
Pollen Competition and Paternal Contribution during Artificially Controlled Pollination of Black Locust (Robinia pseudoacacia L.) without Castration

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Controlled Pollination and DNA Extraction
2.2. PCR Amplification with SSR Primers
2.3. Paternity and Data Analysis
3. Results
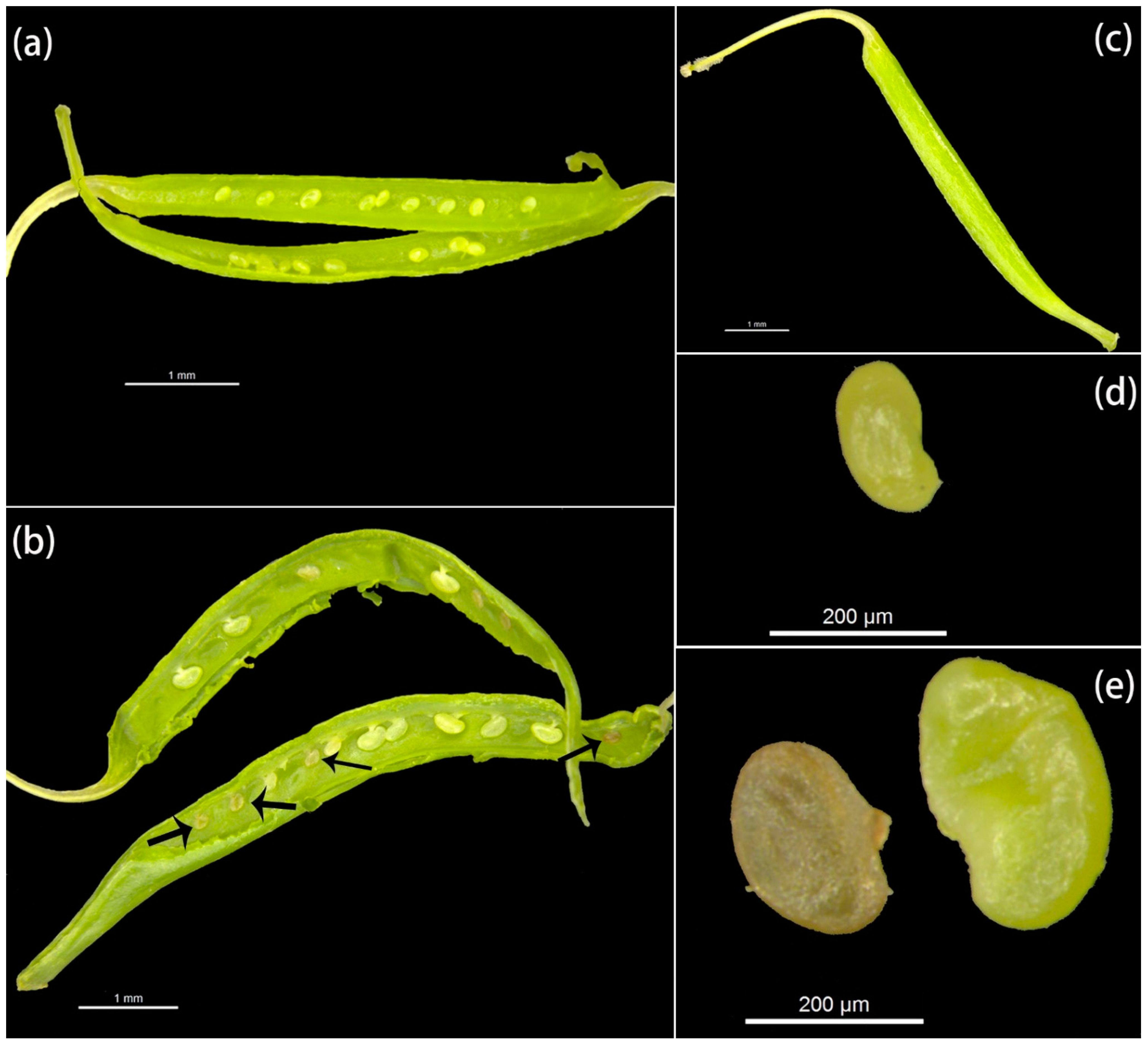
