
Bark Thickness and Heights of the Bark Transition Area of Scots Pine


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Research Design
2.3. Data Analysis and Statistics
3. Results
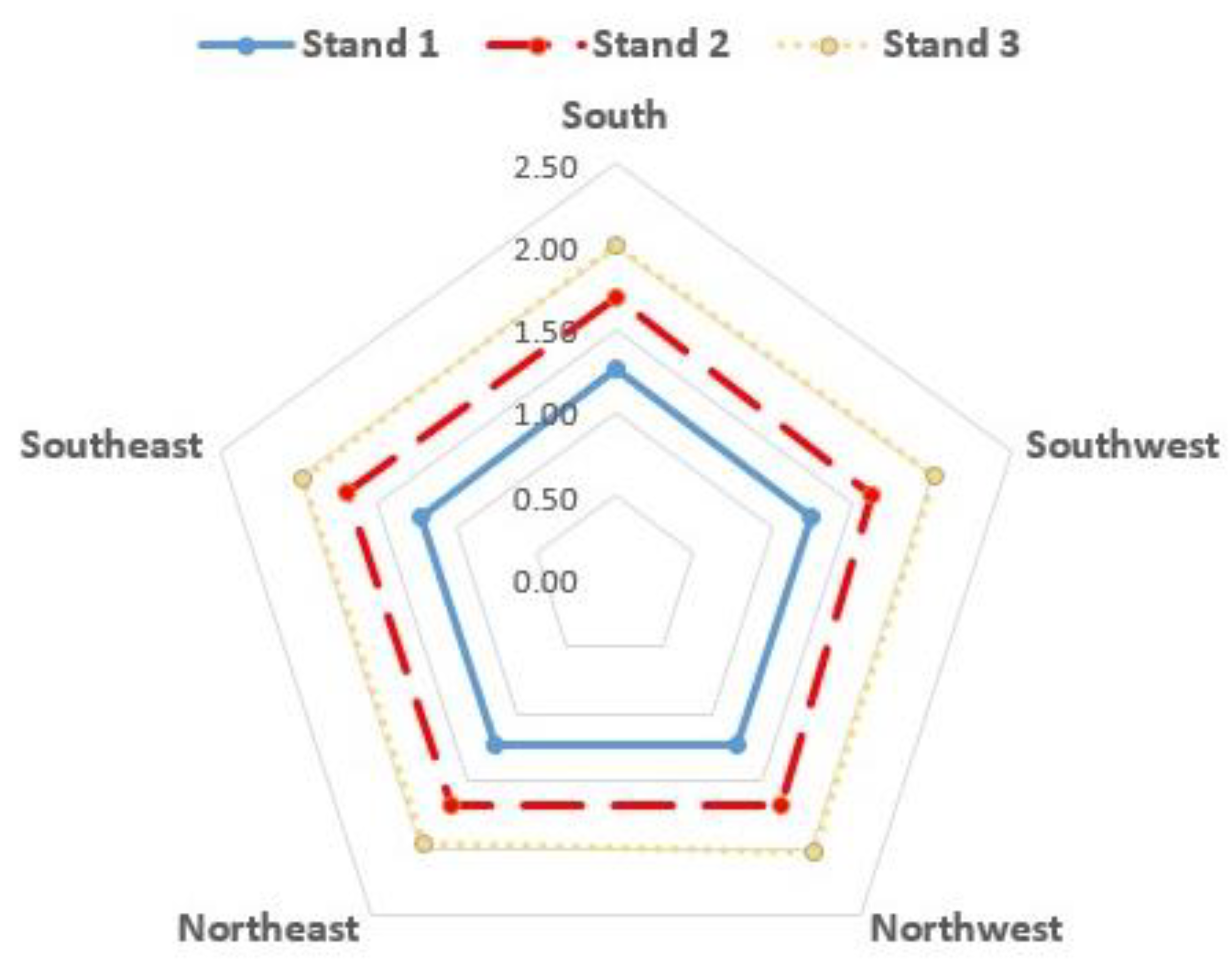
3.1. Bark Thickness
