2.1. Inventory and Site Data
Retrospective inventory and site data were derived from UTHF, located in Furano (central Hokkaido, Japan). This site experiences a mean annual temperature and precipitation of 6.4 °C and 1297 mm, respectively [
35]. Dominant stratified soils include brown forest soil, dark brown forest soil, black soil, and podzol [
36]. Mixed conifer–hardwood forest, typical of the cool temperate zone, covers most of the site area. Tree species commonly found in this forest area are
Fraxinus mandshurica,
Ulmus davidiana var
. japonica,
Alnus hirsuta, and
Salix spp. in deciduous swamp forests at lower elevations of less than 300 m; a coniferous and broad-leaved mixed forest dominated by
A. sachalinensis at middle elevations (300–600 m), scattered forests mixed with
P. jezoensis,
Picea glehnii, and
Betula ermanii at upper elevations (800–1200 m), and alpine vegetation (e.g.,
Pinus pumila) in the upper forest limit (>1200 m).
The Stand-based Silvicultural Management System (SSMS), in other words, a natural forest management system based on selected cutting and natural regeneration, is currently being employed extensively in UTHF except for research plots [
37,
38]. In SSMS, 10–17% of the stand volume is harvested by single-tree selection, with a cutting cycle of 15–20 years, by removing defective (e.g., diseased, senescent, non-vigorous, and twisted) and over-matured trees. This system helps maintain tree health and productivity in the stand and controls the stand composition [
37]. To determine cutting rates, permanent plots and long-term ecological research plots have been established and periodically assessed. Among these permanent plots, 25 are located in the preserved area, ranging in size from 0.04 to 2.25 ha, and have an elevation between 380 and 1290 m. Within these plots, DBH measurements of all trees with a DBH ≥ 5 cm are regularly performed by UTHF staff; in most cases, 5-year interval and census data are available for the last five decades. Other than these periodical measurements, no human intervention has occurred in this preserved area for several decades.
2.2. Survival Data
Survival data were derived from periodic surveys on preserved permanent sample plots. We selected four plots, ranging in size from 0.04 to 2.25 ha, located at an elevation range of 570 to 690 m with similar slope aspects and slope angles (
Figure 1;
Table 1). The main soil types found in the plots are brown forest and podzolic soils [
35]. In addition, the stands in the four plots are all classified as “coniferous selective cutting with poor regeneration” in UTHF, where no continuous sufficient and new in-growth trees are expected. All four plots were within close proximity and had similar species composition. The typical vegetation of the plots was coniferous and broad-leaved mixed forests dominated by
A. sachalinensis,
P. jezoensis,
Acer spp., and
Tilia spp. Therefore, the four plots were aggregated in further analyses on tree survival.
We used tree census data (species, DBH, living or dead status, cause of death, etc.) of the plots, obtained between 1989 and 2019. Within this period, we set three observation periods of 1989–1999 (period 1), 1999–2009 (period 2), and 2009–2019 (period 3). We detected the number of tree rings at breast height (1.3 m) in 2019 using a semi-nondestructive device, RESISTOGRAPH
® [
39]. All target trees were living and had a DBH ≥ 5 cm in 1989; some of these trees died by 2019. Live/dead trees and new in-growth trees were counted at the end of each observation period. The connected RESISTOGRAPH (Heidelberg, Germany) data logger recorded measurements, which were transferred to a computer for further analyses. The field measurement tree ring data (radius at breast height −2.5 cm) in 2019 were extracted via the DECOM
TM software (Philadelphia, PA, USA), which is used for annual tree ring detection. The “radius at breast height −2.5 cm” reveals the tree age after in-growth if we set DBH = 5 cm as an in-growth border.
Figure 2 shows the annual rings in one living
A. sachalinensis tree detected using the DECOM
TM software. RESISTOGRAPH measurements were ineffective for severely rotten and center-decayed trees, for which we established the regression equation between the age after in-growth (
y) and the radius −2.5 cm (
w) [
40]. Next, the age after in-growth was estimated by inputting the radius data into simple three-dimensional equations fitted to a scatter diagram (
Table 2). This method was followed for 20% of the sample trees.