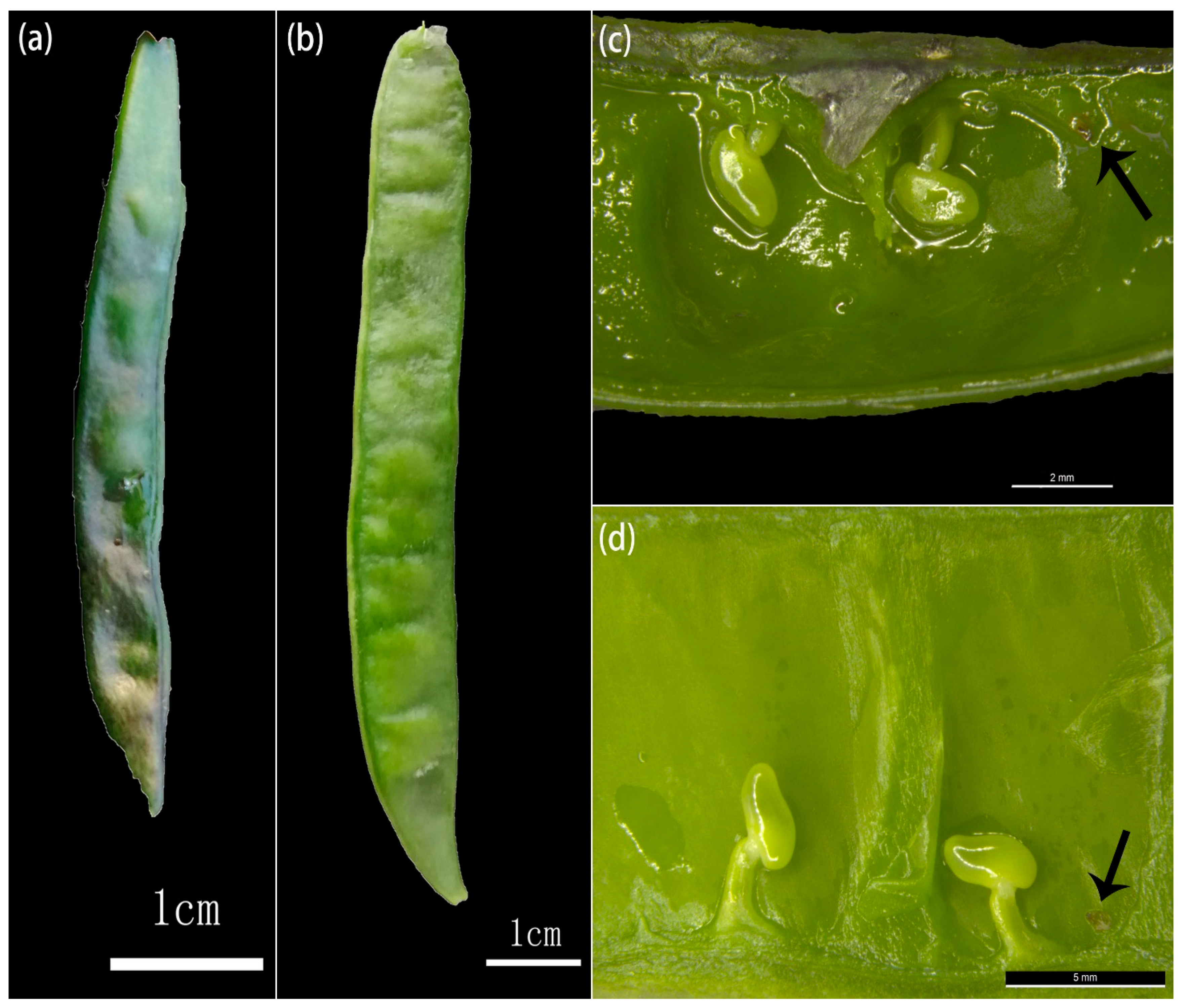
3.1. Early Development of Offspring
3.2. Composition of Immature Zygotic Embryo Pollen Donors According to Developmental Stage
