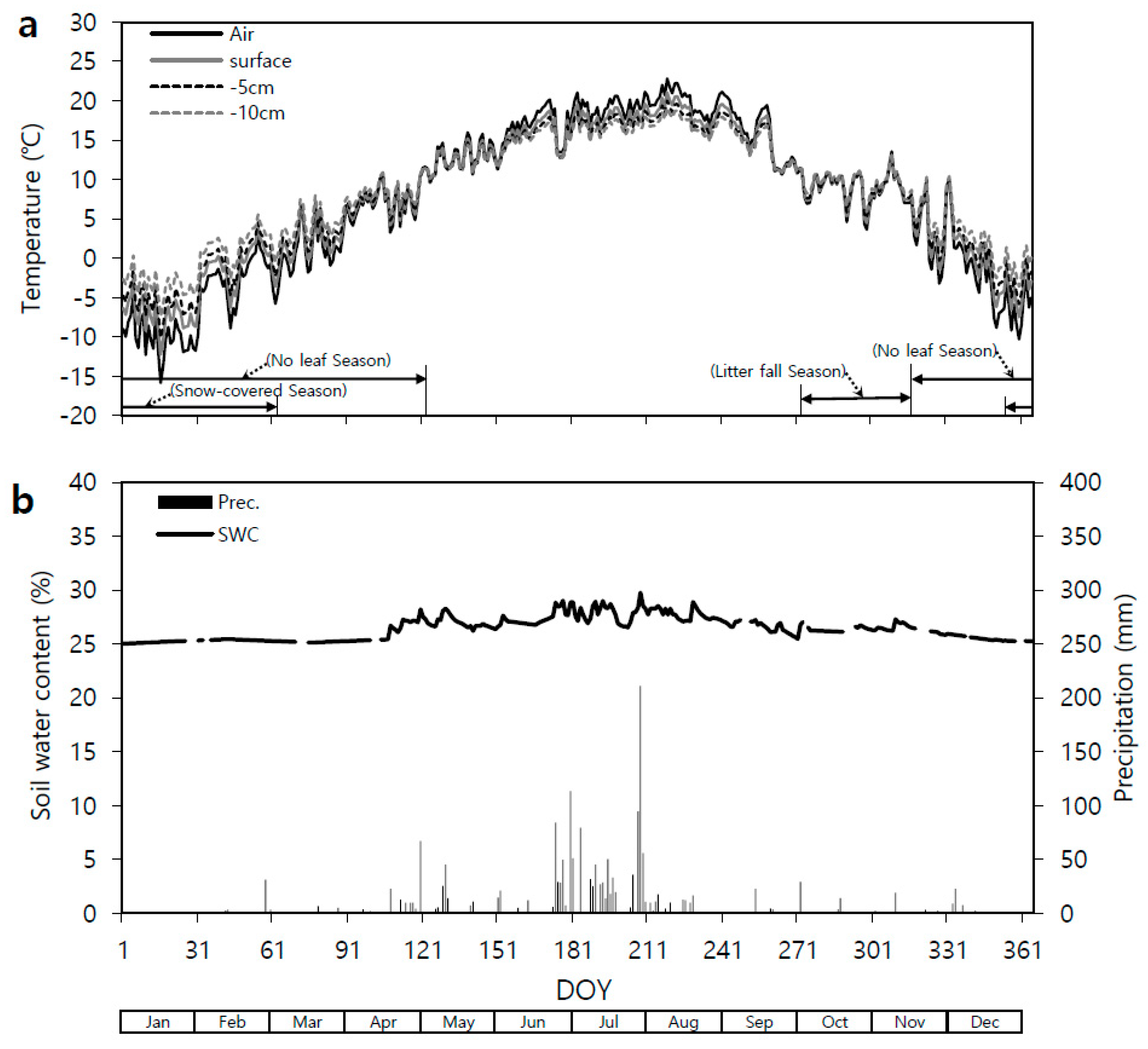
2.3. Measuring Temperature and Soil Water Content
2.4. Data Analysis
3. Results
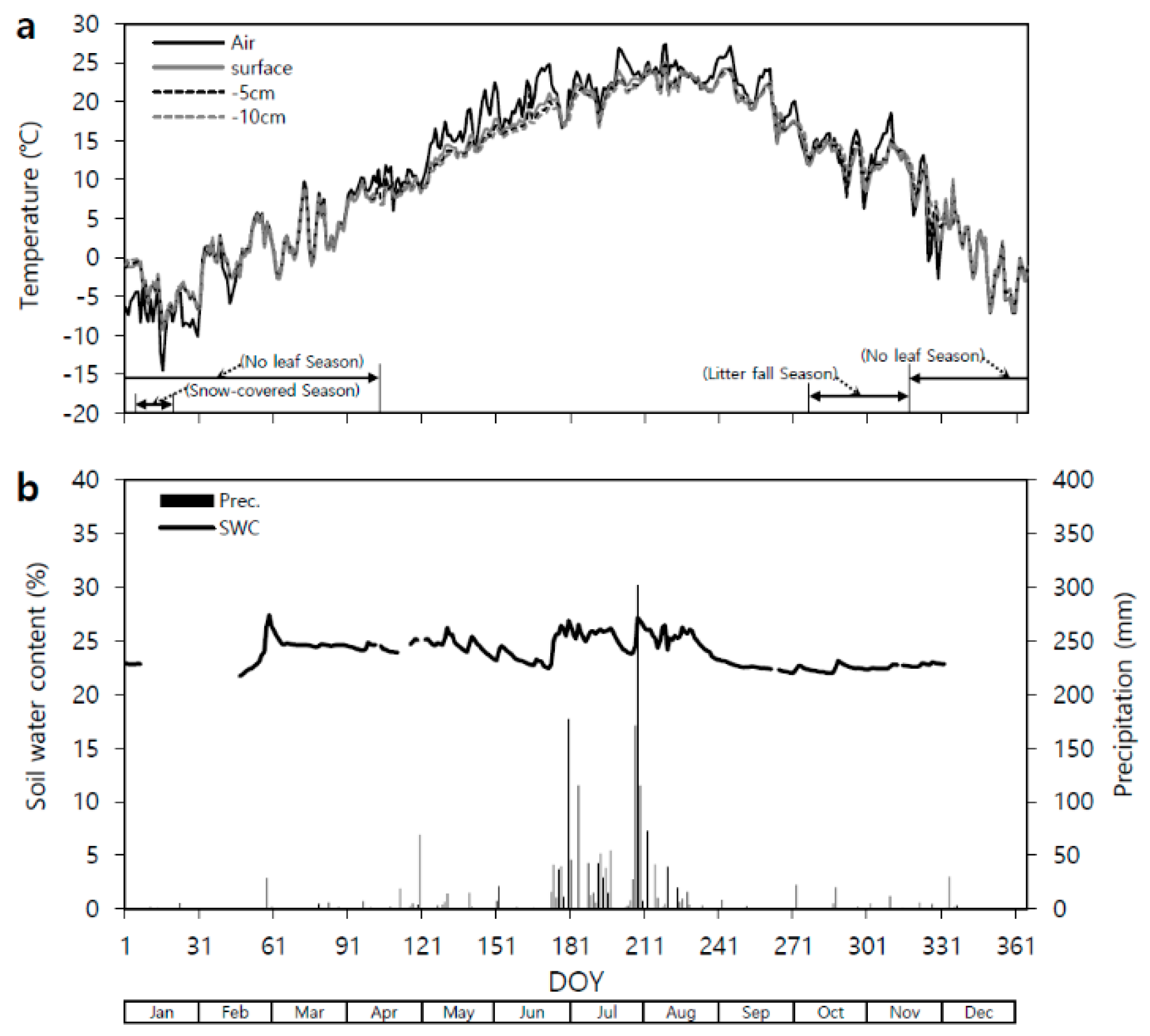
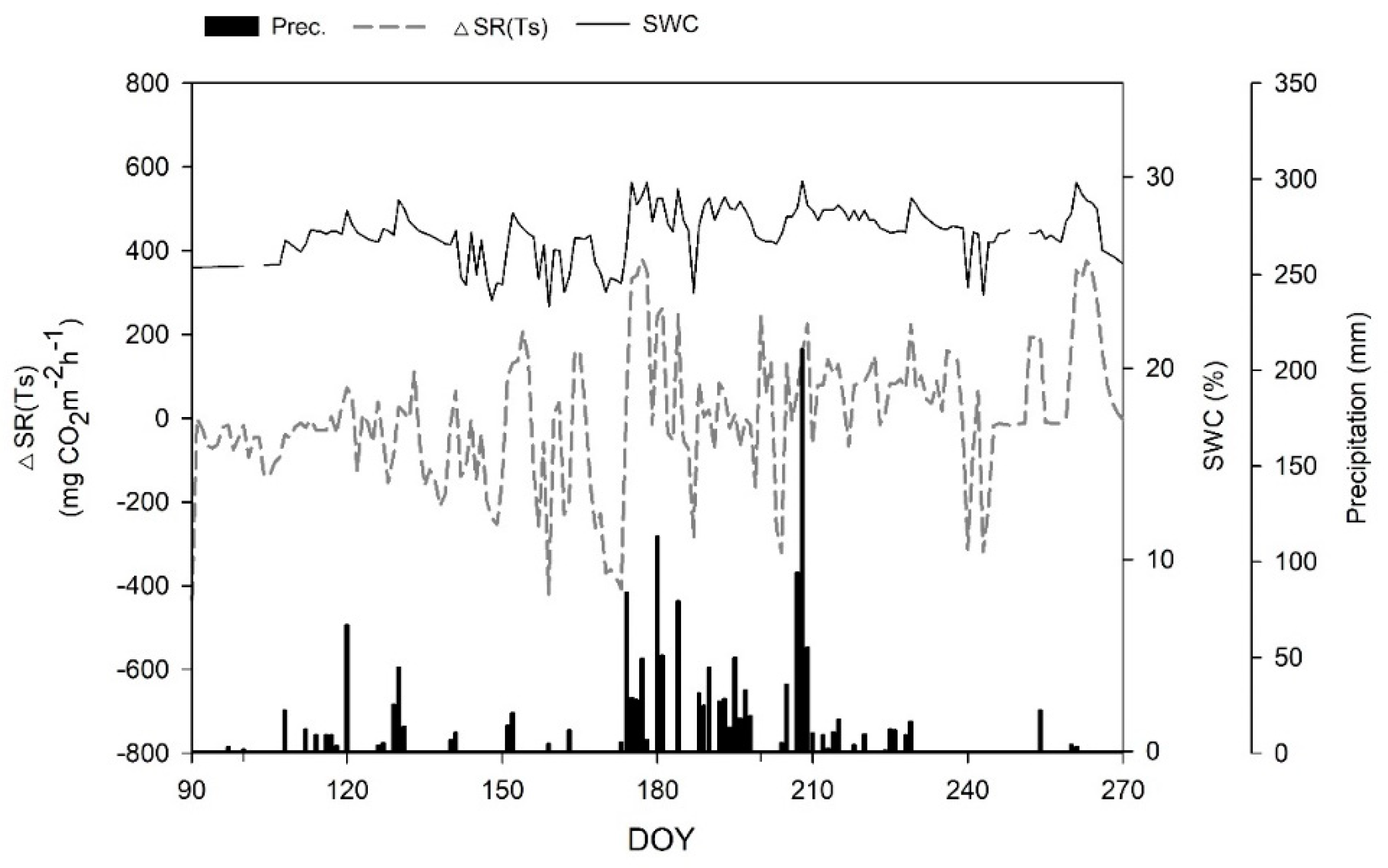
3.1. Seasonal Temperature and Soil Water Content Variation
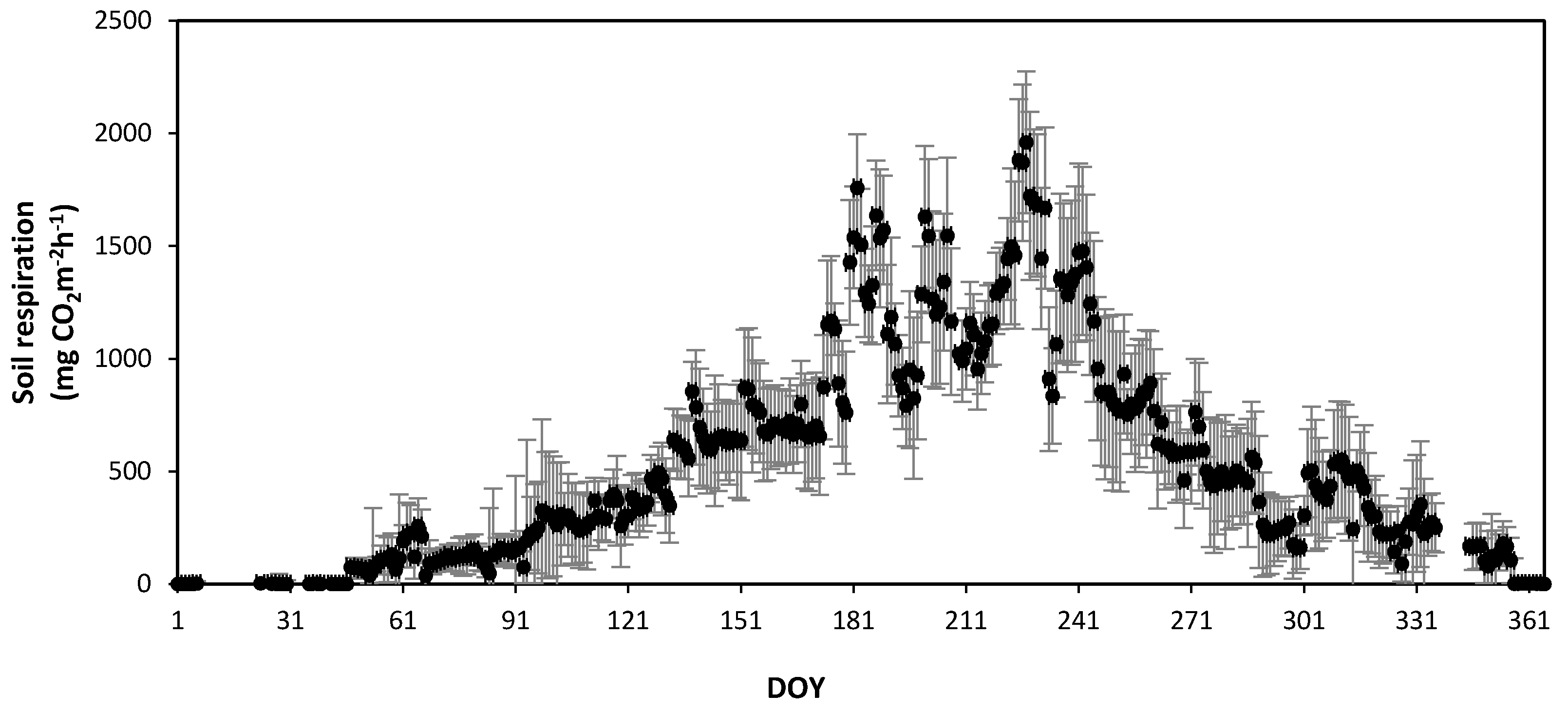
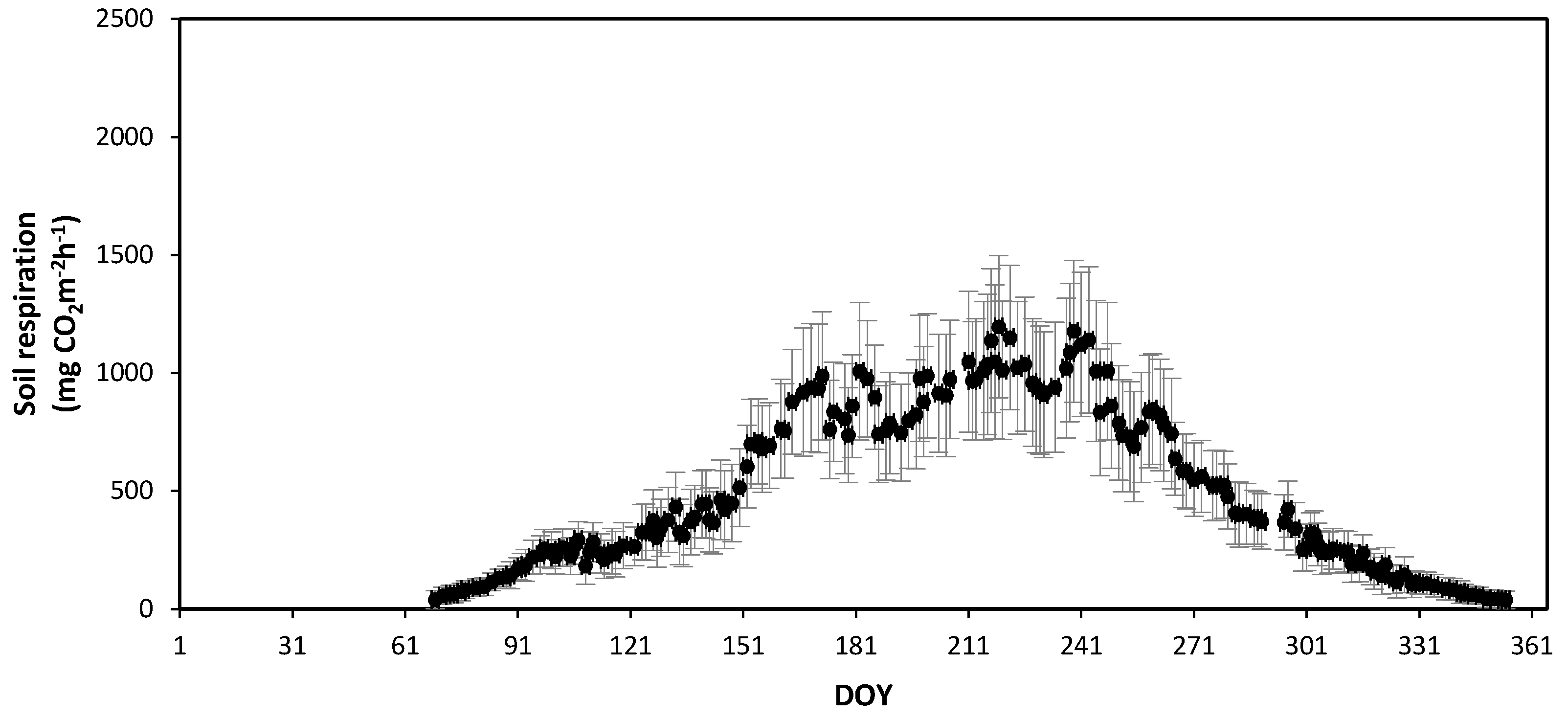
3.2. Seasonal Variations in Soil Respiration
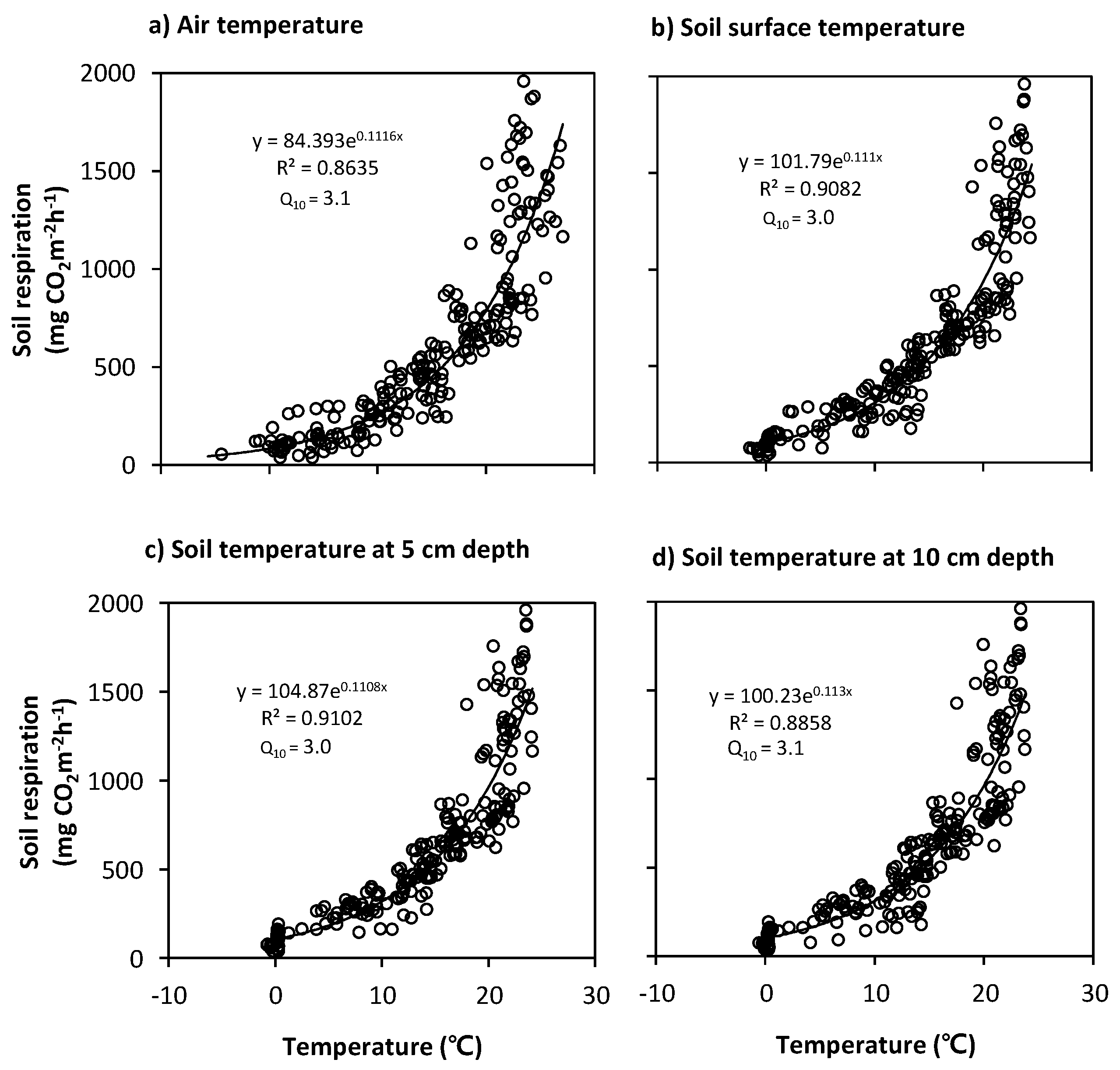
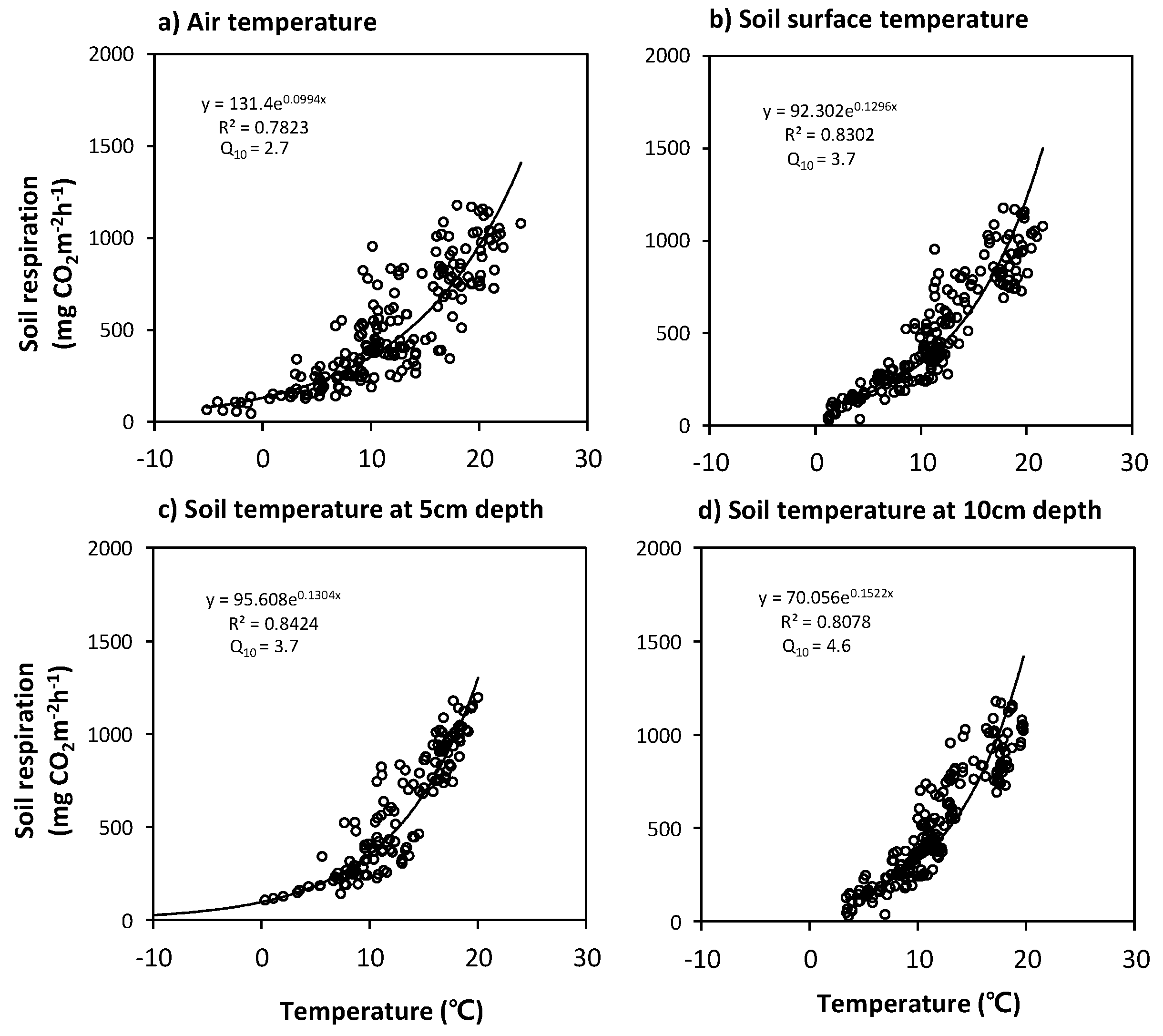
3.3. Relationships between Soil Respiration and Temperature
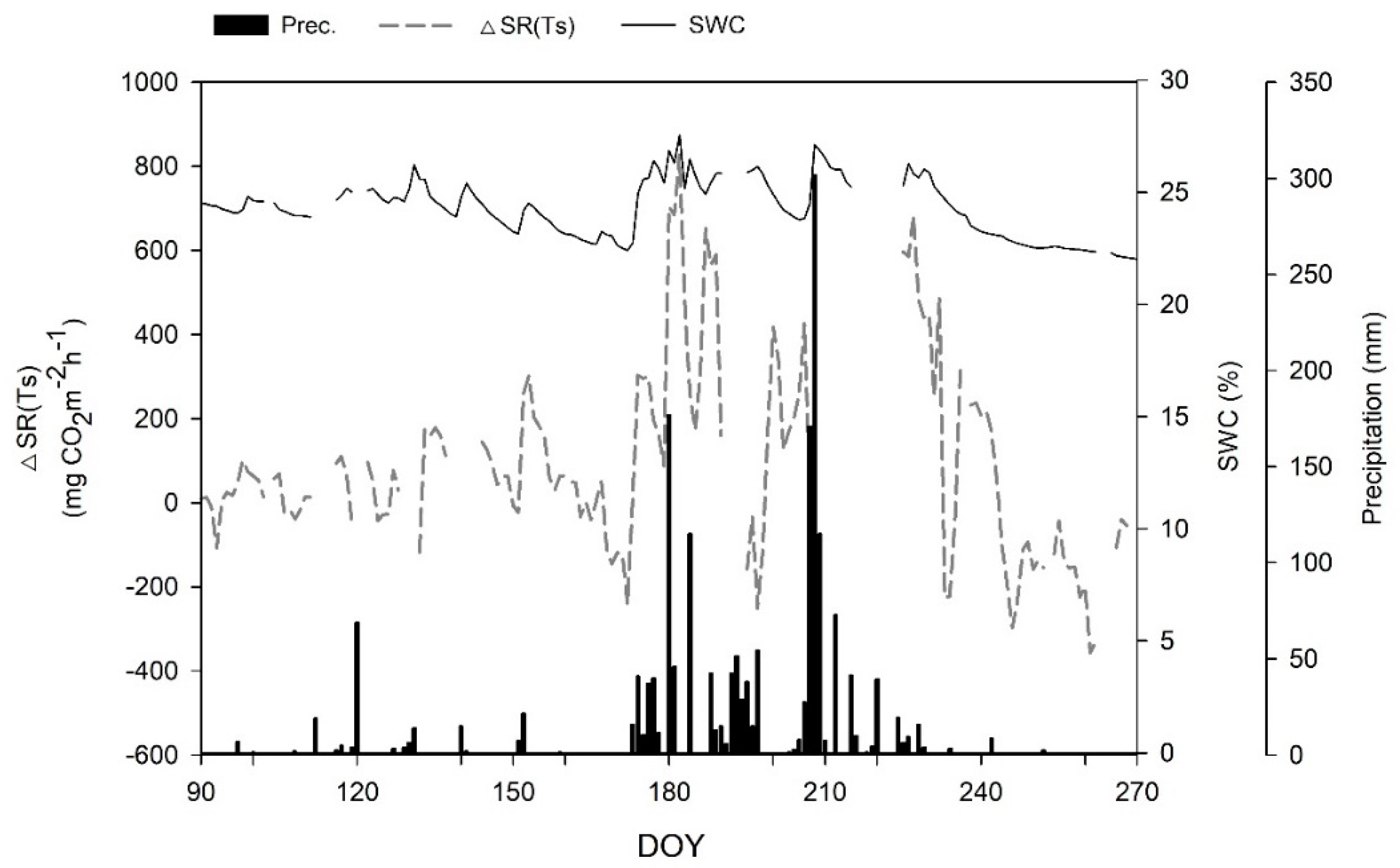
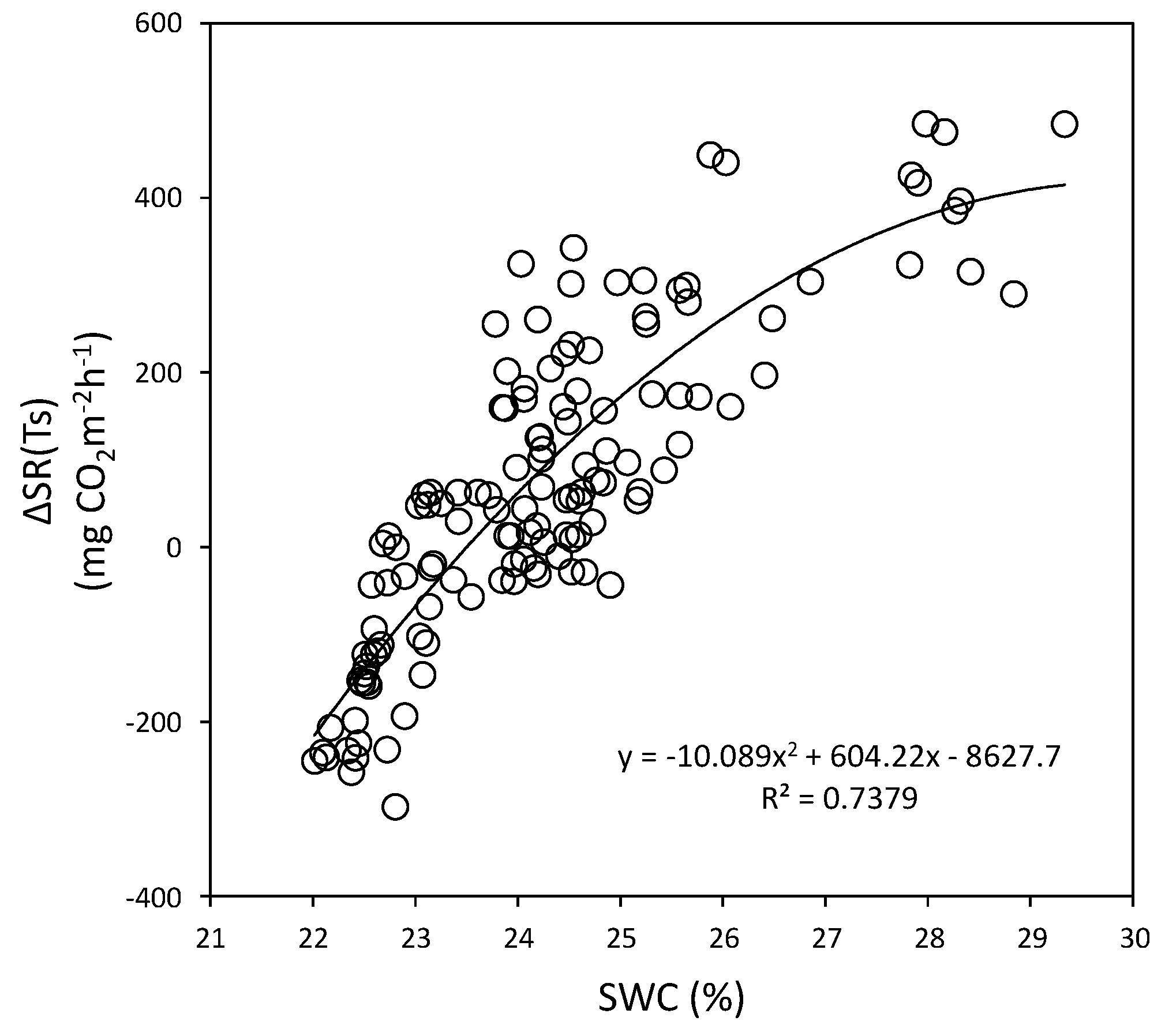
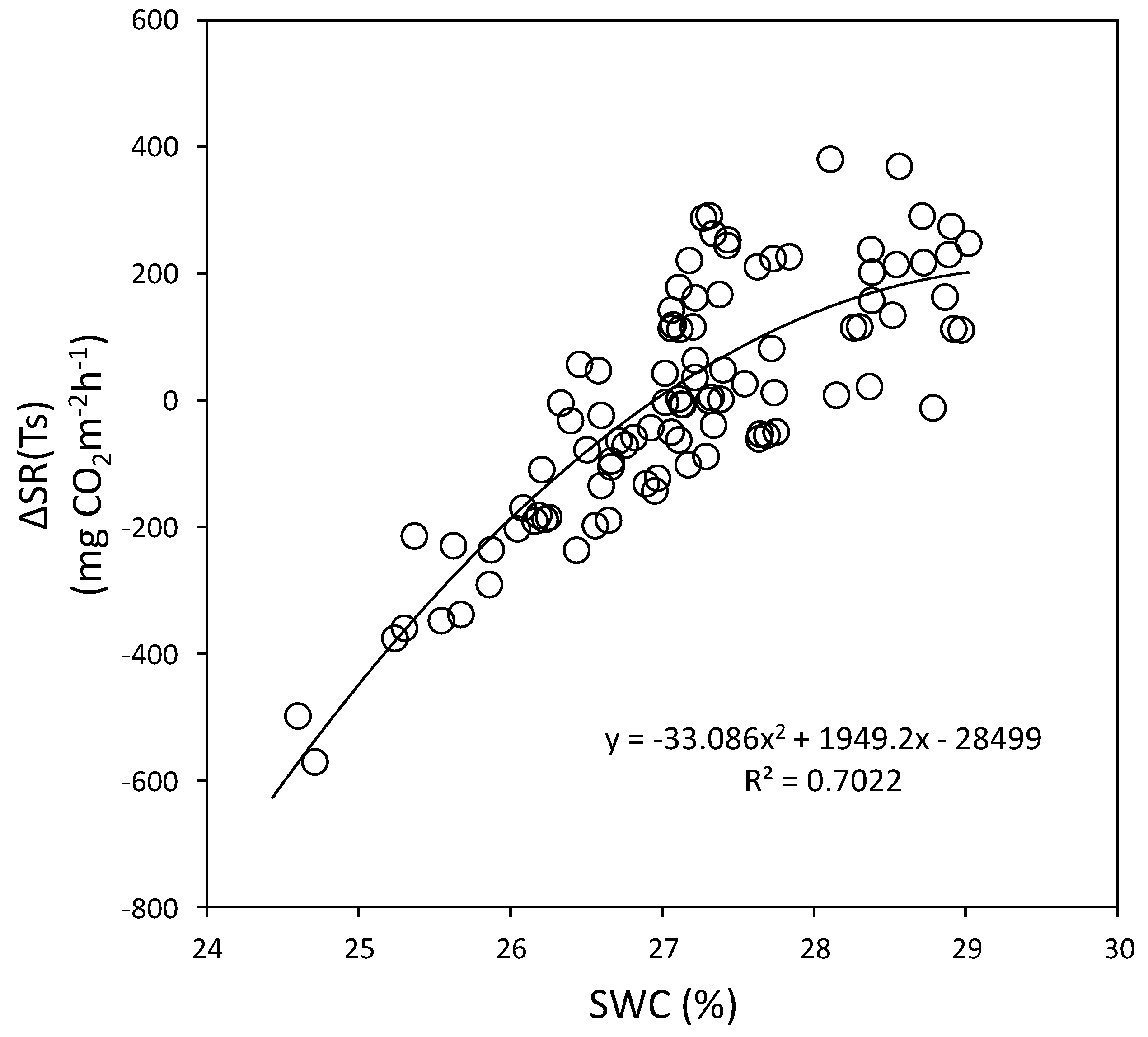
3.4. Relationships between Soil Respiration and Soil Water Content
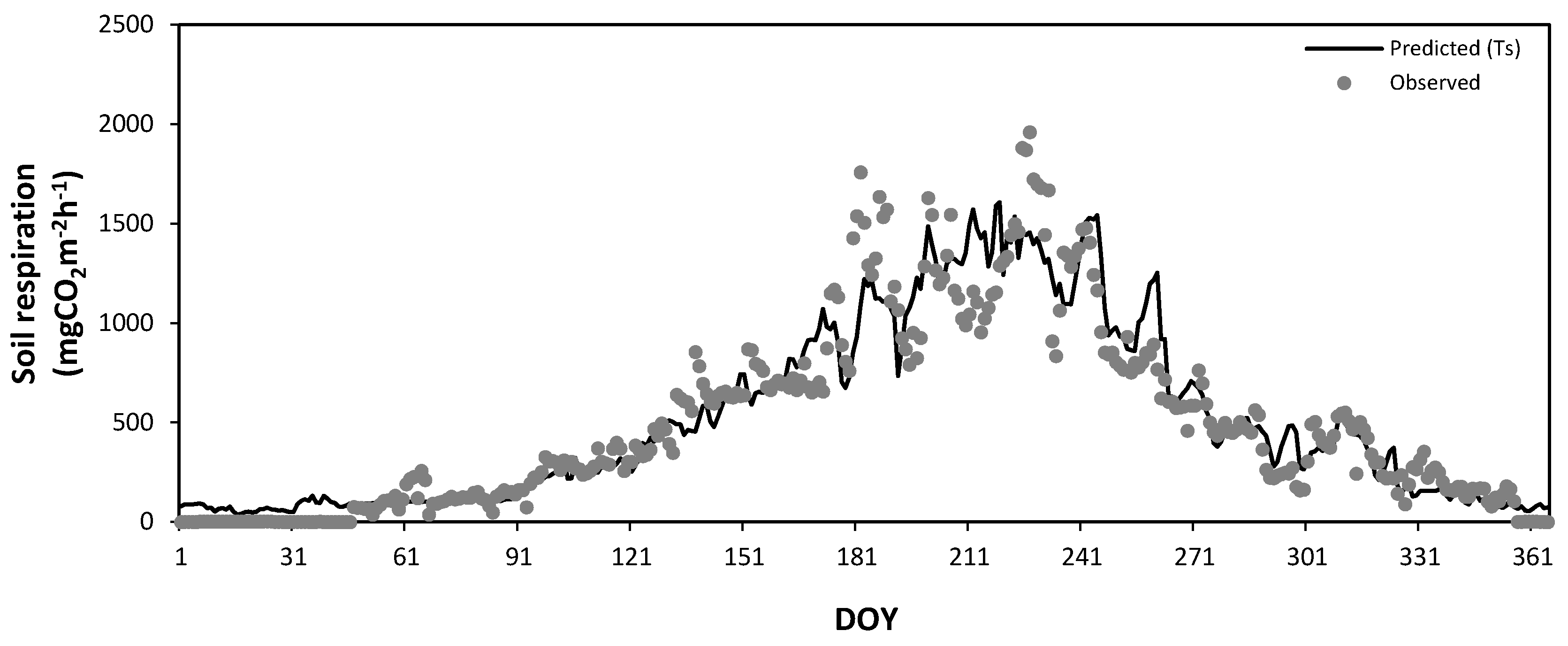
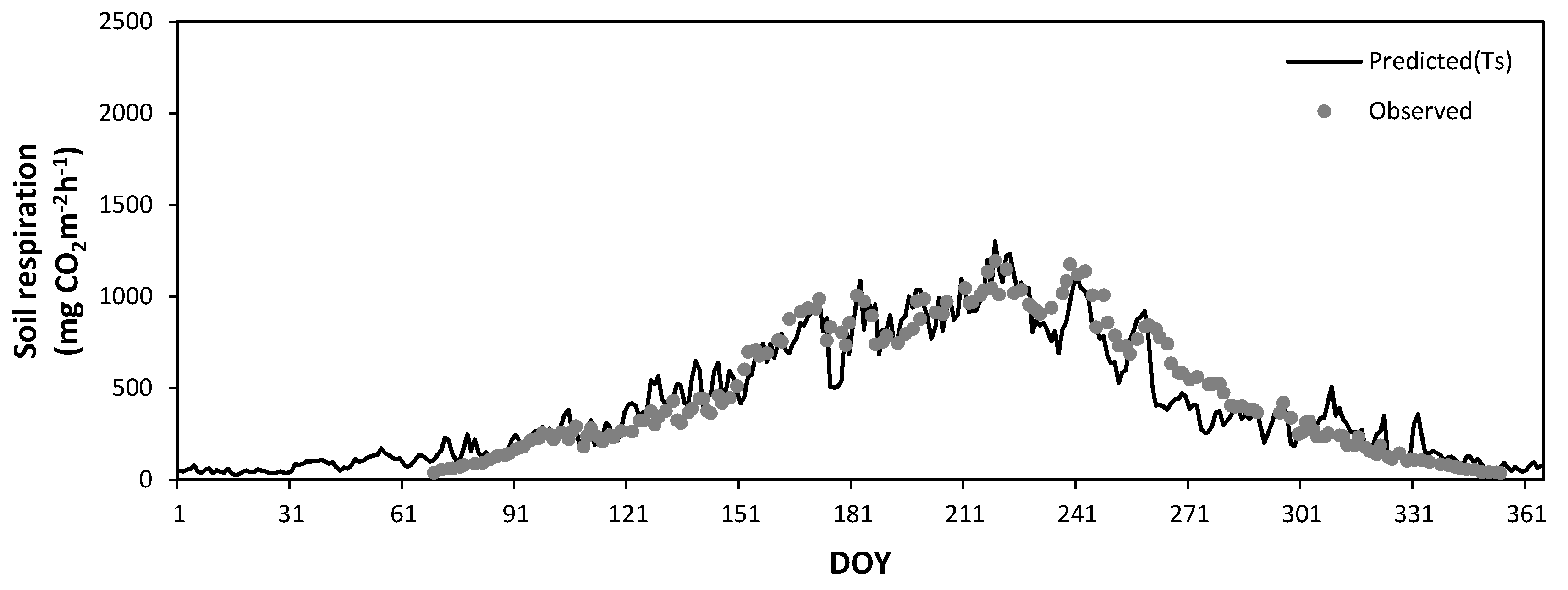
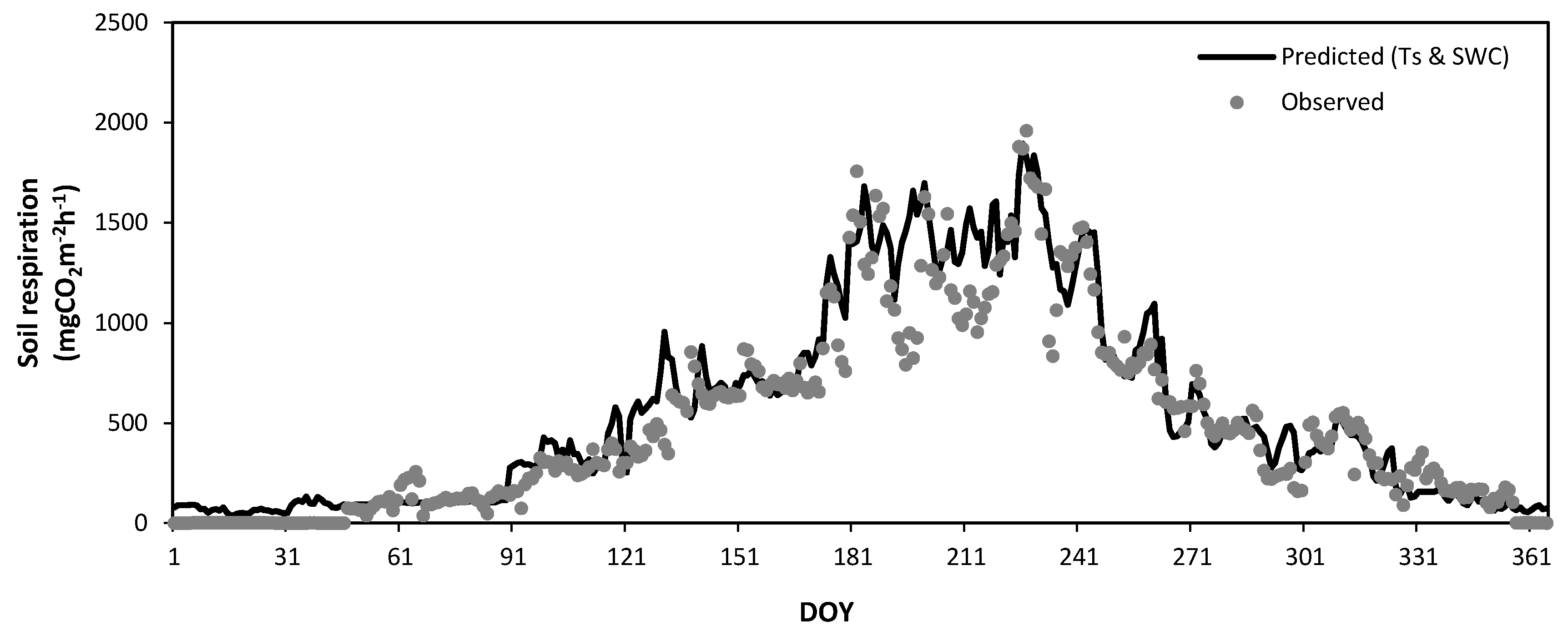
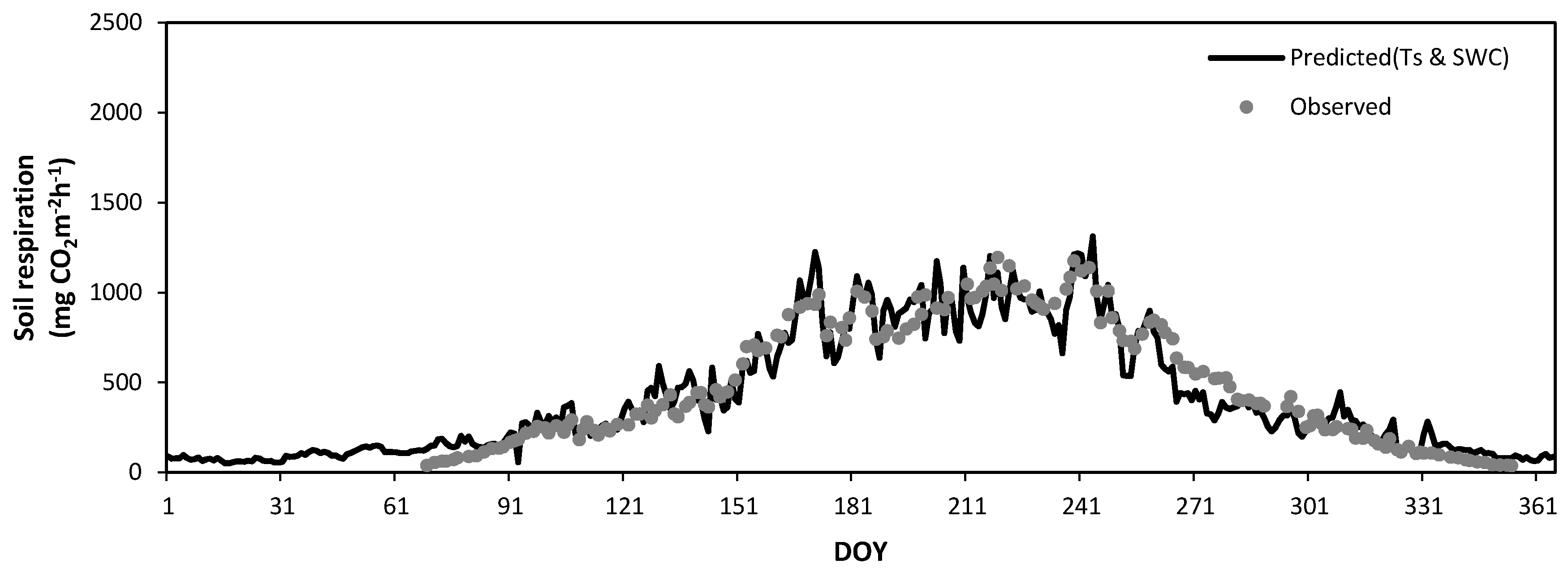
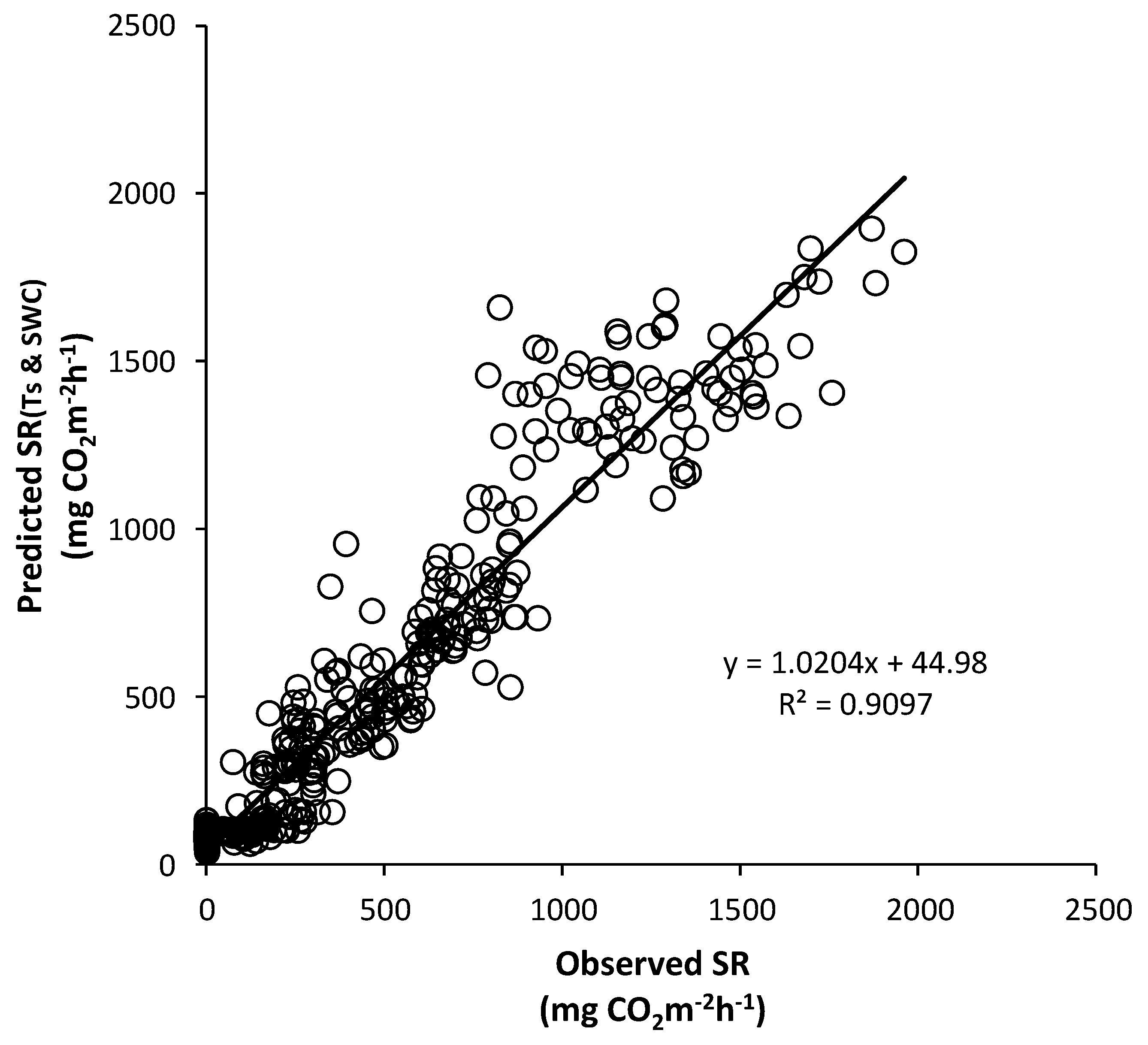
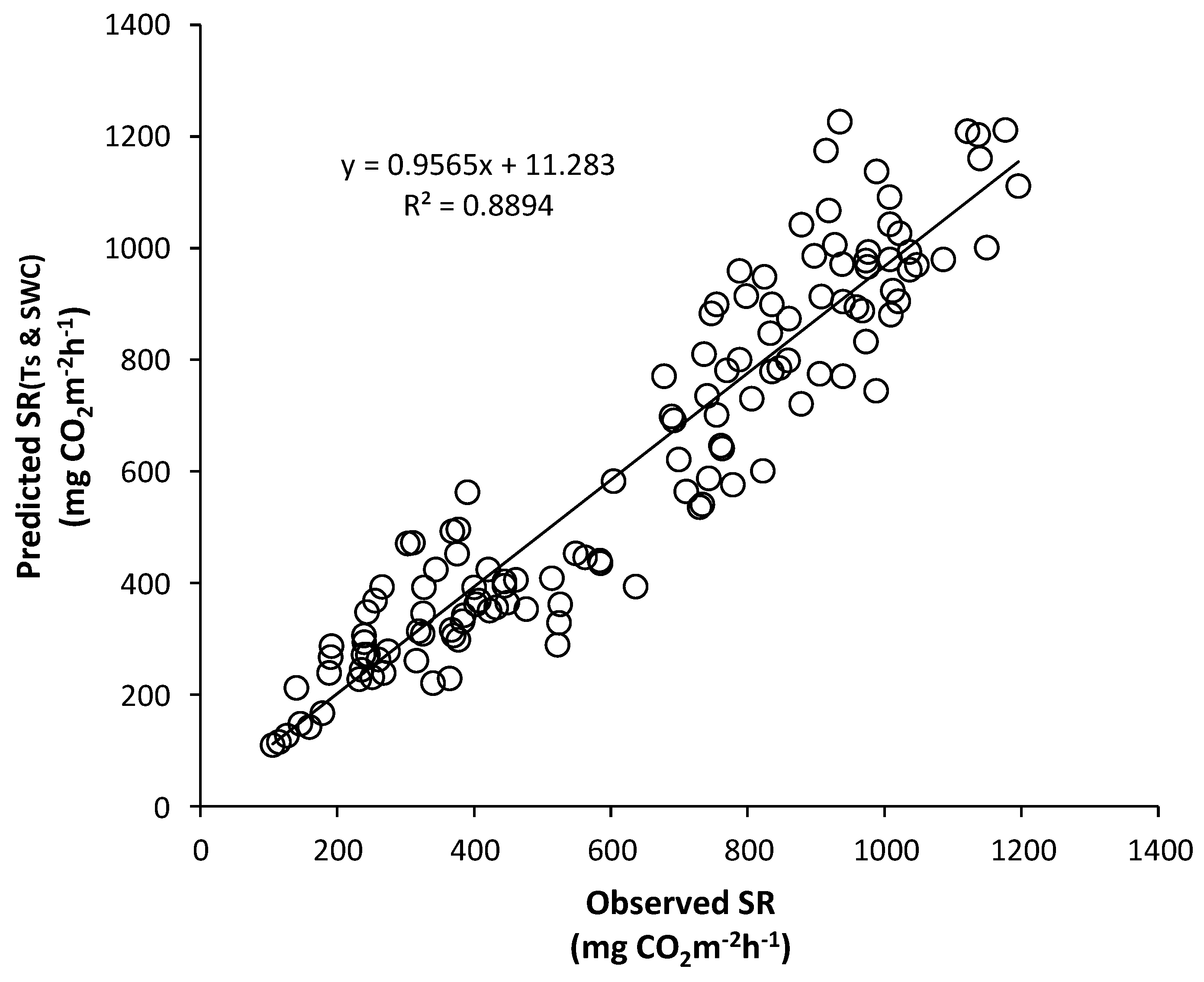
3.5. Seasonal Variation of Soil Respiration Estimated by Soil Temperature and Soil Water Content
3.6. Annual Total Soil Respiration Rates
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Raich, J.W.; Potter, C.S.; Bhagawati, D. Interannual variability in global soil respiration, 1980–1994. Glob. Chang. Biol. 2002, 8, 800–812. [Google Scholar] [CrossRef]
- Trumbore, S. Carbon respired by terrestrial ecosystems-recent progress and challenges. Glob. Chang. Biol. 2006, 12, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Roby, M.C.; Scott, R.L.; Barron-Gafford, G.A.; Hamerlynck, E.P.; Moore, D.J.P. Environmental and vegetative controls on soil CO2 efflux in three semiarid ecosystems. Soil Syst. 2019, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Silletta, L.C.; Cavallaro, A.; Kowal, R.; Pereyra, D.A.; Silva, R.A.; Arias, N.S.; Goldstein, G.; Scholz, F.G.; Bucci, S.J. Temporal and spatial variability in soil CO2 efflux in the Patagonian steppe. Plant Soil 2019, 444, 165–176. [Google Scholar] [CrossRef]
