Estimating Needle and Shoot Inclination Angle Distributions and Projection Functions in Five Larix principis-rupprechtii Plots via Leveled Digital Camera Photography

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plots Description
2.2. Data Acquisition
2.3. Data Processing
2.3.1. Needle and Shoot Inclination Angle Estimation
2.3.2. Fitting the Needle and Shoot Inclination Angle Measurements
2.3.3. Needle and Shoot Projection Function Calculation
3. Results and Discussion
3.1. Comparison of Manual and Quasi-Automatic Methods Used to Derive the Needle Inclination Angle Measurements
3.2. Comparison of Beta and Ellipsoidal Distribution Functions for Fitting the Shoot or Needle Inclination Angle Measurements
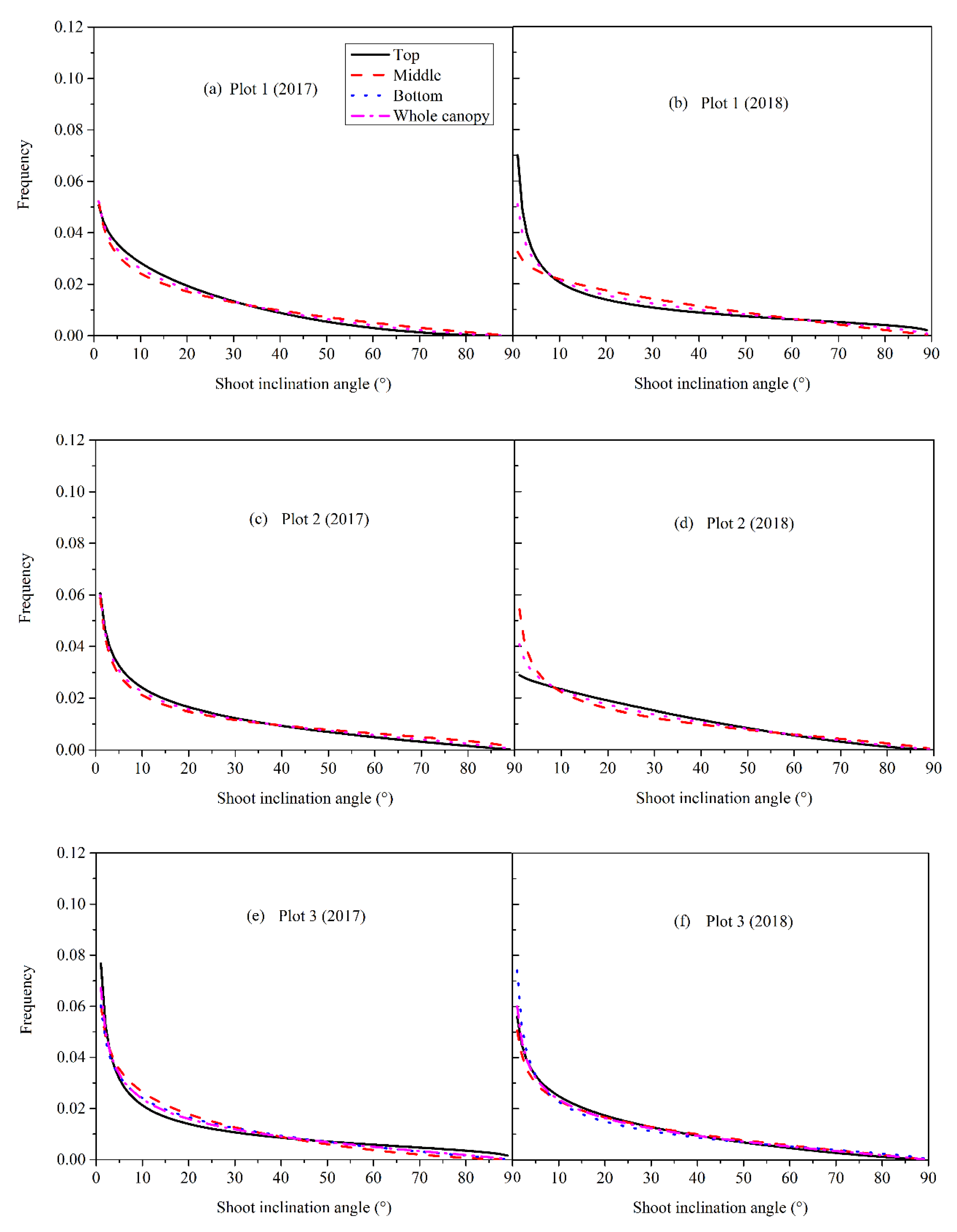
3.3. Shoot Angle Distribution
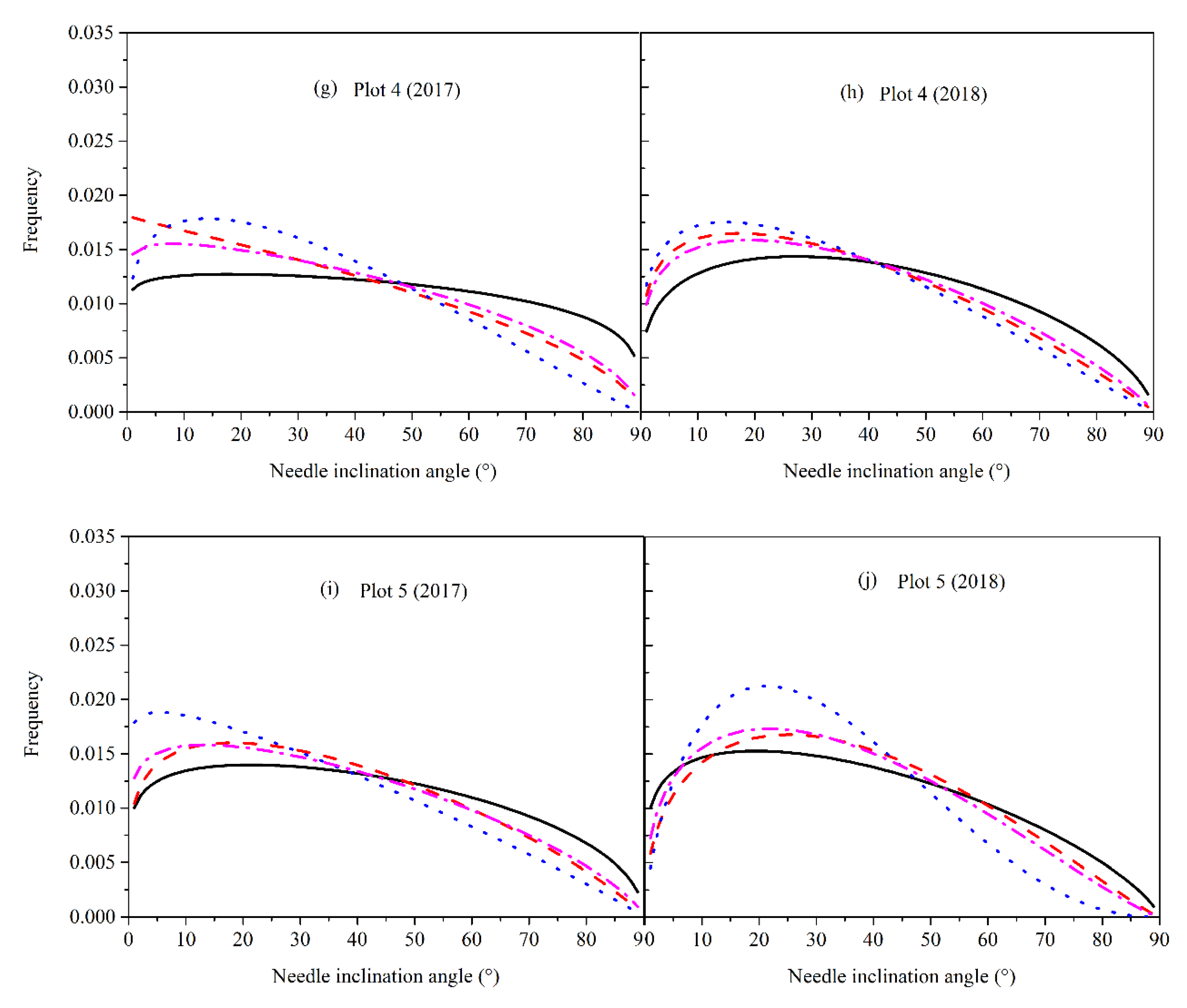
3.4. Needle Angle Distribution
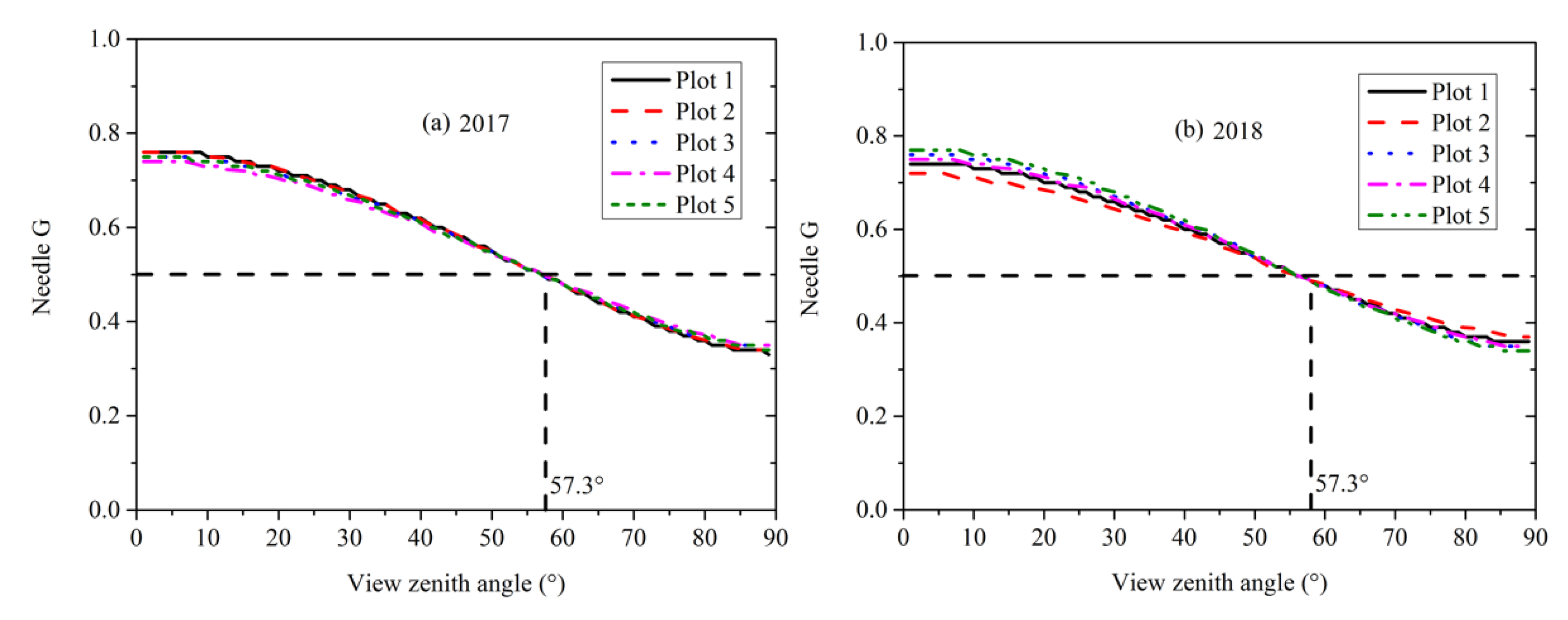
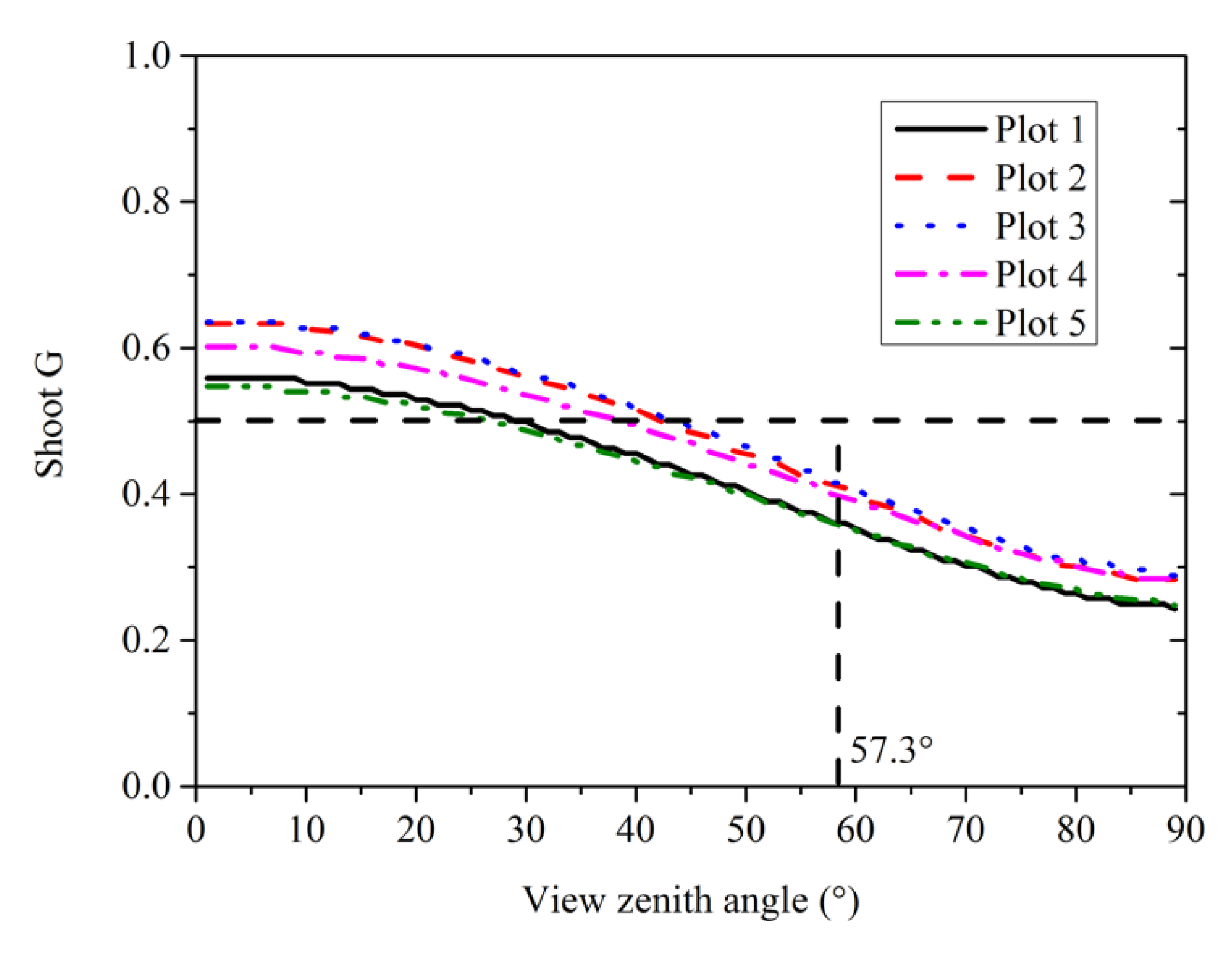
3.5. Needle and Shoot Projection Functions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zou, X.; Mõttus, M.; Tammeorg, P.; Torres, C.L.; Takala, T.; Pisek, J.; Mäkelä, P.; Stoddard, F.L.; Pellikka, P. Photographic measurement of leaf angles in field crops. Agric. For. Meteorol. 2014, 184, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Ross, J. The Radiation Regime and Architecture of Plant Stands; Dr. W. Junk Publ.: The Hague, The Netherlands, 1981. [Google Scholar]
- Chianucci, F.; Pisek, J.; Raabe, K.; Marchino, L.; Ferrara, C.; Corona, P. A dataset of leaf inclination angles for temperate and boreal broadleaf woody species. Ann. For. Sci. 2018, 75, 50. [Google Scholar] [CrossRef] [Green Version]
- Norman, J.M.; Campbell, G.S. Canopy structure. In Plant Physiological Ecology; Pearcy, R.W., EEhleringer, J.R., Mooney, H.A., RRundel, P.W., Eds.; Springer: Dordrecht, The Netherlands, 1989; pp. 301–325. [Google Scholar]
- Lang, A.R.G. Leaf orientation of a cotton plant. Agric. Meteorol. 1973, 11, 37–51. [Google Scholar] [CrossRef]
- Sinoquet, H.; Rivet, P. Measurement and visualization of the architecture of an adult tree based on a three-dimensional digitising device. Trees 1997, 11, 265–270. [Google Scholar] [CrossRef]
- Sonohat, G.; Sinoquet, H.; Kulandaivelu, V.; Combes, D.; Lescourret, F. Three-dimensional reconstruction of partially 3D-digitized peach tree canopies. Tree Physiol. 2006, 26, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Kucharik, C.J.; Norman, J.M.; Gower, S.T. Measurements of leaf orientation, light distribution and sunlit leaf area in a boreal aspen forest. Agric. For. Meteorol. 1998, 91, 127–148. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A.; Adams, R.S. Evaluation of hemispherical photography for determining plant area index and geometry of a forest stand. Agric. For. Meteorol. 1991, 56, 129–143. [Google Scholar] [CrossRef]
