Tree-Ring Analysis of Intermediate Hawthorn (Crataegus media Bechst.) in NW Poland


Abstract
:1. Introduction
2. Material and Methods
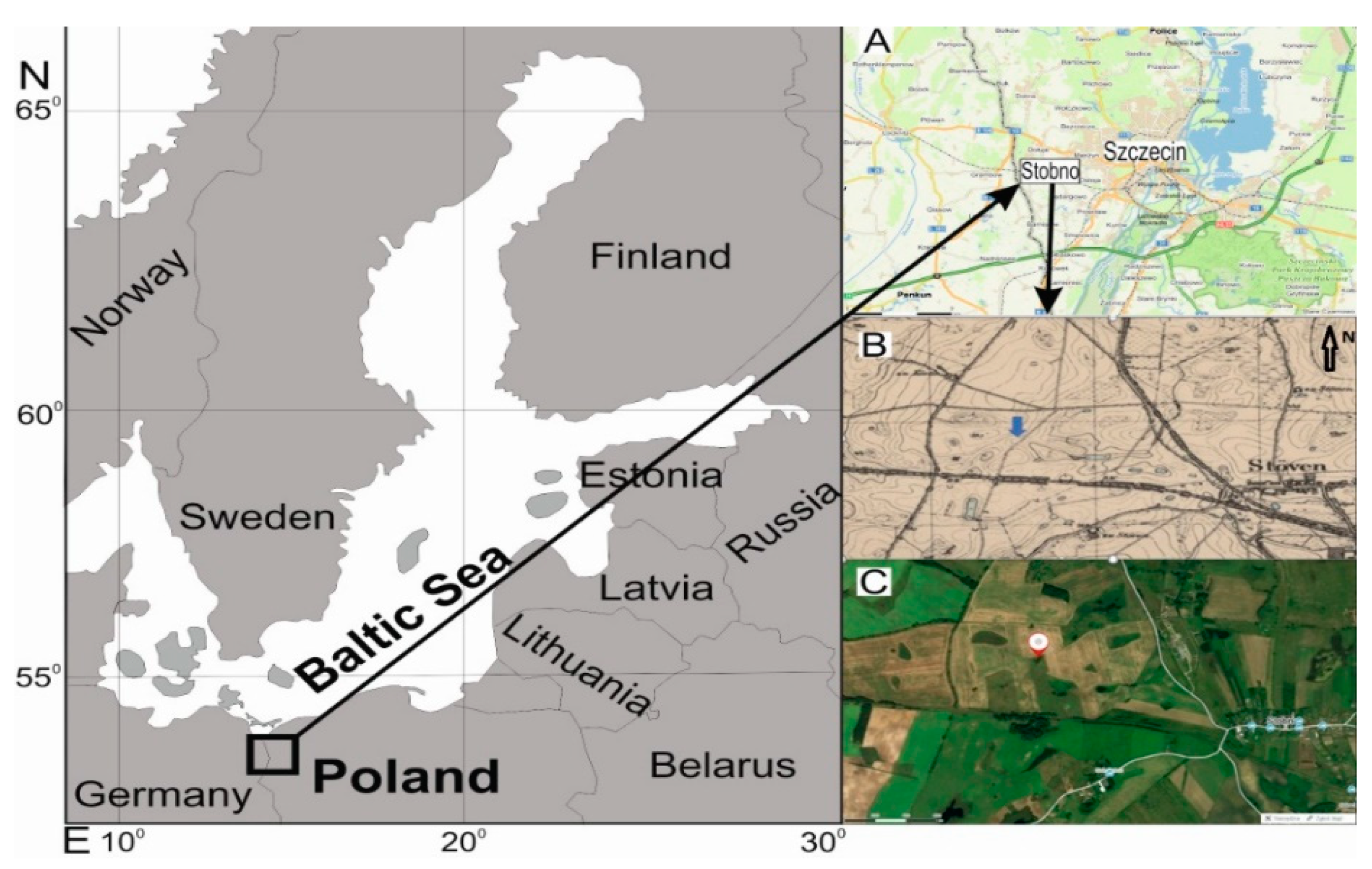
2.1. Study Area
2.2. Tree-Ring Data
2.3. Climate
3. Results
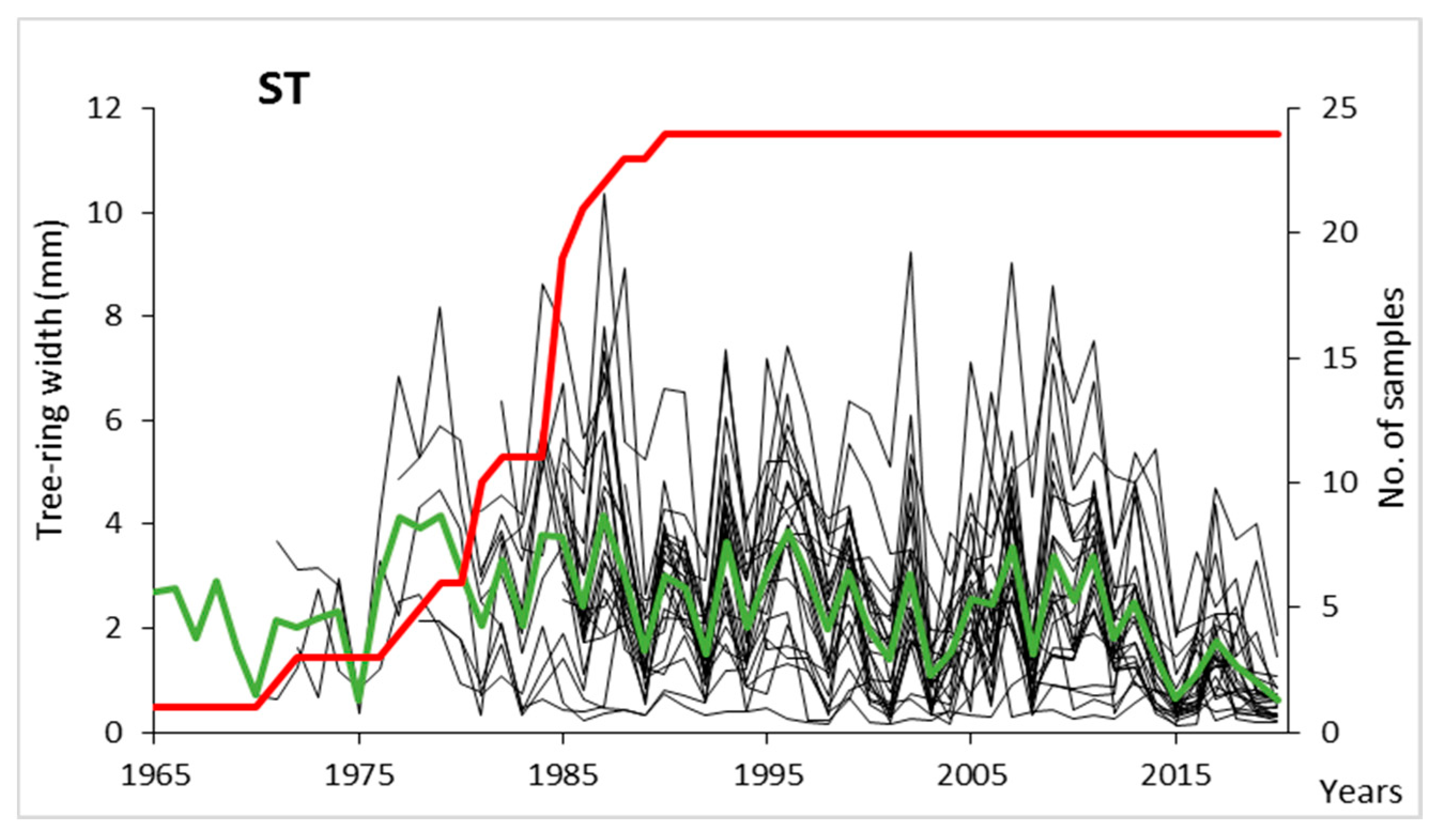
3.1. Ring-Width Chronology
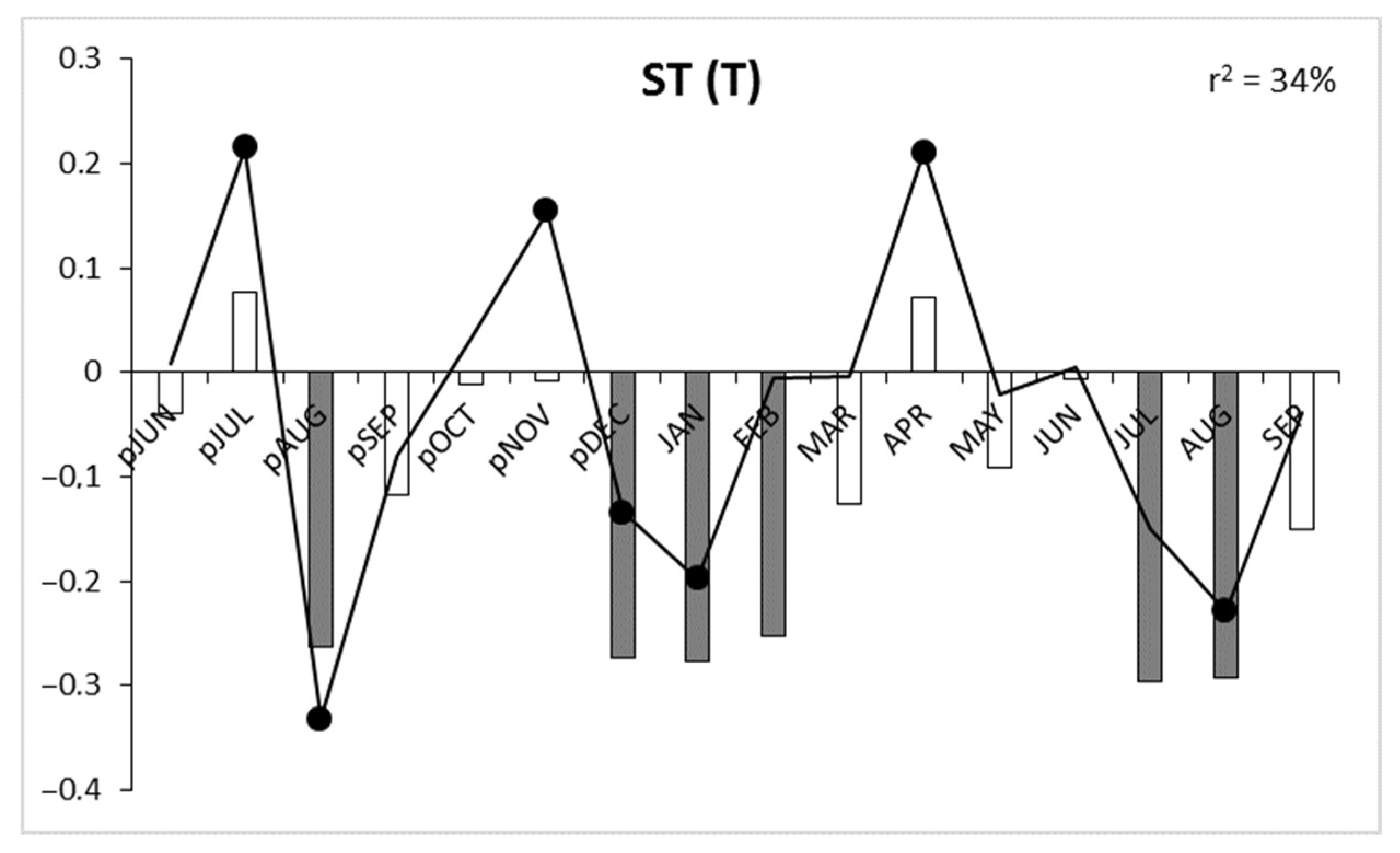
3.2. Correlation and Response Function
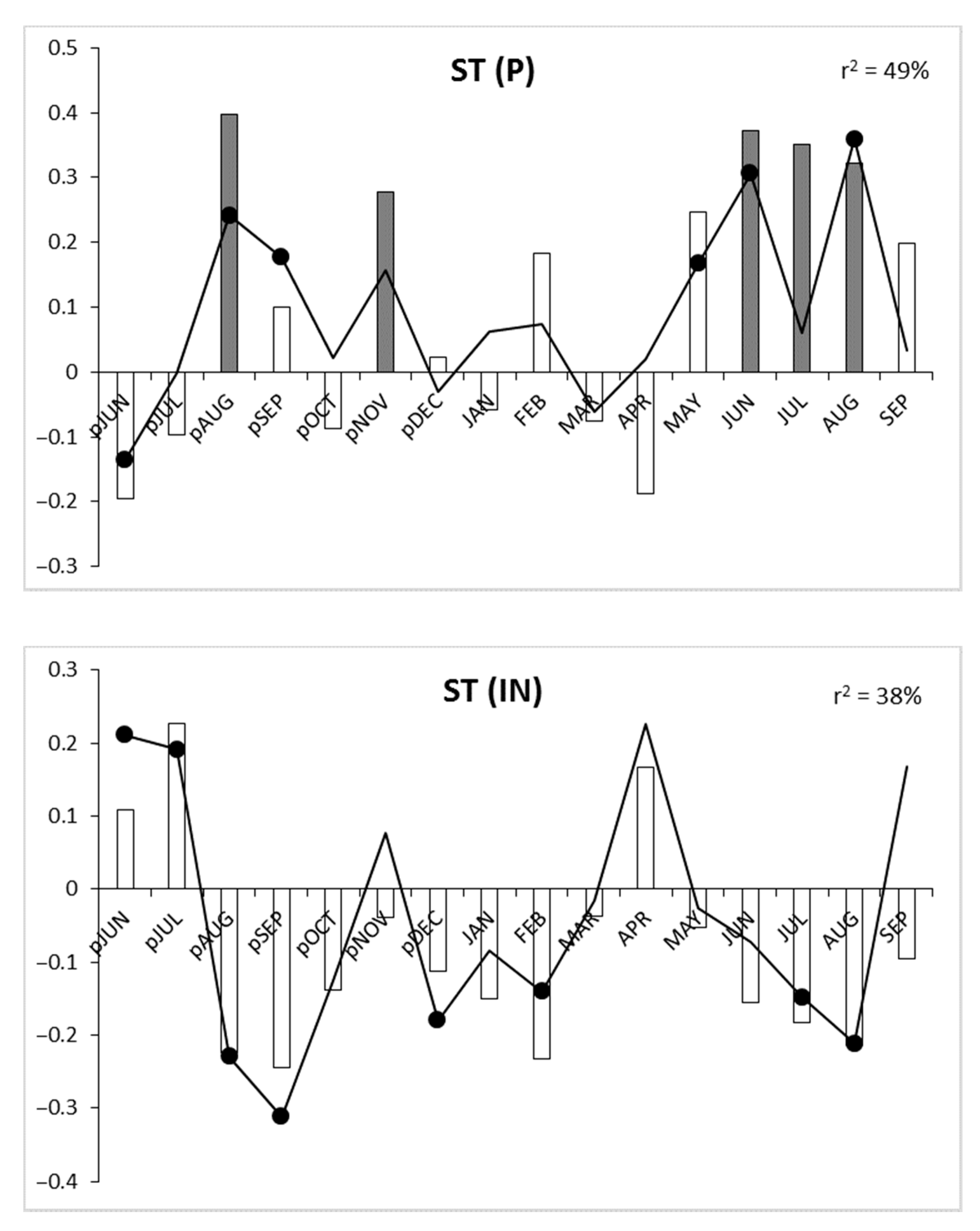
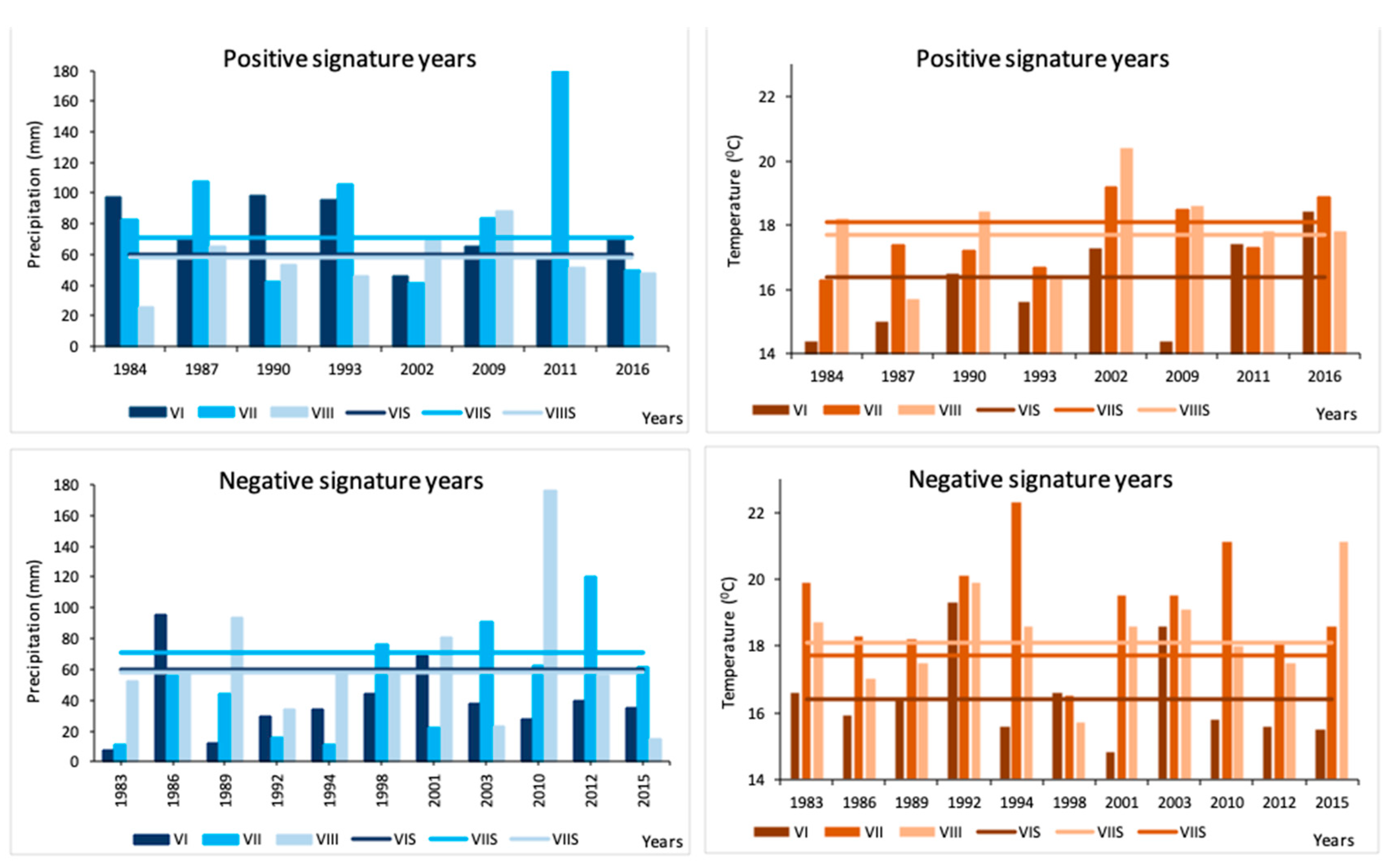