- Cox, P.M.; Betts, R.A.; Jones, C.D.; Spall, S.A.; Totterdell, I.J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 2000, 408, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Goh, K.M. Carbon sequestration and stabilization in soils: Implications for soil productivity and climate change. Soil Sci. Plant Nutr. 2004, 50, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Adair, E.C.; Reich, P.B.; Trost, J.J.; Hobbie, S.E. Elevated CO2 stimulates grassland soil respiration by increasing carbon inputs rather than by enhancing soil moisture. Glob. Chang. Biol. 2011, 17, 3546–3563. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Will changes in soil organic carbon act as a positive or negative feedback on global warming? Biogeochemistry 2000, 48, 21–51. [Google Scholar] [CrossRef]
- Scott-Denton, L.E.; Sparks, K.L.; Monson, R.K. Spatial and temporal controls of soil respiration rate in a high-elevation, subalpine forest. Soil Biol. Biochem. 2003, 35, 525–534. [Google Scholar] [CrossRef]
- Liang, N.; Nakadai, T.; Hirano, T.; Qu, L.; Koike, T.; Fujinuma, Y.; Inoue, G. In site comparison of four approaches to estimating soil CO2 efflux in a northern larch (Larix kaempferi Sarg.) forest. Agric. For. Meteorol. 2004, 123, 97–117. [Google Scholar] [CrossRef]
- Hou, L.; Lei, R.; Liu, J.; Shang, L. Soil respiration in Pinus tabulaeformis forest during dormant period at Huoditang forest zone in the Qinling Mountains, China. Acta Ecol. Sin. 2008, 28, 4070–4077. [Google Scholar]
- Joo, S.J.; Park, M.S.; Kim, G.S.; Lee, C.S. CO2 flux in a cool-temperate deciduous forest (Quercus mongolica) of Mt. Nam in Seoul, Korea. J. Ecol. Field Biol. 2011, 34, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Curtis, P.S.; Hanson, P.J.; Bolstad, P.; Barford, C.; Randolph, J.C.; Schmid, H.P.; Wilson, K.B. Biometric and eddy-covariance based estimates of annual carbon storage in five eastern North American deciduous forests. Agric. For. Meteorol. 2002, 113, 3–19. [Google Scholar] [CrossRef]