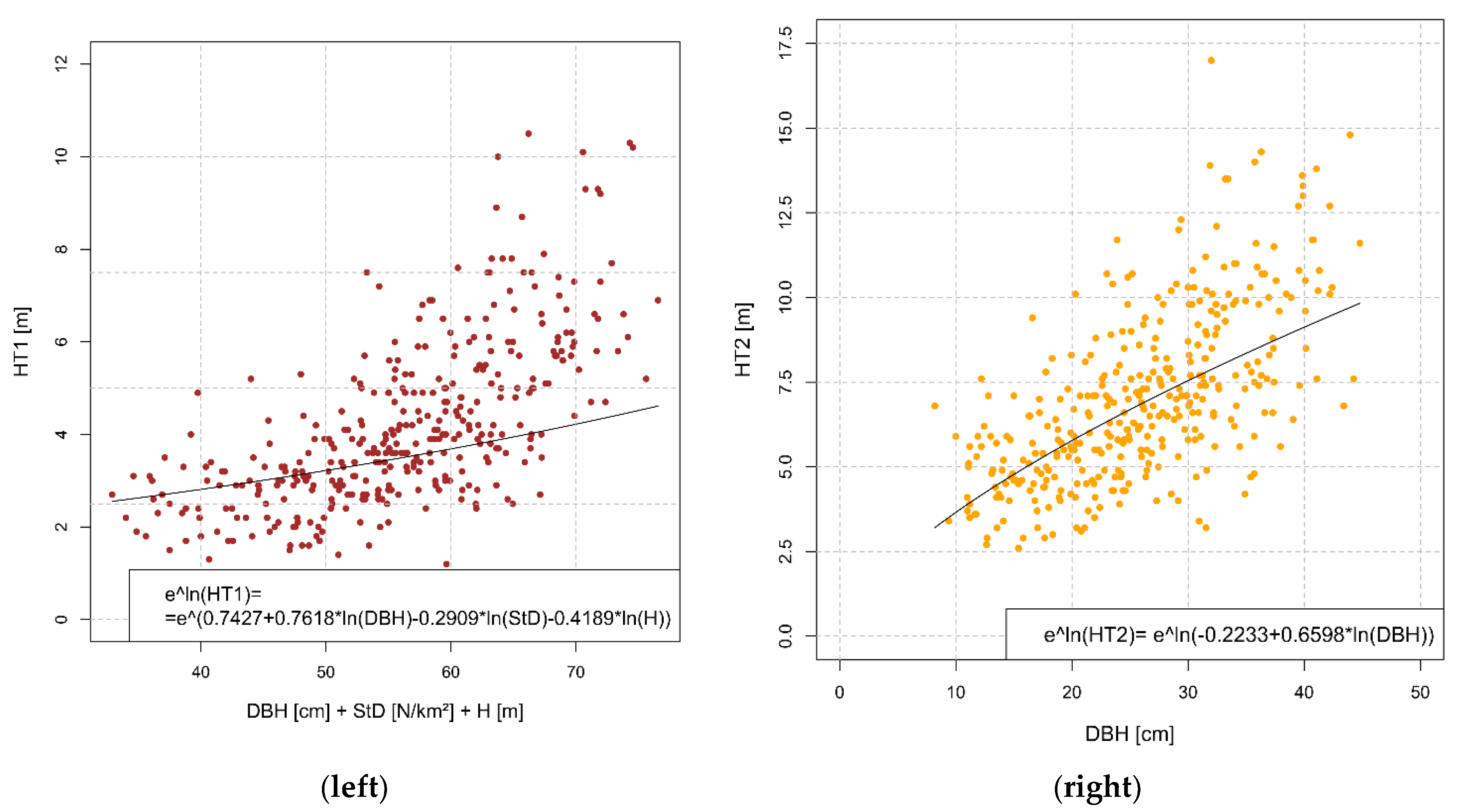
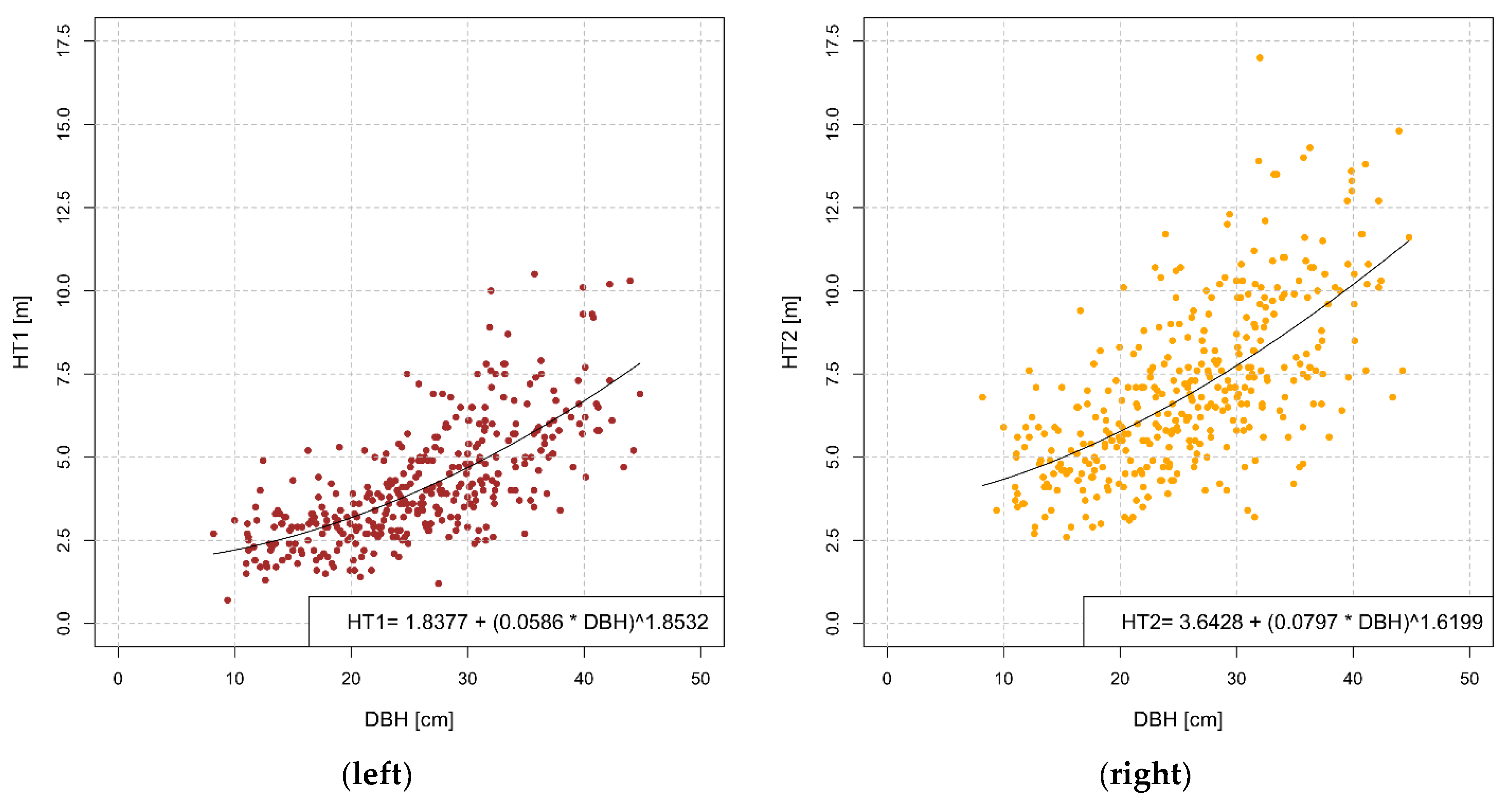
3.2. Bark Transition Heights
4. Discussion
4.1. Bark Thickness
4.2. Bark Transition Heights
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FVA. Baumartenporträt: Die Waldkiefer. Available online: https://www.waldwissen.net/de/lebensraum-wald/baeume-und-waldpflanzen/nadelbaeume/die-waldkiefer (accessed on 14 December 2020).
- BWI³. Dritte Bundeswaldinventur—Ergebnisdatenbank. Available online: https://bwi.info/ (accessed on 12 January 2020).