3.3. Development and Composition of Pollen Donors of Aborted Seeds
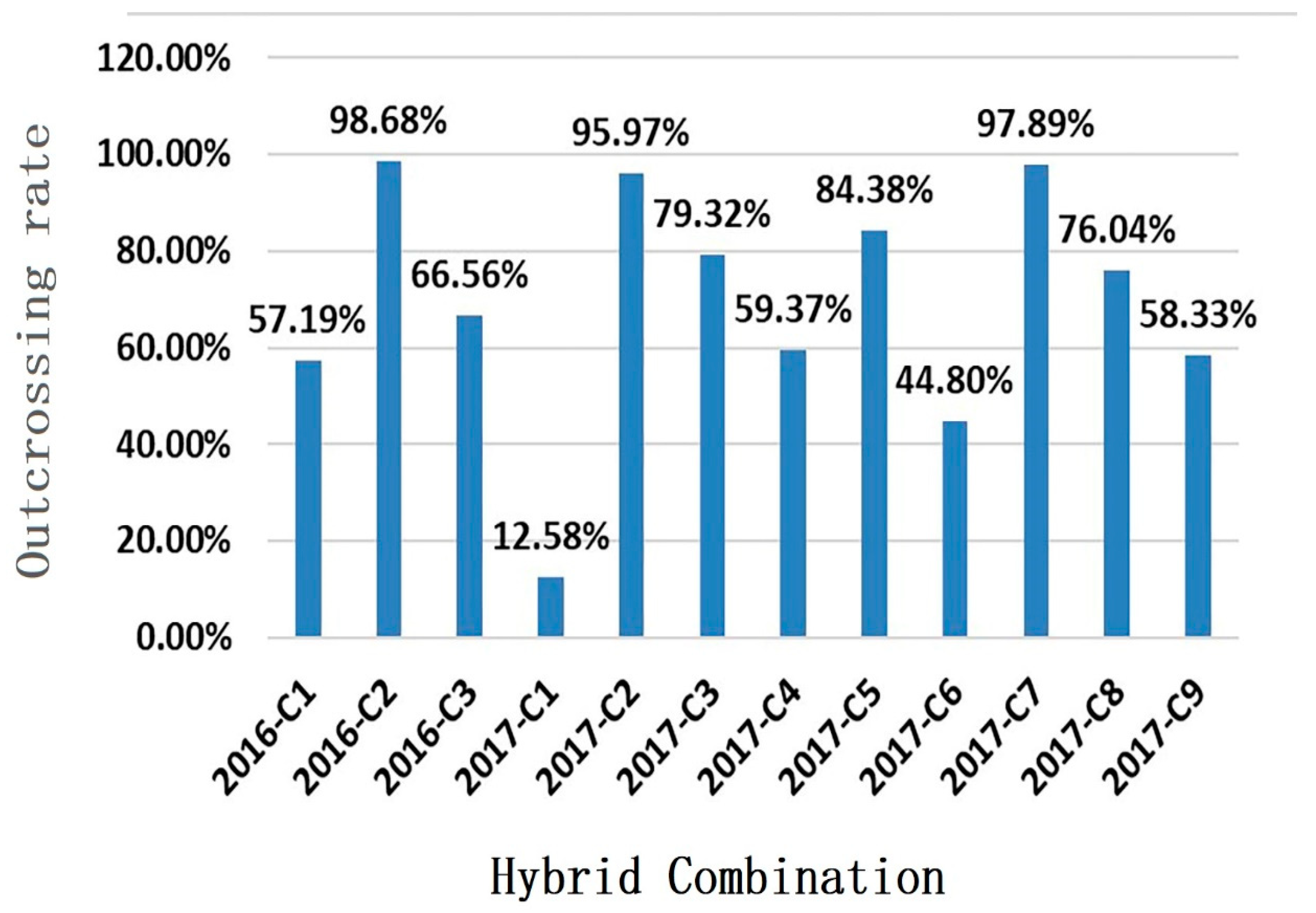
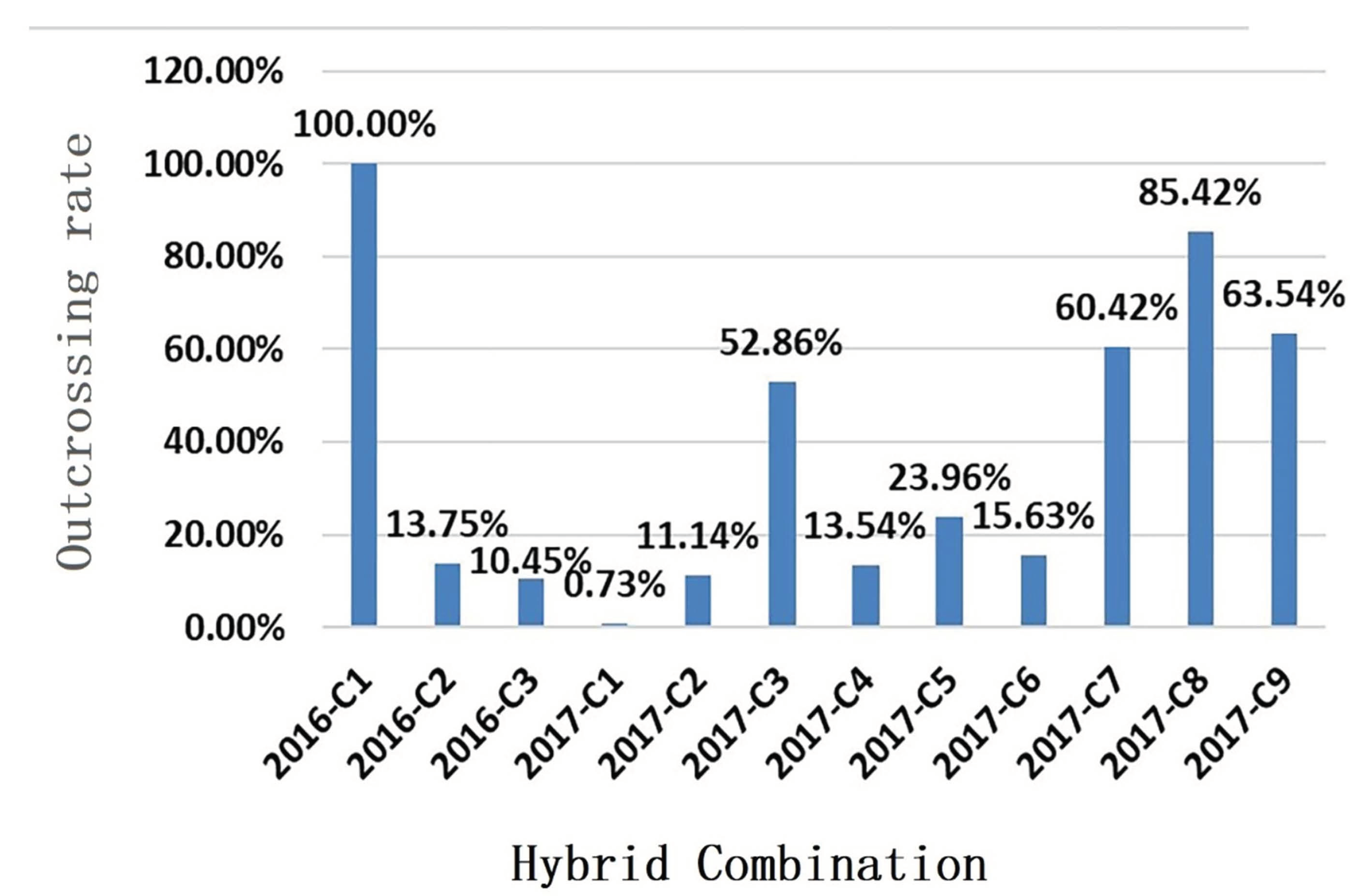
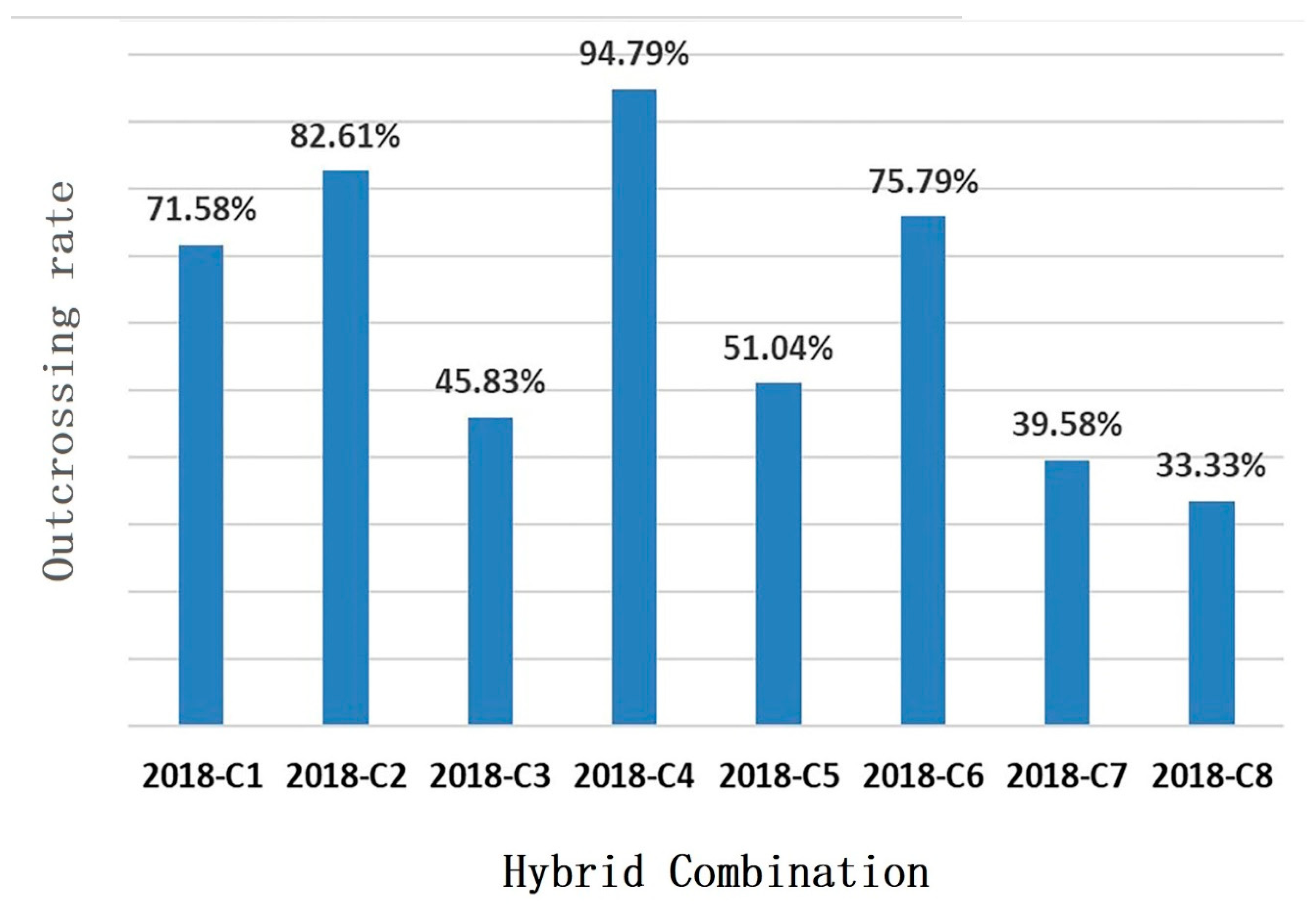