- Fang, J.Y.; Guo, Z.; Hu, H.; Kato, T.; Muraoka, H.; Son, Y. Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth. Glob. Chang. Biol. 2014, 20, 2019–2030. [Google Scholar] [CrossRef] [PubMed]
- Qubaja, R.; Grünzweig, J.M.; Rotenberg, E.; Yakir, D. Evidence for large carbon sink and long residence time in semiarid forests based on 15 year flux and inventory records. Glob. Chang. Biol. 2020, 26, 1626–1637. [Google Scholar] [CrossRef]
- LIN, B.; Jiamin, G. Carbon sinks and output of China’s forestry sector: An ecological economic development perspective. Sci. Total Environ. 2019, 655, 1169–1180. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Rustad, L.E.; Huntington, T.G.; Boone, R.D. Controls on soil respiration: Implications for climate change. Biogeochemistry 2000, 48, 1–6. [Google Scholar] [CrossRef]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Bowden, R.D.; Nadelhoffer, K.J.; Boone, R.D.; Melillo, J.M.; Garrison, J.B. Contributions of aboveground litter, belowground litter, and root respiration to total soil respiration in a mixed hardwood forest. Can. J. For. Res. 1993, 23, 1402–1407. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- D’Andrea, E.; Guidolotti, G.; Scartazza, A.; De Angelis, P.; Matteucci, G. Small-Scale Forest Structure Influences Spatial Variability of Belowground Carbon Fluxes in A Mature Mediterranean Beech Forest. Forests 2020, 11, 255. [Google Scholar] [CrossRef] [Green Version]
- Law, B.E.; Ryan, M.G.; Anthoni, P.M. Seasonal and annual reparation of a ponderosa pine ecosystem. Glob. Chang. Biol. 1999, 5, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Janssens, I.A.; Matteucci, H.G.; Kowalski, A.S.; Buchman, N.; Epron, D.; Pilegaard, K.; Kutsch, W.; Longdoz, B.; Grunwald, T. Productivity overshadows temperature in determining soil ecosystem reparation across European forests. Glob. Chang. Biol. 2001, 7, 269–278. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.J.; Luo, Y. On the variability of respiration in terrestrial ecosystems: Moving beyond Q10. Glob. Chang. Biol. 2006, 12, 154–164. [Google Scholar] [CrossRef]