3.3. Signature Years
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guitian, J.; Fuentes, M. Reproductive biology of Crataegus monogyna in northwestern Spain. Acta Ecol. 1992, 13, 3–11. [Google Scholar]
- Oklejewicz, K.; Chwastek, E.; Szewczyk, B.; Mitka, J. Chorological Aspects of the Occurrence of Hawthorn in the Polish Carpathians; Wydawnictwo Uniwersytetu Rzeszowskiego: Rzeszów, Poland, 2014; pp. 1–210. [Google Scholar]
- Hassler, M. World Plants: Synonymic Checklists of the Vascular Plants of the World (version Nov 2018). In 2019, Species 2000 & ITIS Catalogue of Life; Roskov, Y., Ower, G., Orrell, T., Nicolson, D., Bailly, N., Kirk, P.M., Bourgoin, T., DeWalt, R.E., Decock, W., Nieukerken, E., et al., Eds.; Naturalis: Leiden, The Netherlands, 2019; ISSN 2405-884X. Annual Checklist; Species 2000; Available online: www.catalogueoflife.org/annual-checklist/2019 (accessed on 9 November 2020).
- Fitzgerald, H.; Helpdesk, G.N. Nordic Crop Wild Relative (CWR) Checklist. Version 1.16. Nordic Genetic Resource Center (NORDGEN). 2020. Available online: https://www.gbif.org/dataset/8027d8d5-c8bc-4d54-bee9-f854f141b442 (accessed on 9 November 2020).
- Sobral, M.; Larrinaga, A.R.; Guitián, J. Do seed-dispersing birds exert selection on optimal plant trait combinations? Correlated phenotypic selection on the fruit and seed size of hawthorn (Crataegus monogyna). Evol. Ecol. 2010, 24, 1277–1290. [Google Scholar] [CrossRef]
- Annotated Bibliography on the Ecology and Management of Invasive Species: Common hawthorn (Crataegus monogyna). Prepared by Judith Cullington & Associates, Victoria, BC for the Garry Oak Ecosystems Recovery Team and the Nature Conservancy of Canada, March 2002. Available online: https://stewardshipcentrebc.ca/PDF_docs/GOERT/Publications/Inv_Bibliographies/Bib_cratmono.pdf (accessed on 9 November 2020).