- Schütt, P.; Stimm, B. Pinus sylvestris. In Enzyklopädie der Holzgewächse: Handbuch und Atlas der Dendrologie/begründet von Peter Schütt; Roloff, A., Weisgerber, H., Lang, U.M., Stimm, B., Eds.; Wiley-VCH: Weinheim, Germany, 2007; ISBN 9783527321414. [Google Scholar]
- Routa, J.; Brännström, H.; Hellström, J.; Laitila, J. Influence of storage on the physical and chemical properties of Scots pine bark. Bioenerg. Res. 2020, 29, 53. [Google Scholar] [CrossRef]
- Bauhus, J.; Kouki, J.; Paillet, Y.; Asbeck, T.; Marchetti, M. How does the forest-based bioeconomy impact forest biodiversity? In Towards a Sustainable European Forest-Based Bioeconomy: Assessment and the Way Forward; Winkel, G., Ed.; European Forest Institute: Joensuu, Finland, 2017; pp. 67–76. ISBN 9789525980417. [Google Scholar]
- Jansone, Z.; Muizniece, I.; Blumberga, D. Analysis of wood bark use opportunities. Energy Procedia 2017, 128, 268–274. [Google Scholar] [CrossRef]
- Eberhardt, T.L. Longleaf Pine Inner Bark and Outer Bark Thicknesses: Measurement and Relevance. South. J. Appl. For. 2013, 37, 177–180. [Google Scholar] [CrossRef]
- Romero, C. Bark: Structure and functional ecology. Adv. Econ. Bot. 2014, 17, 5–25. [Google Scholar]
- Diamantopoulou, M.J.; Özçelik, R.; Yavuz, H. Tree-bark volume prediction via machine learning: A case study based on black alder’s tree-bark production. Comput. Electron. Agric. 2018, 151, 431–440. [Google Scholar] [CrossRef]
- Marshall, H.D.; Murphy, G.E.; Lachenbruch, B. Effects of bark thickness estimates on optimal log merchandising. For. Prod. J. 2006, 87–92. [Google Scholar]
- Neumann, M.; Lawes, M.J. Quantifying carbon in tree bark: The importance of bark morphology and tree size. Methods Ecol. Evol. 2021, 12, 646–654. [Google Scholar] [CrossRef]
- Hammond, D.H.; Varner, J.M.; Kush, J.S.; Fan, Z. Contrasting sapling bark allocation of five southeastern USA hardwood tree species in a fire prone ecosystem. Ecosphere 2015, 6, art112. [Google Scholar] [CrossRef] [Green Version]
- Dantas, V.d.L.; Pausas, J.G. The lanky and the corky: Fire-escape strategies in savanna woody species. J. Ecol. 2013, 101, 1265–1272. [Google Scholar] [CrossRef]
- Diamantopoulou, M.J. Artificial neural networks as an alternative tool in pine bark volume estimation. Comput. Electron. Agric. 2005, 48, 235–244. [Google Scholar] [CrossRef]
- Murphy, G.; Cown, D. Within-tree, between-tree, and geospatial variation in estimated Pinus radiata bark volume and weight in New Zealand. N. Z. J. Sci. 2015, 45, 55. [Google Scholar] [CrossRef] [Green Version]
- Volz, K.-R. Untersuchung über die EIgenschaften der Rinde von Ficht, Kiefer und Buche und ihr Eignung als Rohstoff für Flachpressplatten: Dissertation zur Erlangung des Doktorgrades der Forstlichen Fakultät der Georg-August-Universität Göttingen. Ph.D. Thesis, Georg-August-Universität Göttingen, Göttingen, Germany, 1974. [Google Scholar]
- UNECE. Forest product Conversion Factors for the UNECE Region: Geneva Timber and Forest; Discussion Paper 49. Available online: https://unece.org/fileadmin/DAM/timber/publications/DP-49.pdf (accessed on 8 October 2021).