- Mo, W.; Lee, M.S.; Masaki, U.; Motoko, I. Seasonal and annual variations in soil respiration in a cool-temperate deciduous broad-leaved forest in Japan. Agric. For. Meteorol. 2005, 134, 81–94. [Google Scholar] [CrossRef]
- Reich, P.B. The carbon dioxide exchange. Science 2010, 329, 774–775. [Google Scholar] [CrossRef] [Green Version]
- Raich, J.W.; Tufekcioglu, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Li, H.J.; Yan, J.X.; Yue, X.F.; Wang, M.B. Significance of soil temperature and moisture for soil respiration in a Chinese mountain area. Agric. For. Meteorol. 2008, 148, 490–503. [Google Scholar] [CrossRef]
- Fenn, K.M.; Malhi, Y.; Morecroft, M.D. Soil CO2 efflux in a temperate deciduous forest: Environmental drivers and component contributions. Soil Biol. Biochem. 2010, 42, 1685–1693. [Google Scholar] [CrossRef]
- Estruch, C.; Macek, P.; Armas, C.; Pistón, N.; Pugnaire, F.I. Species identity improves soil respiration predictions in a semiarid scrubland. Geoderma 2020, 363, 114153. [Google Scholar] [CrossRef]
- Zhang, H.; Qian, Z.; Zhuang, S. Effects of Soil Temperature, Water Content, Species, and Fertilization on Soil Respiration in Bamboo Forest in Subtropical China. Forests 2020, 11, 99. [Google Scholar] [CrossRef] [Green Version]
- Lingfei, Y.; Wang, H.; Wang, Y.; Zhang, Z.; Chen, L.; Liang, N.; He, J.S. Temporal variation in soil respiration and its sensitivity to temperature along a hydrological gradient in an alpine wetland of the Tibetan Plateau. Agric. For. Meteorol. 2020, 282, 107854. [Google Scholar]