- Williams, P.A.; Buxton, R.P. Hawthorn (Crataegus monogyna) Populations in Mid-Canterbury. N. Z. J. Ecol. 1986, 9, 11–17. [Google Scholar]
- Sallabanks, R. Fruit fate, frugivory, and fruit characteristics: A study of the hawthorn, Crataegus monogyna (Rosaceae). Oecologia 1992, 91, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Sallabanks, R. Fruiting plant attractiveness to avian seed dispersers: Native vs. Invasive Crataegus in western Oregon. Madrono 1993, 40, 108–116. [Google Scholar]
- Phipps, J.B. Introduction to the red-fruited hawthorns (Crataegus, Rosaceae) of Western North America. Can. J. Bot. 1998, 76, 1863–1899. [Google Scholar]
- Gostyńska-Jakuszewska, M.; Hrabetova-Uhrova, A. Distribution of Crataegus in Poland and Czechoslovakia. Preslia Praha 1983, 55, 9–24. [Google Scholar]
- Seneta, W. Drzewa i krzewy liściaste. Tom II; Wydawnictwo Naukowe PWN: Warszawa, Poland, 1994. [Google Scholar]
- Seneta, W.; Dolatowski, J. Dendrology; PWN: Warszawa, Poland, 2004; pp. 1–559. [Google Scholar]
- Özcan, M.; Hacıseferoğulları, H.; Marakoğlu, T.; Arslan, D. Hawthorn (Crataegus spp.) fruit: Some physical and chemical properties. J. Food Eng. 2005, 69, 409–413. [Google Scholar] [CrossRef]
- Kumar, D.; Arya, V.; Bhat, Z.A.; Khan, N.A.; Prasad, D.N. The genus Crataegus: Chemical and pharmacological perspectives. Rev. Bras. Farm. 2012, 22, 1187–1200. [Google Scholar] [CrossRef]
- Kulczyński, B.; Gramza-Michałowska, A. Health-promoting potential of hawthorn fruits and flowers. Probl. Hig. Epidemiol. 2016, 97, 24–28. [Google Scholar]
- Nazhand, A.; Lucarini, M.; Durazzo, A.; Zaccardelli, M.; Cristarella, S.; Souto, S.B.; Silva, A.M.; Severino, P.; Souto, E.B.; Santini, A. Hawthorn (Crataegus spp.): An Updated Overview on Its Beneficial Properties. Forests 2020, 11, 564. [Google Scholar] [CrossRef]
- Good, J.E.G.; Bryant, R.; Carlill, P. Distribution, Longevity and Survival of Upland Hawthorn (Crataegus monogyna) Scrub in North Wales in Relation to Sheep Grazing. J. Appl. Ecol. 1990, 27, 272. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. An updated list of species used in tree-ring research. Tree Ring Bull. 1993, 53, 17–43. [Google Scholar]
- Schweingruber, F.H. Microscopic Wood Anatomy. Swiss Federal Institute for Forest, Snow and Landscape Research; WSL: Birmensdorf, Switzerland, 1990; pp. 1–226. [Google Scholar]
- Kondracki, J. Geografia Fizyczna Polski; PWN: Warszawa, Poland, 1988; pp. 1–464. [Google Scholar]
- Abadi, H.S.N.; Soltani, A.; Saeidi, Z.; Zadeh, S.G. Study of Spatial Distribution of the Hawthorn (Crataegus monogyna) Trees Attacked by Orchard Ermine (Yponomeuta padella) in Bazoft Forests of Chaharmahal and Bakhtiari Province. Iran. J. Appl. Ecol. 2016, 4, 39–49. [Google Scholar] [CrossRef]
- Taggar, G.K.; Arora, R. Insect Biotypes and Host Plant Resistance. In Breeding Insect Resistant Crops for Sustainable Agriculture; Arora, R., Sandhu, S., Eds.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Mindur, B. Dendrometer 1,0. 2000. Available online: www.ictinternational.com/support/software/ (accessed on 9 November 2020).
- Holmes, R.J. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating Crossdating accuracy: A manual and tutorial for the compuret program COFECHA. Tree Ring Res. 2001, 57, 205–221. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Guide for Computer Program ARSTAN. In The International Tree-Ring Data Bank Program Library Version 2.0 User’s Manual; Grissino-Mayer, H.D., Holmes, R.L., Fritts, H.C., Eds.; Laboratory of Tree-Ring Research: Tuscon, AZ, USA, 1996; pp. 75–87. [Google Scholar]
- Holmes, R.J. Dendrochronology Program Library. Users Manual. 1994. Available online: https://www.ltrr.arizona.edu/software.html (accessed on 9 November 2020).
- Fritts, H.C. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976; pp. 1–582. [Google Scholar]
- Cook, E.R.; Kairiukstis, A. Methods of Dendrochronology; Kluwer Academic Publishers: New York, NY, USA, 1992; pp. 1–394. ISBN 0-7923-0586-8. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Walanus, A. Instrukcja Obsługi Programu TCS. Program TCS do Obliczania Lat Wskaźnikowych. 2002. Available online: https://www.tcs.com/careers/ace-program/en-us (accessed on 9 November 2020).
- Meyer, F.D. Pointer years analysis in dendrochronology: A comparison of methods. Dendrochronologia 1997, 16–17, 193–204. [Google Scholar]
- Jetschke, G.; Van Der Maaten, E.; Van Der Maaten-Theunissen, M. Towards the extremes: A critical analysis of pointer year detection methods. Dendrochronologia 2019, 53, 55–62. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree rings. In Basics and Applications of Dendrochronology; Kluwer Academic Publishers: New York, NY, USA, 1989; pp. 1–276. ISBN 978-0-7923-0559-0. [Google Scholar]
- Kaennel, M.; Schweingruber, F.H. Multilingual Glossary of Dendrochronology; WSL FNP: Haupt, Germany, 1990; pp. 1–467. ISBN 3-258-05259-X. [Google Scholar]
- Krawczyk, A. Program Komputerowy Tree Rings. 1995. Available online: https://www.ldeo.columbia.edu/tree-ring-laboratory/resources/software (accessed on 9 November 2020).