- Stängle, S.M.; Sauter, U.H.; Dormann, C.F. Comparison of models for estimating bark thickness of Picea abies in southwest Germany: The role of tree, stand, and environmental factors. Ann. For. Sci. 2017, 74, 49. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.C.; Jackson, J.F. Estimating the Allometry of Tree Bark. Am. Midl. Nat. 1995, 134, 99. [Google Scholar] [CrossRef]
- Nicolai, V. The bark of trees: Thermal properties, microclimate and fauna. Oecologia 1986, 69, 148–160. [Google Scholar] [CrossRef]
- Shearman, T.M.; Varner, J.M. Variation in Bark Allocation and Rugosity Across Seven Co-occurring Southeastern US Tree Species. Front. For. Glob. Change 2021, 4, 346. [Google Scholar] [CrossRef]
- Dengler, A. 52jährige finnische und märkische Kiefern im Forstamt Eberswalde: Untersuchungen aus dem Waldbau-Institut der Forstlichen Hochschule Eberswalde. Z. Forst Jagdwes. 1937, 69, 555–566. [Google Scholar]
- Wagenknecht, E. Untersuchungen über den Spiegelrindenanteil verschiedener Kiefernrassen im Zusammenhang mit der Ästigkeit. Z. Forst Jagdwes. 1939, 21, 505–526. [Google Scholar]
- Erteld, W. Die Bedeutung der Spiegelrinde an der Kiefer. Sozial. Forstwirtsch. 1963, 13, 331–335. [Google Scholar]
- Houston Durrant, T.; de Rido, D.; Caudullo, G. Pinus sylvestris in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; European Commission: Luxembourg, 2016; pp. 132–133. [Google Scholar]
- Altherr, E.; Unfried, P.; Hradetzky, V. Statistische Rindenbeziehungen als Hilfsmittel zur Ausformung und Aufmessung unentrindeten Stammholzes: Teil 1: Kiefer, Buche, Hainbuche, Esche und Roterle; Forstlichen Versuchs- und Forschungsanstalt Baden-Württemberg: Freiburg, Germany, 1974; Volume 61. [Google Scholar]
- Hamilton, G.J. Forest Mensuration Handbook; Her Majesty’s Stationery Office: London, UK, 1985. [Google Scholar]
- Sedmíková, M.; Löwe, R.; Jankovský, M.; Natov, P.; Linda, R.; Dvořák, J. Estimation of Over- and Under-Bark Volume of Scots Pine Timber Produced by Harvesters. Forests 2020, 11, 626. [Google Scholar] [CrossRef]
- Berendt, F.; Pegel, E.; Blasko, L.; Cremer, T. Bark proportion of Scots pine industrial wood. Eur. J. Wood Wood Prod. (Holz Roh Werkst.) 2021, 128, 268. [Google Scholar] [CrossRef]
- DWD. CDC Open Data. Available online: https://opendata.dwd.de/climate_environment/CDC/observations_germany/climate/annual/more_precip/recent/ (accessed on 18 April 2021).
- LFE. Forstliche Standortskartierung Brandenburg. 2021. Available online: http://www.brandenburg-forst.de:80/geoserver/wms?request=GetCapabilities& (accessed on 29 April 2021).
- Stängle, S.M.; Weiskittel, A.R.; Dormann, C.F.; Brüchert, F. Measurement and prediction of bark thickness in Picea abies: Assessment of accuracy, precision, and sample size requirements. Can. J. For. Res. 2016, 46, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Freise, C. Die relative Kronenlänge als Steuerungsparameter des Einzelbaumwachstums der Fichte. Ph.D. Thesis, Albert-Ludwigs-Universität Freiburg, Freiburg, Germany, 2005. [Google Scholar]
- Epp, P. Zur Abschätzung des Zuwachspotenzials von Fichten und Tannen nach starker Freistellung in Plenterüberführungsbeständen; Albert Ludwigs Universität Freiburg: Freiburg, Germany, 2003. [Google Scholar]
- Gerecke, K.-L. Herleitung und Anwendung von “Referenzbäumen” zur Beschreibung des Wachstumsganges vorherrschender Tannen. Ph.D. Thesis, Albert-Ludwigs-Universität Freiburg, Freiburg, Germany, 1988. [Google Scholar]
- Kramer, H. Kronenaufbau und Kronenentwicklung gleichalter Fichtenpflanzbestände. Allg. Forst Jagdztg. 1962, 133, 249–256. [Google Scholar]
- Kramer, H.; Dong, P.H. Kronenanalyse für Zuwachsuntersuchungen in immissionsgeschädigten Nadelholzbeständen. Forst Holz 1985, 40, 115–118. [Google Scholar]
- Spathelf, P. Orientierungshilfe zur Prognose und Steuerung des Wachstums von Fichten (Picea abies (L.) Karst.) und Tannen (Abies alba Mill.) in Überführungswäldern mit Hilfe der relativen Kronenlänge. Ph.D. Thesis, Albert-Ludwigs-Universität Freiburg, Freiburg, Germany, 1999. [Google Scholar]
- R Core Team. The R Project for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2002; Available online: https://www.R-project.org/ (accessed on 18 April 2021).