Table 3 further illustrates the method applied to determine the in-growth years of trees.
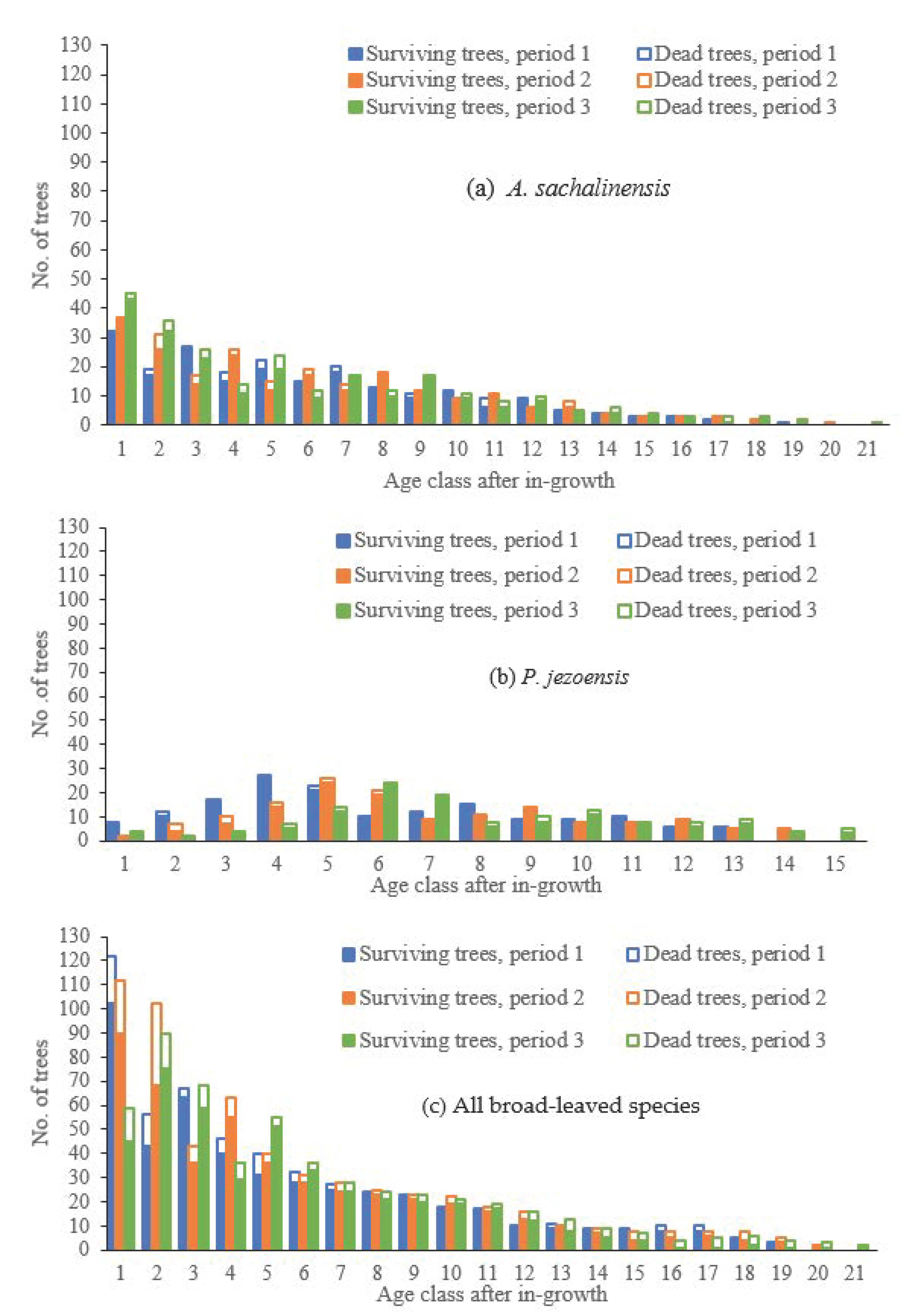
Survival models were built for two major coniferous species present in UTHF,
A. sachalinensis and
P. jezoensis. All broad-leaved trees were considered collectively to develop survival models. The model data of period 3 were for 260
A. sachalinensis, 138
P. jezoensis, and 521 broad-leaved trees (see
Table 3 for the number of dead and living trees in period 3 included in this study). The ages of target trees in period 3 were first identified using the above-mentioned methods. The ages in periods 1 and 2 were calculated by deducting 10 and 20 years from those in period 3. Finally, tree ages were classified, with 10 years per age class.