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Updated world map of the Köppen-Geiger climateclassification. Hydrology and Earth System Sciences Discussions. Eur. Geosci. Union 2007, 4, 439–473. [Google Scholar]
- Cedro, A. Dendrochronologiczna analiza drzew rosnących w Puszczy Bukowej. W: (Red. Grażyna Domian i Krzysztof Ziarnek) Księga Puszczy Bukowej. In Tom I: Środowisko Przyrodnicze; PPH Zapol Dmochowski, Sobczyk sp.j.; Regionalna Dyrekcja Ochrony Środowiska w Szczecinie: Szczecin, Poland, 2010; pp. 122–128. [Google Scholar]
- Cedro, A. Dendrochronology of Yew in Poland and Western Ukraine; Zapol: Szczecin, Poland, 2012; pp. 1–230. ISBN 978-83-7518-386-3. [Google Scholar]
- Cedro, A. Dendrochronological Analysis of the Wild Service Tree (Sorbus torminalis L.) in Poland; Zapol: Szczecin, Poland, 2016; pp. 1–210. ISBN 978-83-7518-815-8. [Google Scholar]
- Koprowski, M.; Zielski, A. Dendrochronology of Norway spruce (Picea abies (L.) Karst.) from two range centres in lowland Poland. Trees 2006, 20, 383–390. [Google Scholar] [CrossRef]
- Koprowski, M. Dendrochronological analysis of European beech (Fagus sylvatica L.) tree rings in the Iława Forest District. Sylwan 2006, 5, 44–50. Available online: https://www.researchgate.net/publication/257046958_Dendrochronological_analysis_of_European_beech_Fagus_sylvatica_L_tree_rings_in_the_Ilawa_Forest_District (accessed on 9 November 2020).
- Friedrichs, D.A.; Büntgen, U.; Frank, D.C.; Esper, J.; Neuwirth, B.; Löffler, J. Complex climate controls on 20th century oak growth in Central-West Germany. Tree Physiol. 2009, 29, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misi, D.; Puchałka, R.; Pearson, C.; Robertson, I.; Koprowski, M. Differences in the Climate-Growth Relationship of Scots Pine: A Case Study from Poland and Hungary. Forests 2019, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Scharnweber, T.; Heinze, L.; Cruz-García, R.; Van Der Maaten-Theunissen, M.; Wilmking, M. Confessions of solitary oaks: We grow fast but we fear the drought. Dendrochronologia 2019, 55, 43–49. [Google Scholar] [CrossRef]
- Dell’Oro, M.; Mataruga, M.; Sass-Klaassen, U.; Fonti, P. Climate change threatens on endangered relict Serbian spruce. Dendrochronologia 2020, 59, 125651. [Google Scholar] [CrossRef]