- RStudio Team. R Studio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 18 April 2021).
- Wooldridge, J.M. Introductory Econometrics: A Modern Approach, 7th ed.; Cengage Learning South-Western: Cincinnati, OH, USA, 2020; ISBN 9781337558860. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7137-0. [Google Scholar]
- Kuhn, M. Caret: Classification and Regression Training. 2020. Available online: https://CRAN.R-project.org/package=caret (accessed on 26 September 2021).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Elzhov, T.V.; Mullen, K.M.; Spiess, A.-N.; Bolker, B. minpack.lm: R Interface to the Levenberg-Marquardt Nonlinear Least-Squares Algorithm Found in MINPACK, Plus Support for Bounds. 2016. Available online: https://CRAN.R-project.org/package=minpack.lm (accessed on 26 September 2021).
- Laasasenaho, J.; Melkas, T.; Aldén, S. Modelling bark thickness of Picea abies with taper curves. For. Ecol. Manag. 2005, 206, 35–47. [Google Scholar] [CrossRef]
- Kahriman, A. A bark thickness model vor calabrian pine in Turkey. In Proceedings of the 2nd International Conference on Science, Ecology and Technology, Barcelona, Spain, 14–16 October 2016; pp. 661–670. [Google Scholar]
- Sonmez, T.; Keles, S.; Tilki, F. Effect of aspect, tree age and tree diameter on bark thickness of Picea orientalis. Scand. J. For. Res. 2007, 22, 193–197. [Google Scholar] [CrossRef]
- Gordon, A. Estimating bark thickness of Pinus Radiata. N. Z. J. For. Sci. 1983, 13, 340–353. [Google Scholar]
- Liepins, J.; Liepins, K. Evaluation of bark volume of four tree species in Latvia. Res. Rural. Dev. 2015, 2, 22–28. [Google Scholar]
- Berendt, F.; de Miguel-Diez, F.; Wallor, E.; Blasko, L.; Cremer, T. Comparison of different approaches to estimate bark volume of industrial wood at disc and log scale. Sci. Rep. 2021, 11, 15630. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Stand 1 | Stand 2 | Stand 3 | All |
|---|---|---|---|---|
| WGS84 Coordinates | 13.762056, 52.818503 | 13.752174, 52.82378 | 13.744799, 52.815309 | |
| Plot radius (in m) | 12.62 | 15.45 | 17.84 | |
| Cumulative plot area (in m²) | 1500 | 2250 | 3000 | |
| Stand age (in years) | 64 | 69 | 84 | |
| Trees measured (N) | 156 | 118 | 101 | 375 |
| Stand density (N/ha) | 1040 | 524 | 337 | |
| Mean DBH ± SD (in cm) | 18.5 ± 5.2 | 27.8 ± 4.9 | 34.5 ± 5.3 | 25.7 ± 8.4 |
| Mean BT ± SD (in cm) | 1.24 ± 0.41 | 1.68 ± 0.39 | 1.99 ± 0.39 | 1.58 ± 0.50 |
| Mean H ± SD (in m) | 18.9 ± 2.8 | 25.5 ± 2.0 | 26.8 ± 1.9 | 23.1 ± 4.3 |