- Zhao, P.; Pumpanen, J.; Kang, S. Spatio-temporal variability and controls of soil respiration in a furrow-irrigated vineyard. Soil Tillage Res. 2020, 196, 104424. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.J. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.; Taylor, J.A. On the temperature-dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Wan, S.; Norby, R.J.; Ledford, J. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Glob. Chang. Biol. 2007, 13, 2411–2424. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment; Elsevier Academic Press: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Borken, W.; Savage, K.; Davidson, E.A.; Trumbore, S.E. Effects of experimental drought on soil respiration and radiocarbon efflux from a temperate forest soil. Glob. Chang. Biol. 2006, 12, 177–193. [Google Scholar] [CrossRef] [Green Version]
- Kwak, Y.S.; Kim, J.H. Secular changes of density, litterfall, phytomass and primary productivity in Mongolian oak (Quercus mongolica) forest. Korean J. Ecol. 1992, 15, 19–33. [Google Scholar]
- Son, Y.; Jun, Y.C.; Lee, Y.Y.; Kim, R.H.; Yang, S.Y. Soil carbon dioxide evolution, litter decomposition, and nitrogen availability four years after thinning in a Japanese larch plantation. Commun. Soil Sci. Plant Anal. 2004, 35, 1111–1122. [Google Scholar] [CrossRef]
- Kim, W.S.; Cho, J.I.; Myong, G.M.; Mano, M.; Komori, D.; Kim, S.D. Quality Assessment of Data from the Daegwallyeong Flux Measurement Station (DFMS) Based on Short-term Experiments. J. Agric. Meteorol. 2008, 64, 111–120. [Google Scholar] [CrossRef]
- Yi, M.J. Soil CO2 evolution in Quercus variabilis and Q. mongolica forests in Chunchon, Kangwon province. J. Korean For. Soc. 2003, 92, 263–269. [Google Scholar]
- Kang, S.; Doh, S.; Lee, D.; Lee, D.; Jin, V.L.; Kimball, J.S. Topographic and climatic controls on soil respiration in six temperate mixed-hardwood forest slopes, Korea. Glob. Chang. Biol. 2003, 9, 1427–1437. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C. Effects of micro-environment in ridge and southern slope on soil respiration in Quercus mongolica forest. J. Ecol. Environ. 2018, 42, 26. [Google Scholar] [CrossRef]