- Wagner, S.; Hagemeier, M. Method of segmentation affects leaf inclination angle estimation in hemispherical photography. Agric. For. Meteorol. 2006, 139, 12–24. [Google Scholar] [CrossRef]
- Macfarlane, C.; Arndt, S.K.; Livesley, S.J.; Edgar, A.C.; White, D.A.; Adams, M.A.; Eamus, D. Estimation of leaf area index in eucalypt forest with vertical foliage, using cover and fullframe fisheye photography. For. Ecol. Manag. 2007, 242, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, C.; Hoffman, M.; Eamus, D.; Kerp, N.; Higginson, S.; McMurtrie, R.; Adams, M. Estimation of leaf area index in eucalypt forest using digital photography. Agric. For. Meteorol. 2007, 143, 176–188. [Google Scholar] [CrossRef]
- Ryu, Y.; Sonnentag, O.; Nilson, T.; Vargas, R.; Kobayashi, H.; Wenk, R.; Baldocchi, D.D. How to quantify tree leaf area index in an open savanna ecosystem: A multi-instrument and multi-model approach. Agric. For. Meteorol. 2010, 150, 63–76. [Google Scholar] [CrossRef]
- Raabe, K.; Pisek, J.; Sonnentag, O.; Annuk, K. Variations of leaf inclination angle distribution with height over the growing season and light exposure for eight broadleaf tree species. Agric. For. Meteorol. 2015, 214–215, 2–11. [Google Scholar] [CrossRef]
- McNeil, B.E.; Pisek, J.; Lepisk, H.; Flamenco, E.A. Measuring leaf angle distribution in broadleaf canopies using UAVs. Agric. For. Meteorol. 2016, 218–219, 204–208. [Google Scholar] [CrossRef]
- Qi, J.; Xie, D.; Li, L.; Zhang, W.; Mu, X.; Yan, G. Estimating Leaf Angle Distribution From Smartphone Photographs. IEEE Geosci. Remote Sens. Lett. 2019, 16, 1190–1194. [Google Scholar] [CrossRef]
- Zheng, G.; Moskal, L.M. Leaf Orientation Retrieval From Terrestrial Laser Scanning (TLS) Data. Geosci. Remote Sens. IEEE Trans. 2012, 50, 3970–3979. [Google Scholar] [CrossRef]
- Hosoi, F.; Omasa, K. Factors contributing to accuracy in the estimation of the woody canopy leaf area density profile using 3D portable lidar imaging. J. Exp. Bot. 2007, 58, 3463–3473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoi, F.; Omasa, K. Estimating leaf inclination angle distribution of broad-leaved trees in each part of the canopies by a high-resolution portable scanning lidar. J. Agric. Meteorol. 