| Mean HC ± SD (in m) | 14.1 ± 2.2 | 19.2 ± 2.2 | 19.1 ± 2.3 | 17.0 ± 3.3 |
| Mean HT 1 ± SD (in m) | 3.0 ± 0.9 | 4.3 ± 1.5 | 5.8 ± 1.7 | 4.2 ± 1.8 |
| Mean HT2 ± SD (in m) | 5.6 ± 1.8 | 7.1 ± 2.2 | 9.1 ± 2.5 | 7.0 ± 2.5 |
| Variables | ln(HT1) | ln(DBH) | ln(H) | ln(HC) | ln(StD) | ln(PD) | Variables |
|---|---|---|---|---|---|---|---|
| 1 | 0.7293 | 0.6391 | 0.4975 | −0.6675 | −0.6668 | ln(HT1) | |
| ln(HT2) | 1 | 1 | 0.8486 | 0.6238 | −0.8104 | −0.8135 | ln(DBH) |
| ln(DBH) | 0.6553 | 1 | 1 | 0.8393 | −0.8250 | −0.8120 | ln(H) |
| ln(H) | 0.5532 | 0.8486 | 1 | 1 | −0.7063 | −0.6960 | ln(HC) |
| ln(HC) | 0.4157 | 0.6238 | 0.8393 | 1 | 1 | 0.9439 | ln(StD) |
| ln(StD) | −0.5409 | −0.8104 | −0.8250 | −0.7063 | 1 | 1 | ln(PD) |
| ln(PD) | −0.5461 | −0.8135 | −0.8120 | −0.6960 | 0.9439 | 1 | |
| ln(HT2) | ln(DBH) | ln(H) | ln(HC) | ln(StD) | ln(PD) |
| Variable | Estimate | SE | p-Value | Adj-R² | RSE |
|---|---|---|---|---|---|
| ln(HT1) | 0.5135 | 0.2965 | |||
| Intercept | 0.7427 | 0.5469 | 0.175 | ||
| Coef. ln(DBH) | 0.7618 | 0.0999 | 2.11 × 10−13 | ||
| Coef. ln(StD) | −0.2909 | 0.1985 | 0.0355 | ||
| Coef. ln(H) | −0.4189 | 0.0586 | 1.07 × 10−6 | ||
| ln(HT2) | 0.3935 | 0.2781 | |||
| Intercept | −0.2233 | 0.1378 | 0.106 | ||
| Coef. ln(DBH) | 0.6598 | 0.0428 | 1.19 × 10−41 |
| Variable | Estimate | SE | p-Value | RSE |
|---|---|---|---|---|
| HT1 | 1.2527 | |||
| Intercept a | 1.8376 | 0.3766 | 1.59 × 10−6 | |
| b | 0.0586 | 0.0102 | 1.85 × 10−8 | |
| c | 1.8532 | 0.3121 | 6.79 × 10−9 | |
| HT2 | 1.9323 | |||
| Intercept a | 3.6428 | 0.7179 | 6.23 × 10−7 | |
| b | 0.0797 | 0.0247 | 1.38 × 10−3 | |
| c | 1.6199 | 0.3613 | 9.84 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilms, F.; Duppel, N.; Cremer, T.; Berendt, F. Bark Thickness and Heights of the Bark Transition Area of Scots Pine. Forests 2021, 12, 1386. https://doi.org/10.3390/f12101386
Wilms F, Duppel N, Cremer T, Berendt F. Bark Thickness and Heights of the Bark Transition Area of Scots Pine. Forests. 2021; 12(10):1386. https://doi.org/10.3390/f12101386
Chicago/Turabian StyleWilms, Florian, Nils Duppel, Tobias Cremer, and Ferréol Berendt. 2021. "Bark Thickness and Heights of the Bark Transition Area of Scots Pine" Forests 12, no. 10: 1386. https://doi.org/10.3390/f12101386
APA StyleWilms, F., Duppel, N., Cremer, T., & Berendt, F. (2021). Bark Thickness and Heights of the Bark Transition Area of Scots Pine. Forests, 12(10), 1386. https://doi.org/10.3390/f12101386