- Kim, G.S.; Song, H.K.; Lee, C.H.; Cho, H.J.; Lee, C.S. Ecological comparison of Mongolian oak (Quercus mongolica Fisch. Ex Ledeb.) community between Mt. Nam and Mt. Jeombong as a Long Term Ecological Research (LTER) site. J. Ecol. Field Biol. 2011, 34, 75–85. [Google Scholar] [CrossRef]
- Lee, C.S.; Moon, J.S.; Hwangbo, J.K.; You, Y.H. Selection of pollution-tolerant plants and restoration planning to recover the forest ecosystem degraded by air pollution in the industrial complex. Korean J. Biol. Sci. 2002, 6, 59–64. [Google Scholar] [CrossRef]
- Lee, C.S.; Kim, J.H.; Yi, H.; You, Y.H. Seedling establishment and regeneration of Korean red pine (Pinus densiflora S. et Z.) forests in Korea. For. Ecol. Manag. 2004, 199, 423–432. [Google Scholar] [CrossRef]
- FAO-UNESCO. World Reference Base for Soil Resources; FAO-UNESCO: Rome, Italy, 1998. [Google Scholar]
- Kim, G.Z.; Kim, J.H. Evaluation of Ecological Niche for Major Tree Species in the Natural Deciduous Forest of Mt. Chumbong. J. Korean Soc. For. Sci. 2001, 90, 380–387. [Google Scholar]
- Xu, M.; Qi, Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Glob. Chang. Biol. 2001, 7, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Lytle, D.E.; Cronan, C.S. Comparative soil CO2 evolution, litter decay, and root dynamics in clear cut and uncut spruce fir forest. For. Ecol. Manag. 1998, 103, 121–128. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Wang, W.J.; Dalal, R.C.; Moody, P.W.; Smith, C.J. Relationships of soil respiration to microbial biomass, substrate availability and clay content. Soil Biol. Biochem. 2003, 35, 273–284. [Google Scholar] [CrossRef]
- Eberwein, J.R.; Oikawa, P.Y.; Allsman, L.A.; Jenerette, G.D. Carbon availability regulates soil respiration response to nitrogen and temperature. Soil Biol. Biochem. 2015, 88, 158–164. [Google Scholar] [CrossRef]