2015, 71, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Bailey, B.N.; Mahaffee, W.F. Rapid measurement of the three-dimensional distribution of leaf orientation and the leaf angle probability density function using terrestrial LiDAR scanning. Remote Sens. Environ. 2017, 194, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Vicari, M.B.; Pisek, J.; Disney, M. New estimates of leaf angle distribution from terrestrial LiDAR: Comparison with measured and modelled estimates from nine broadleaf tree species. Agric. For. Meteorol. 2019, 264, 322–333. [Google Scholar] [CrossRef]
- Liu, J.; Skidmore, A.K.; Wang, T.; Zhu, X.; Premier, J.; Heurich, M.; Beudert, B.; Jones, S. Variation of leaf angle distribution quantified by terrestrial LiDAR in natural European beech forest. ISPRS J. Photogramm. Remote Sens. 2019, 148, 208–220. [Google Scholar] [CrossRef]
- Itakura, K.; Hosoi, F. Estimation of Leaf Inclination Angle in Three-Dimensional Plant Images Obtained from Lidar. Remote Sens. 2019, 11, 344. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Cao, L.; Xue, L.; Chen, B.; An, F.; Yun, T. Extraction of Leaf Biophysical Attributes Based on a Computer Graphic-based Algorithm Using Terrestrial Laser Scanning Data. Remote Sens. 2018, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, T.; Skidmore, A.K.; Jones, S.; Heurich, M.; Beudert, B.; Premier, J. Comparison of terrestrial LiDAR and digital hemispherical photography for estimating leaf angle distribution in European broadleaf beech forests. ISPRS J. Photogramm. Remote Sens. 2019, 158, 76–89. [Google Scholar] [CrossRef]
- Leblanc, S.G.; Chen, J.M.; Fernandes, R.; Deering, D.W.; Conley, A. Methodology comparison for canopy structure parameters extraction from digital hemispherical photography in boreal forests. Agric. For. Meteorol. 2005, 129, 187–207. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zheng, G.; Eitel, J.U.H.; Magney, T.S.; Moskal, L.M. Retrieving forest canopy extinction coefficient from terrestrial and airborne lidar. Agric. For. Meteorol. 2017, 236, 1–21. [Google Scholar] [CrossRef]
- Pisek, J.; Ryu, Y.; Alikas, K. Estimating leaf inclination and G-function from leveled digital camera photography in broadleaf canopies. Trees Struct. Funct. 2011, 25, 919–924. [Google Scholar] [CrossRef]
- Pisek, J.; Sonnentag, O.; Richardson, A.D.; Mõttus, M. Is the spherical leaf inclination angle distribution a valid assumption for temperate and boreal broadleaf tree species? Agric. For. Meteorol. 2013, 169, 186–194. [Google Scholar] [CrossRef]