2.3. Fundamentals of Survival Analysis of Major Species
Survival probability functions were the essential elements of this analysis, as they reflect the overall performance of the major species; they were developed based on previous studies [
20,
22,
32,
41,
42,
43]. The survival data demonstrated that both dead and living trees were present in the study plots at the end of the observation period. When we carry out survival analysis, the issue of missing data appears most of the time. Therefore, the censoring and truncation techniques must be incorporated to overcome this issue. Right censoring occurs when a target leaves the study before an event occurs or the follow-up ends before mortality happens. In contrast, left truncation occurs when an object is not observed from the beginning of the study, but rather enters the study at a later time in the observation period [
20,
44]. Furthermore, if target trees survived at the
t-th age class in the observation period, then observed mortalities were truncated and censored at both beginning and end of the observation period.
Mortality rates (
) were calculated using age class after in-growth (
T), according to Equation (1):
where
is the mortality rate in the
t-th age class.
If a tree survives the
t-th age class during the observation period, the conditional probability is defined as follows:
Following previous methods [
45,
46,
47] and considering Equations (1) and (2), we described the likelihood function (
L) of the observation as follows:
where
dt is the number of dead trees and
at is the number of surviving trees in the
t-th age class during the observation period.
The mortality probability (
) of a new in-growth tree in the
t-th age class was defined as follows:
The survival probability (
) in the
t-th age class was defined as follows:
Therefore, based on Equation (1), the mortality rate (
) can be expressed as follows:
The maximum likelihood estimators of
can be calculated by the first-order derivation of Equation (3), as shown in Equation (7):
Considering that Equations (5) and (6) can be combined as follows:
the survival function can be converted into:
Equation (9) can also be expressed as follows:
The survival function is described by the following formula:
where
represents the survival function and
represents the hazard function in the
t-th age class.
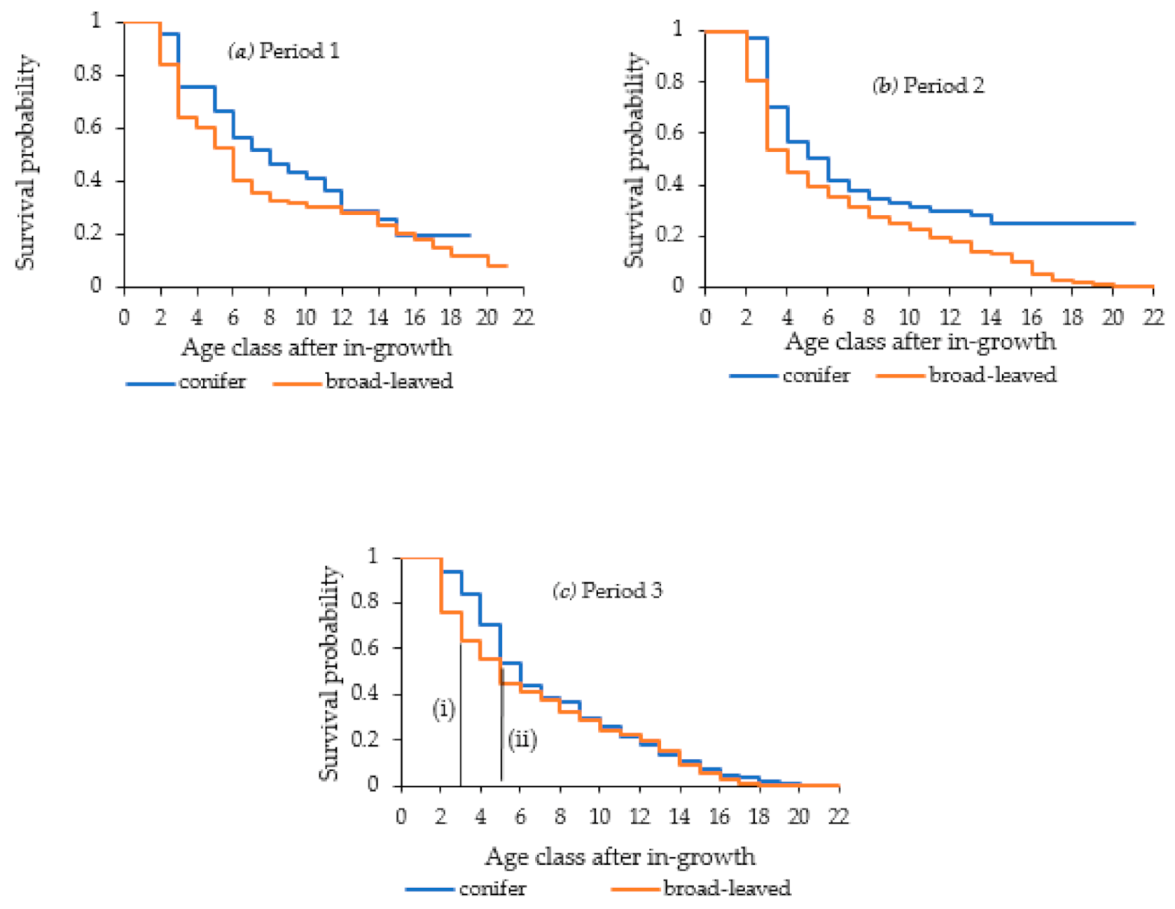
This
is the Kaplan–Meier estimate, which is an important tool for analyzing censored data [