- Raich, J.W.; Schlesinger, W.H. The global carbon-dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus. B Chem. Phys. Meteorol. 1992, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Uchida, M.; Mo, W.; Nakatsubo, T.; Tsuchiya, Y.; Horikoshi, T.; Koizumi, H. Microbial activity and litter decomposition under snow cover in a cool-temperate broad-leaved deciduous forest. Agric. For. Meteorol. 2005, 134, 102–109. [Google Scholar] [CrossRef]
- Pumpanen, J.; Ilvesnemi, H.; Peramaki, M.; Hari, P. Seasonal patterns of soil CO2 efflux and soil air CO2 concentration in a Scots pine forest: Comparison of two chamber techniques. Glob. Chang. Biol. 2003, 9, 371–381. [Google Scholar] [CrossRef]
- Billings, S.A.; Richter, D.D.; Yarie, J. Soil carbon dioxide fluxes and profile concentrations in two boreal forests. Can. J. For. Res. 1998, 28, 1773–1783. [Google Scholar] [CrossRef]
- Cook, F.J.; Orchard, V.A. Relationships between soil respiration and soil moisture. Soil Biol. Biochem. 2008, 40, 1013–1018. [Google Scholar] [CrossRef]
- Skopp, J.; Jawson, M.D.; Doran, J.W. Steady-state aerobicmicrobial activity as a function of soil water content. Soil Sci. Soc. Am. J. 1990, 54, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Curiel Youste, J.; Janssens, J.; Carrara, I.A.; Meiresonne, A.; Ceulemans, L. Interactive effects of temperature and precipitation on soil respiration in a temperate maritime pine forest. Tree Physiol. 2003, 23, 1263–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogberg, P.; Nordgren, A.; Buchmann, N. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.; Pegoraro, E.; Tedeschi, V.; de Parri, I.; Jarvis, P.G.; Valentini, R. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Glob. Chang. Biol. 2002, 8, 851–866. [Google Scholar] [CrossRef]
- Kordowski, K.; Kuttler, W. Carbon dioxide fluxes over an urban park area. Atmos. Environ. 2010, 44, 2722–2730. [Google Scholar] [CrossRef]