- Utsugi, H.; Araki, M.; Kawasaki, T.; Ishizuka, M. Vertical distributions of leaf area and inclination angle, and their relationship in a 46-year-old Chamaecyparis obtusa stand. For. Ecol. Manag. 2006, 225, 104–112. [Google Scholar] [CrossRef]
- Zou, J.; Leng, P.; Hou, W.; Zhong, P.; Chen, L.; Mai, C.; Qian, Y.; Zuo, Y. Evaluating Two Optical Methods of Woody-to-Total Area Ratio with Destructive Measurements at Five Larix gmelinii Rupr. Forest Plots in China. Forests 2018, 9, 746. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Zuo, Y.; Zhong, P.; Hou, W.; Leng, P.; Chen, B. Performance of Four Optical Methods in Estimating Leaf Area Index at Elementary Sampling Unit of Larix principis-rupprechtii Forests. Forests 2019, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Kimes, D.S.; Kirchner, J.A. Diurnal variations of vegetation canopy structure. Int. J. Remote Sens. 1983, 4, 257–271. [Google Scholar] [CrossRef]
- Zou, J.; Hou, W.; Chen, L.; Wang, Q.; Zhong, P.; Zuo, Y.; Luo, S.; Leng, P. Evaluating the impact of sampling schemes on leaf area index measurements from digital hemispherical photography in Larix gmeliniiLarix principis-rupprechtii Rupr. forest plots. For. Ecosyst. 2020, 7, 52. [Google Scholar] [CrossRef]
- Gioi, R.G.v.; Jakubowicz, J.; Morel, J.; Randall, G. LSD: A Fast Line Segment Detector with a False Detection Control. IEEE Trans. Pattern Anal. Mach. Intell. 2010, 32, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Gioi, R.G.v.; Jakubowicz, J.; Morel, J.-M.; Randall, G. LSD: A Line Segment Detector. Image Process. Line 2012, 2, 35–55. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Wang, C.; Cheng, J.; Chen, B.; Jia, F.; Chen, Z.; Li, J. Line segment extraction for large scale unorganized point clouds. ISPRS J. Photogramm. Remote Sens. 2015, 102, 172–183. [Google Scholar] [CrossRef]
- Cho, N.; Yuille, A.; Lee, S. A Novel Linelet-Based Representation for Line Segment Detection. IEEE Trans. Pattern Anal. Mach. Intell. 2018, 40, 1195–1208. [Google Scholar] [CrossRef]
- Hofer, M.; Maurer, M.; Bischof, H. Efficient 3D scene abstraction using line segments. Comput. Vis. Image Underst. 2017, 157, 167–178. [Google Scholar] [CrossRef]
- Tang, G.; Xiao, Z.; Liu, Q.; Liu, H. A Novel Airport Detection Method via Line Segment Classification and Texture Classification. IEEE Geosci. Remote Sens. Lett. 2015, 12, 2408–2412. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, L.; Huang, S.; Yan, L.; Dissanayake, G. Line matching based on planar homography for stereo aerial images. ISPRS J. Photogramm. Remote Sens. 2015, 104, 1–17. [Google Scholar] [CrossRef] [Green Version]