23]. Survival analysis is performed to describe the distribution of tree mortality using Kaplan–Meier estimates in the form of step-wise curves [
23].
It is meaningful to assess whether there are differences in survival (or cumulative incidence of the event) among different groups. For this purpose, there are several tests available to compare survival among independent groups.
Survival probabilities can be compared using the log-rank test [
48], which tests a null hypothesis (i.e., no significant difference in survival between consecutive periods in this study) and the expectation of an equal number of deaths (
E) in each of the two groups of each species. The observed (i.e., real) number of deaths is indicated by
O in the following equation:
Wilcoxon test [
49] can be used as an alternative to the log-rank test. It emphasizes the information at the beginning of the survival curve where the number at risk is significant, allowing early failures to receive more weight than later failures:
It weights the observed minus expected score at the time tf by the number at risk, w(tf) overall groups at time tf.
Mean lifetime, which is also expressed as mean longevity or mean lifespan, can be considered as the area under survival curve. It is mathematically calculated as the weighted sum of the age and estimated mortality probability described previously herein and can be expressed as follows:
It is meaningful to calculate not only mean lifetime but also “practical” mean lifetime from the forest management perspective based on the survival estimates for the two groups of trees (coniferous and broad-leaved). Common mean lifetime was derived by considering all the age classes whereas practical mean lifetime used only specific age class following the Kaplan–Meier estimates. The particular point of the practical mean lifetime is that it has avoided the drastic decrease of survival probabilities in younger age classes, which commonly happen in natural forest stands due to suppression. We intended to explore the possibility of using the practical mean lifetime for kinds of harvesting standards in tree marking process, so that too short mean lifetime was inconvenient. Finally, these practical mean lifetime values were converted into DBH values by using the equations of
Table 2 to facilitate practical application in the tree marking process of SSMS.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}