- Park, M.S.; Joo, S.J.; Lee, C.S. Effects of an urban park and residential area on the atmospheric CO2 concentration and flux in Seoul Korea. Adv. Atmos. Sci. 2013, 30, 503–514. [Google Scholar] [CrossRef]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed SR | Predicted SR by Ts | Predicted SR by Ts and SWC | |
|---|---|---|---|
| g C m−2 yr−1 | g C m−2 yr−1 | g C m−2 yr−1 | |
| Jan–Mar | 34.5 (25) | 52.8 | 53.8 |
| Apr–Jun | 328.1 (17) | 306.9 | 372.6 |
| Jul–Sep | 678.6 (15) | 704.1 | 746.7 |
| Oct–Dec | 166.2 (21) | 166.3 | 166.3 |
| Annual total | 1207.3 (78) | 1230.0 | 1339.4 |
| Observed SR | Predicted SR by Ts | Predicted SR by Ts and SWC | |
|---|---|---|---|
| g C m−2 yr−1 | g C m−2 yr−1 | g C m−2 yr−1 | |
| Jan–Mar | 25.3 (75) | 36.4 | 37.2 |
| Apr–Jun | 285.0 (51) | 309.3 | 289.1 |
| Jul–Sep | 538.9 (36) | 544.7 | 556.2 |
| Oct–Dec | 126.7 (33) | 120.5 | 120.5 |
| Annual total | 975.9 (195) | 1010.9 | 1003.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.S.; Joo, S.J.; Lee, C.S. Seasonal Variation of Soil Respiration in the Mongolian Oak (Quercus mongolica Fisch. Ex Ledeb.) Forests at the Cool Temperate Zone in Korea. Forests 2020, 11, 984. https://doi.org/10.3390/f11090984
Kim GS, Joo SJ, Lee CS. Seasonal Variation of Soil Respiration in the Mongolian Oak (Quercus mongolica Fisch. Ex Ledeb.) Forests at the Cool Temperate Zone in Korea. Forests. 2020; 11(9):984. https://doi.org/10.3390/f11090984
Chicago/Turabian StyleKim, Gyung Soon, Seung Jin Joo, and Chang Seok Lee. 2020. "Seasonal Variation of Soil Respiration in the Mongolian Oak (Quercus mongolica Fisch. Ex Ledeb.) Forests at the Cool Temperate Zone in Korea" Forests 11, no. 9: 984. https://doi.org/10.3390/f11090984
APA StyleKim, G. S., Joo, S. J., & Lee, C. S. (2020). Seasonal Variation of Soil Respiration in the Mongolian Oak (Quercus mongolica Fisch. Ex Ledeb.) Forests at the Cool Temperate Zone in Korea. Forests, 11(9), 984. https://doi.org/10.3390/f11090984