- W.H. Freeman and Company. Biometry: Principles and Practice of Statistics in Biological Research, 2nd ed.; W.H. Freeman and Company: San Francisco, CA, USA, 1981. [Google Scholar]
- Wang, W.M.; Li, Z.L.; Su, H.B. Comparison of leaf angle distribution functions: Effects on extinction coefficient and fraction of sunlit foliage. Agric. For. Meteorol. 2007, 143, 106–122. [Google Scholar] [CrossRef]
- Pisek, J.; Lang, M.; Nilson, T.; Korhonen, L.; Karu, H. Comparison of methods for measuring gap size distribution and canopy nonrandomness at Järvselja RAMI (RAdiation transfer Model Intercomparison) test sites. Agric. For. Meteorol. 2011, 151, 365–377. [Google Scholar] [CrossRef]
- Campbell, G.S. Derivation of an angle density function for canopies with ellipsoidal leaf angle distributions. Agric. For. Meteorol. 1990, 49, 173–176. [Google Scholar] [CrossRef]
- Ross, J. Radiative transfer in plant communities. In Vegetation and the Atmosphere; Monteith, J.L., Ed.; Academic Press: London, UK, 1975; Volume 1, pp. 13–55. [Google Scholar]
- De Wit, C.T. Photosynthesis of Leaf Canopies; Wageningen University: Wageningen, The Netherlands, 1965. [Google Scholar]
- Lemeur, R.; Blad, B.L. A critical review of light models for estimating the shortwave radiation regime of plant canopies. Agric. Meteorol. 1974, 14, 255–286. [Google Scholar] [CrossRef]
- Chen, J.M.; Rich, P.M.; Gower, S.T.; Norman, J.M.; Plummer, S. Leaf area index of boreal forests: Theory, techniques and measurements. J. Geophys. Res. 1997, 102, 29429–29443. [Google Scholar] [CrossRef]
- Oker-Blom, P.; Kellomäki, S. Effect of angular distribution of foliage on light absorption and photosynthesis in the plant canopy: Theoretical computations. Agric. Meteorol. 1982, 26, 105–116. [Google Scholar] [CrossRef]
- Kull, O.; Broadmeadow, M.; Kruijt, B.; Meir, P. Light distribution and foliage structure in an oak canopy. Trees 1999, 14, 55–64. [Google Scholar] [CrossRef]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- King, D.A. The Functional Significance of Leaf Angle in Eucalyptus. Aust. J. Bot. 1997, 45, 619–639. [Google Scholar] [CrossRef]
- Cescatti, A.; Alessandro, U. Leaf to Landscape. In Photosynthetic Adaptation: Chloroplast to Landscape; Smith, W.K., Vogelmann, T.C., Critchley, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 42–85. [Google Scholar]