- Cedro, A. Climatic changes in Western Pomerania in the Light of Analysis of Tree-Ring Sequences of Scots Pine, Douglas Fir, and Native Species of Oak; Science Research: Szczecin, Poland, 2004; pp. 1–149. ISBN 83-89402-03-3. [Google Scholar]
- Buras, A.; Sass-Klaassen, U.; Verbeek, I.; Copini, P. Provenance selection and site conditions determine growth performance of pedunculate oak. Dendrochronologia 2020, 61, 125705. [Google Scholar] [CrossRef]
- Mijnsbrugge, K.V.; Janssens, A. Differentiation and Non-Linear Responses in Temporal Phenotypic Plasticity of Seasonal Phenophases in a Common Garden of Crataegus monogyna Jacq. Forests 2019, 10, 293. [Google Scholar] [CrossRef] [Green Version]
- Beloiu, M.B.; Stahlmann, R.S.; Beierkuhnlein, C.B. High Recovery of Saplings after Severe Drought in Temperate Deciduous Forests. Forests 2020, 11, 546. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
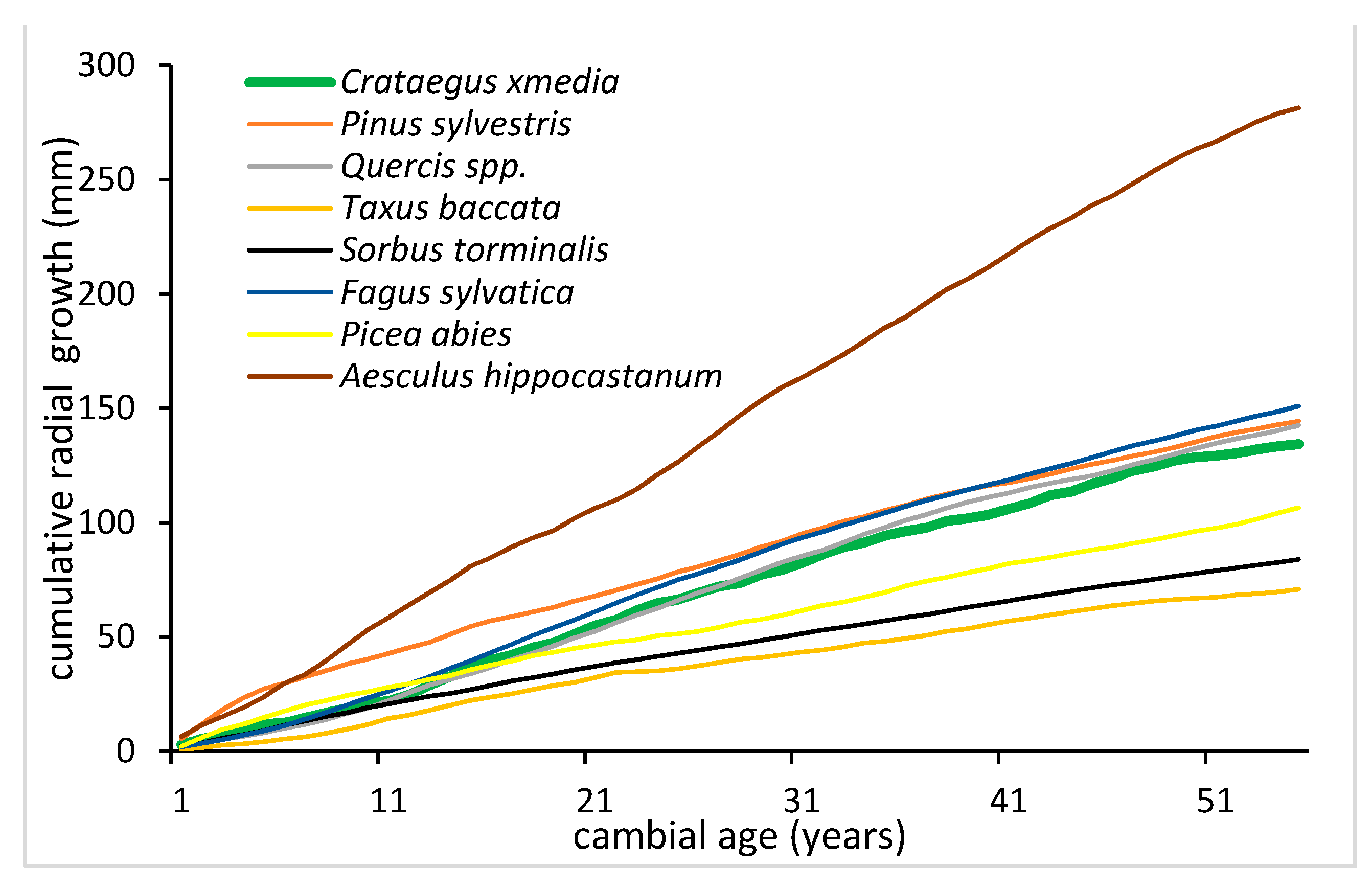{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pinus sylvestris L. | Pinus nigra J.F. Arnold | Picea abies (L.)H.Karst | Taxus baccata L. | Pseudotsuga menziesii Franco | Quercus spp. | Quercus rubra L. | Fagus sylvatica L. | Aesculus hippocastanum L. | Sorbus torminalis L. | Platanus × hispanica | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| t | 3.21 | 2.77 | 2.99 | 2.79 | 2.32 | 3.63 | 3.49 | 2.92 | 3.08 | 2.34 | 2.78 |
| Gl | 60% | 59% | 58% | 62% | 53% | 68% | 66% | 60% | 65% | 50% | 58% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cedro, A.; Cedro, B. Tree-Ring Analysis of Intermediate Hawthorn (Crataegus media Bechst.) in NW Poland. Forests 2021, 12, 29. https://doi.org/10.3390/f12010029
Cedro A, Cedro B. Tree-Ring Analysis of Intermediate Hawthorn (Crataegus media Bechst.) in NW Poland. Forests. 2021; 12(1):29. https://doi.org/10.3390/f12010029
Chicago/Turabian StyleCedro, Anna, and Bernard Cedro. 2021. "Tree-Ring Analysis of Intermediate Hawthorn (Crataegus media Bechst.) in NW Poland" Forests 12, no. 1: 29. https://doi.org/10.3390/f12010029
APA StyleCedro, A., & Cedro, B. (2021). Tree-Ring Analysis of Intermediate Hawthorn (Crataegus media Bechst.) in NW Poland. Forests, 12(1), 29. https://doi.org/10.3390/f12010029