- Weiss, M.; Baret, F.; Smith, G.J.; Jonckheere, I.; Coppin, P. Review of methods for in situ leaf area index (LAI) determination: Part II. Estimation of LAI, errors and sampling. Agric. For. Meteorol. 2004, 121, 37–53. [Google Scholar] [CrossRef]
- Yan, G.; Hu, R.; Luo, J.; Weiss, M.; Jiang, H.; Mu, X.; Xie, D.; Zhang, W. Review of indirect optical measurements of leaf area index: Recent advances, challenges, and perspectives. Agric. For. Meteorol. 2019, 265, 390–411. [Google Scholar] [CrossRef]
- Zou, J.; Zhuang, Y.; Chianucci, F.; Mai, C.; Lin, W.; Leng, P.; Luo, S.; Yan, B. Comparison of Seven Inversion Models for Estimating Plant and Woody Area Indices of Leaf-on and Leaf-off Forest Canopy Using Explicit 3D Forest Scenes. Remote Sens. 2018, 10, 1297. [Google Scholar] [CrossRef] [Green Version]
- Woodgate, W. In-Situ Leaf Area Index Estimate Uncertainty in Forests: Supporting Earth Observation Product Calibration and Validation. Ph.D. Thesis, RMIT University, Melbourne, Australia, 2015. [Google Scholar]
- Woodgate, W.; Disney, M.; Armston, J.D.; Jones, S.D.; Suarez, L.; Hill, M.J.; Wilkes, P.; Soto-Berelov, M.; Haywood, A.; Mellor, A. An improved theoretical model of canopy gap probability for Leaf Area Index estimation in woody ecosystems. For. Ecol. Manag. 2015, 358, 303–320. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot 1 | Plot 2 | Plot 3 | Plot 4 | Plot 5 | |
|---|---|---|---|---|---|
| Longitude and latitude | N 42°24′43″, E 117°19′4″ | N 42°24′2″, E 117°18′40″ | N 42°18′2″, E 117°18′9″ | N 42°25′22″, E 117°19′32″ | N 42°17′42″, E 117°16′53″ |
| Mean tree height (m) * | 19.4 | 20.4 | 12.6 | 13.3 | 8.7 |
| Average diameter at breast height (cm) | 26.6 | 27.2 | 12.7 | 14.1 | 9.2 |
| Stand density (stems/ha) | 464 | 384 | 2320 | 1760 | 3904 |
| Tree age (~years) | 54 | 55 | 21 | 22 | 13 |
| Needle-to-shoot area ratio () ** | 1.36 | 1.20 | 1.18 | 1.23 | 1.37 |
| Litter collection LAI *** | 4.65 | 3.58 | 4.96 | 3.04 | 6.69 |
| Tree species | Larix principis-rupprechtii | ||||
| Plot Name | Height Level | Manual | Quasi-Automatic | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | ||||||||||
| Count | Mean | SD | Count | Mean | SD | Count | Mean | SD | Count | Mean | SD | ||
| Plot 1 | Top | 192 | 34.52 | 22.18 | 146 | 40.96 | 22.0 | 192 | 35.04 | 22.30 | 146 | 41.75 | 21.67 |
| Middle | 189 | 34.5 | 22.44 | 158 | 33.11 | 20.82 | 189 | 34.70 | 22.32 | 158 | 33.43 | 20.85 | |
| Bottom | \ * | \ | \ | \ | \ | \ | \ | \ | \ | \ | \ | \ | |
| All | 381 | 34.51 | 22.28 | 304 | 36.88 | 21.72 | 381 | 34.87 | 22.28 | 304 | 37.43 | 21.62 | |
| Plot 2 | Top | 141 | 39.19 | 22.87 | 259 | 38.27 | 22.99 | 141 | 39.59 | 22.49 | 259 | 39.11 | 23.07 |
| Middle | 136 | 29.97 | 20.72 | 250 | 39.88 | 22.01 | 136 | 30.02 | 20.63 | 250 | 40.63 | 22.13 | |
| Bottom | \ | \ | \ | \ | \ | \ | \ | \ | \ | \ | \ | \ | |
| All | 277 | 34.66 | 22.28 | 509 | 39.06 | 22.51 | 277 | 34.89 | 22.08 | 509 | 39.86 | 22.60 | |
| Plot 3 | Top | 134 | 39 | 22.91 | 227 | 36.27 | 22.0 | 134 | 39.05 | 22.81 | 227 | 36.92 | 22.08 |
| Middle | 135 | 31.99 | 21.52 | 223 | 35.14 | 21.64 | 135 | 32.37 | 20.98 | 223 | 35.47 | 21.64 | |
| Bottom | 124 | 35.79 | 22.72 | 222 | 34.63 | 20.59 | 124 | 36.11 | 22.51 | 222 | 34.82 | 20.31 | |
| All | 393 | 35.58 | 22.51 | 672 | 35.36 | 21.40 | 393 | 35.68 | 22.45 | 672 | 35.75 | 21.35 | |
| Plot 4 | Top | 170 | 41.59 | 24.74 | 108 | 39.7 | 22.75 | 170 | 42.19 | 24.66 | 108 | 39.62 | 22.83 |
| Middle | 171 | 34.59 | 23.09 | 106 | 34.88 | 21.57 | 170 | 34.65 | 23.12 | 106 | 34.70 | 20.87 | |
| Bottom | 160 | 32.71 | 20.83 | 98 | 33.24 | 20.96 | 157 | 33.01 | 20.65 | 98 | 33.60 | 20.96 | |
| All | 501 | 36.37 | 23.26 | 312 | 36.04 | 21.9 | 497 | 36.71 | 23.23 | 312 | 36.05 | 21.69 | |
| Plot 5 | Top | 222 | 39.35 | 23.44 | 185 | 37.0 | 22.38 | 222 | 39.99 | 23.29 | 185 | 37.62 | 22.24 |
| Middle | 222 | 35.71 | 21.88 | 173 | 36.225 | 20.62 | 221 | 35.88 | 21.54 | 173 | 36.81 | 20.32 | |
| Bottom | 212 | 31.89 | 21.56 | 185 | 30.85 | 17.51 | 212 | 32.08 | 21.32 | 185 | 31.64 | 17.48 | |
| All | 656 | 35.71 | 22.49 | 543 | 34.66 | 20.41 | 655 | 36.04 | 22.28 | 543 | 35.32 | 20.24 | |
| Plot Name | Height Level | 2017 | 2018 | Plot Name | Height Level | 2017 | 2018 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Count | Mean | SD | Count | Mean | SD | Count | Mean | SD | Count | Mean | SD | ||||
| Plot 1 | Top | 90 | 19.68 | 16.91 | 83 | 24.73 | 24.2 | Plot 2 | Top | 92 | 22.27 | 20.47 | 96 | 25.91 | 19.46 |
| Middle | 86 | 23.06 | 20.18 | 85 | 27.31 | 21 | Middle | 88 | 25.67 | 23.4 | 90 | 24.76 | 21.97 | ||
| Bottom | \* | \ | \ | \ | \ | \ | Bottom | \ | \ | \ | \ | \ | \ | ||
| All | 176 | 21.33 | 18.6 | 168 | 26.04 | 22.6 | All | 180 | 23.93 | 21.96 | 186 | 25.35 | 20.67 | ||
| Plot 3 | Top | 94 | 23.1 | 23.58 | 92 | 21.93 | 19.6 | Plot 4 | Top | 90 | 24.28 | 22.12 | 83 | 23.81 | 19.77 |
| Middle | 94 | 20.14 | 18.37 | 97 | 24.32 | 21.08 | Middle | 94 | 25.37 | 22.08 | 84 | 21.4 | 19.41 | ||
| Bottom | 90 | 22.26 | 20.43 | 97 | 21.96 | 21.94 | Bottom | 89 | 21.34 | 19.81 | 82 | 20.46 | 19.56 | ||
| All | 278 | 21.83 | 20.87 | 286 | 22.75 | 20.87 | All | 273 | 23.7 | 21.37 | 249 | 21.9 | 19.55 | ||
| Plot 5 | Top | 138 | 25.67 | 23.17 | 84 | 21.3 | 22.88 | ||||||||
| Middle | 134 | 20.3 | 20.53 | 86 | 22.35 | 22.25 | |||||||||
| Bottom | 138 | 20.82 | 19.78 | 92 | 21.41 | 21.44 | |||||||||
| All | 410 | 22.29 | 21.3 | 262 | 21.68 | 22.09 | |||||||||
| Plot Name | Height Level | 2017 | 2018 | Plot Name | Height Level | 2017 | 2018 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D | P | D | P | D | P | D | P | ||||
| Plot 1 | Top | 0.03 | 1.0 | 0.05 | 0.99 | Plot 2 | Top | 0.04 | 1.0 | 0.03 | 0.99 |
| Middle | 0.03 | 1.0 | 0.04 | 0.99 | Middle | 0.04 | 1.0 | 0.04 | 0.95 | ||
| Bottom | \* | \ | \ | \ | Bottom | \ | \ | \ | \ | ||
| All | 0.02 | 1.0 | 0.03 | 0.99 | All | 0.03 | 1.0 | 0.03 | 0.97 | ||
| Plot 3 | Top | 0.06 | 0.94 | 0.04 | 1.0 | Plot 4 | Top | 0.04 | 1.0 | 0.04 | 1.0 |
| Middle | 0.05 | 0.98 | 0.03 | 1.0 | Middle | 0.03 | 1.0 | 0.04 | 1.0 | ||
| Bottom | 0.04 | 1.0 | 0.03 | 1.0 | Bottom | 0.05 | 0.97 | 0.04 | 1.0 | ||
| All | 0.03 | 1.0 | 0.02 | 0.99 | All | 0.02 | 1.0 | 0.03 | 1.0 | ||
| Plot 5 | Top | 0.03 | 1.0 | 0.03 | 1.0 | ||||||
| Middle | 0.04 | 0.98 | 0.04 | 1.0 | |||||||
| Bottom | 0.04 | 0.99 | 0.05 | 0.92 | |||||||
| All | 0.02 | 1.0 | 0.03 | 0.93 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Zhong, P.; Hou, W.; Zuo, Y.; Leng, P. Estimating Needle and Shoot Inclination Angle Distributions and Projection Functions in Five Larix principis-rupprechtii Plots via Leveled Digital Camera Photography. Forests 2021, 12, 30. https://doi.org/10.3390/f12010030
Zou J, Zhong P, Hou W, Zuo Y, Leng P. Estimating Needle and Shoot Inclination Angle Distributions and Projection Functions in Five Larix principis-rupprechtii Plots via Leveled Digital Camera Photography. Forests. 2021; 12(1):30. https://doi.org/10.3390/f12010030
Chicago/Turabian StyleZou, Jie, Peihong Zhong, Wei Hou, Yong Zuo, and Peng Leng. 2021. "Estimating Needle and Shoot Inclination Angle Distributions and Projection Functions in Five Larix principis-rupprechtii Plots via Leveled Digital Camera Photography" Forests 12, no. 1: 30. https://doi.org/10.3390/f12010030
APA StyleZou, J., Zhong, P., Hou, W., Zuo, Y., & Leng, P. (2021). Estimating Needle and Shoot Inclination Angle Distributions and Projection Functions in Five Larix principis-rupprechtii Plots via Leveled Digital Camera Photography. Forests, 12(1), 30. https://doi.org/10.3390/f12010030