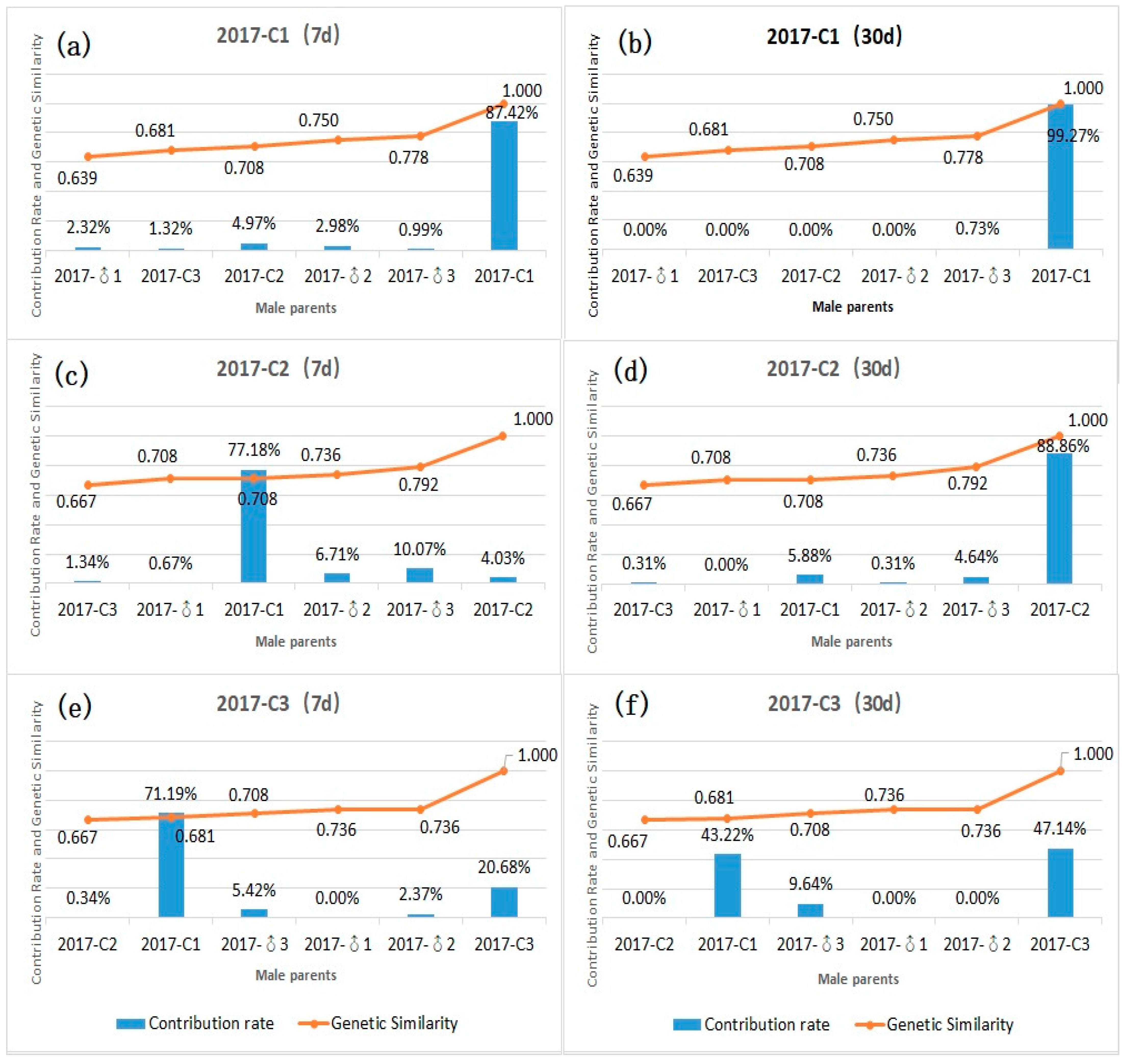
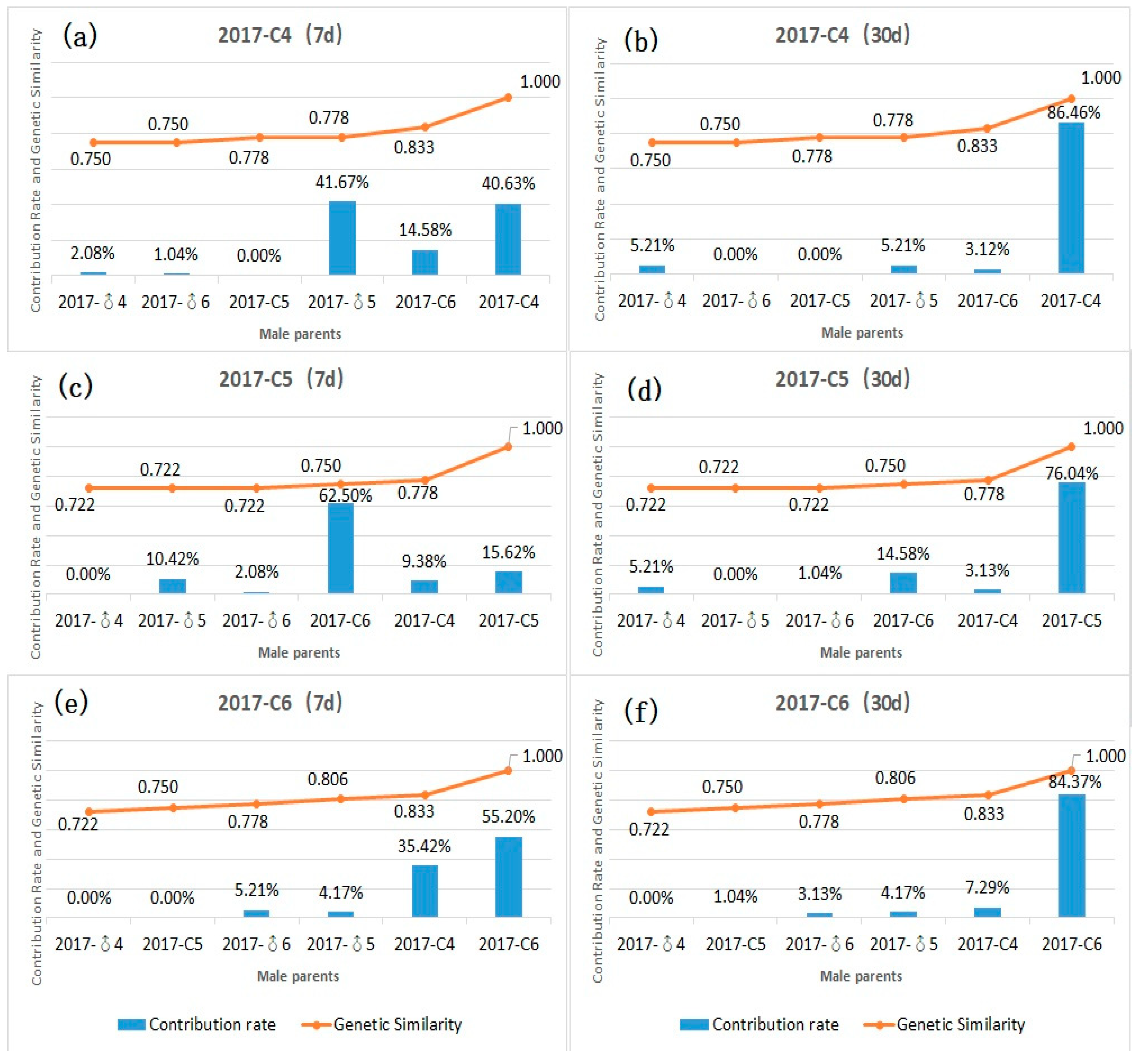
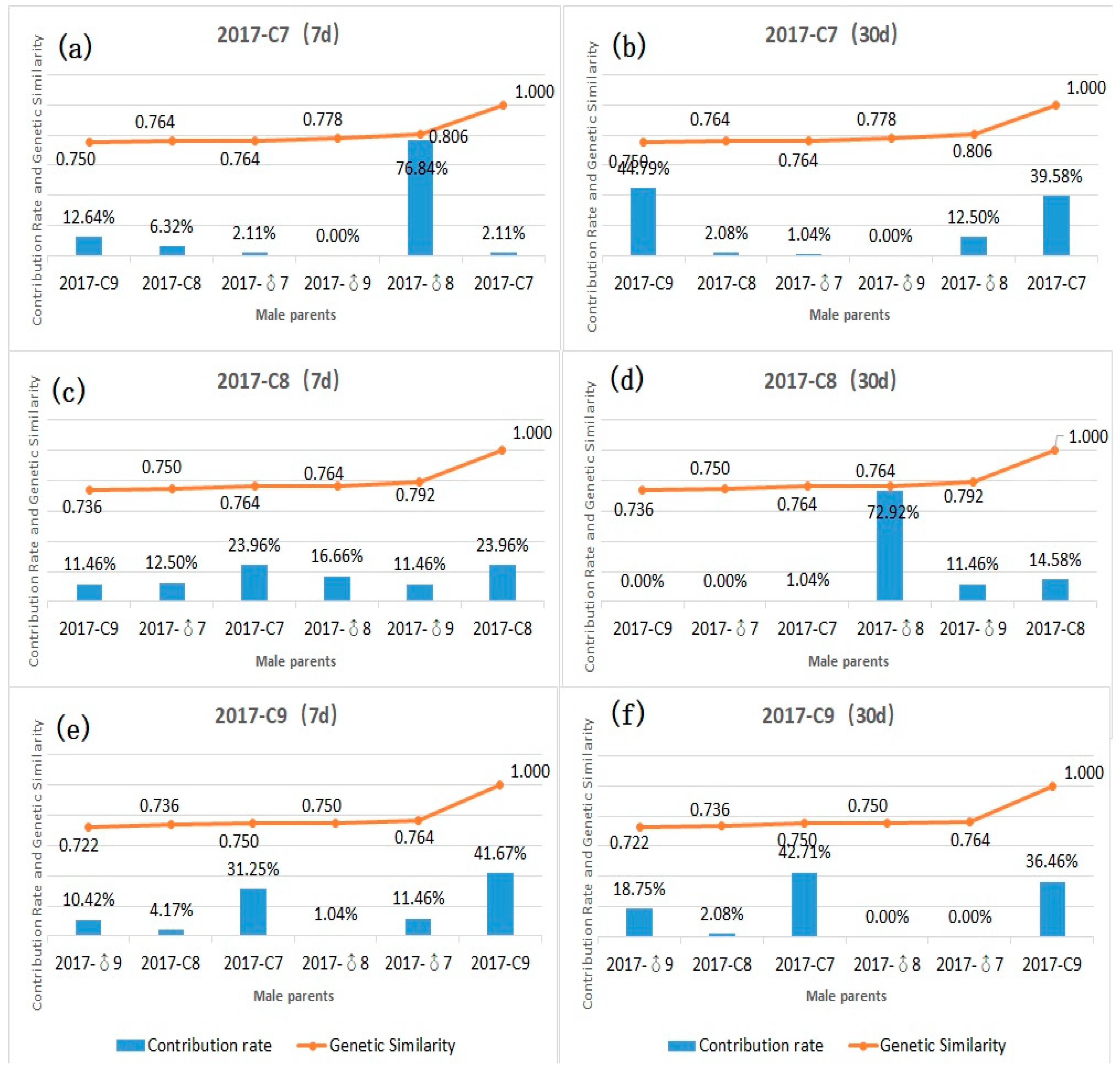
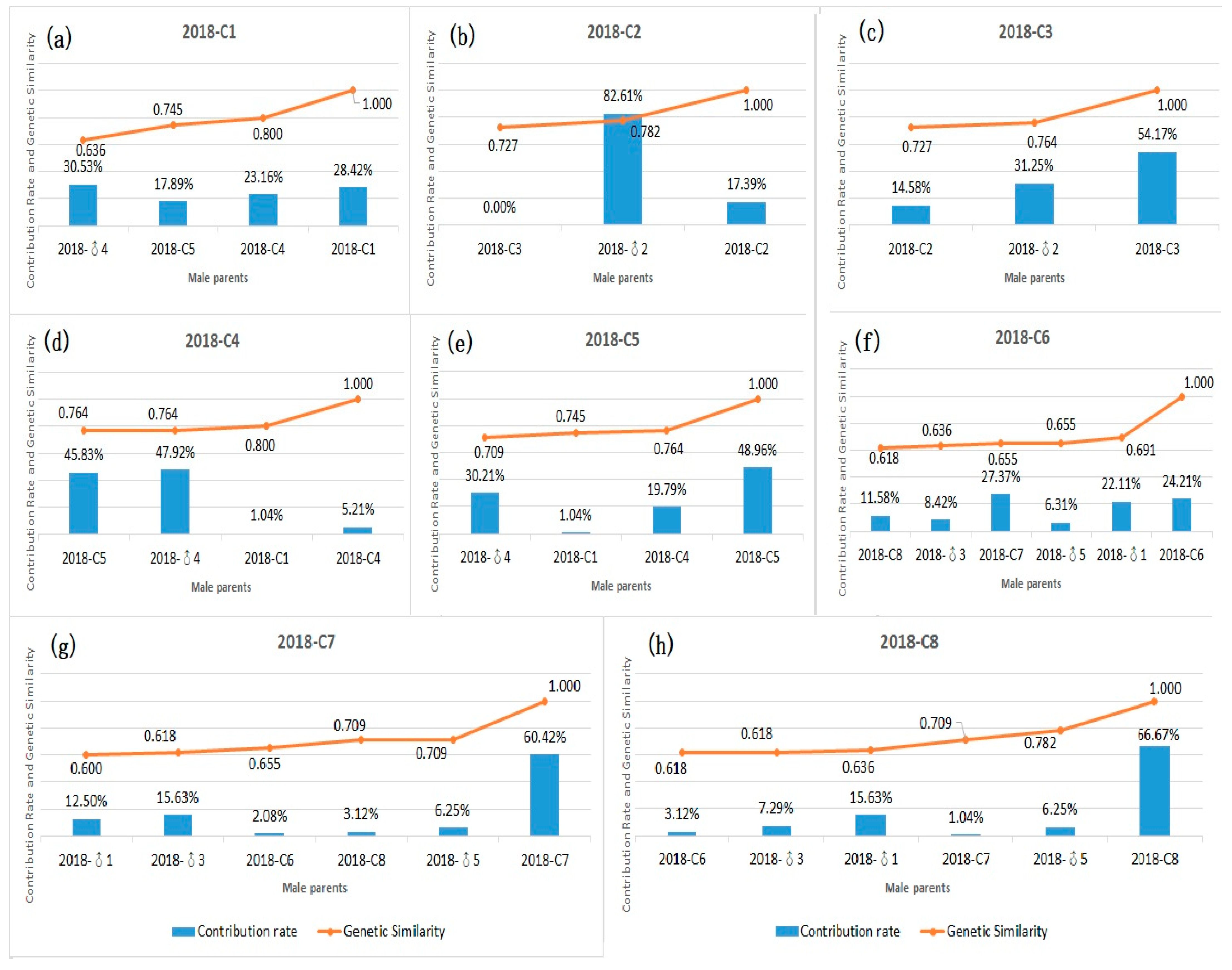
3.4. Contribution Rate and Relationship between Genetic Similarity and Hybrid Affinity of Parents
4. Discussion
4.1. Pollen Competition and Offspring Selection
4.2. Mating System of Black Locust
4.3. Controlled Pollination of Black Locust without Castration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Barrett, R.P.; Mebrahtu, T.; Hanover, J.W. Black Locust: A Multi-Purpose Tree Species for Temperate Climates; Advances in New Crops; Timber Press: Portland, OR, USA, 1990; pp. 278–283. [Google Scholar]
- Bongarten, B.C.; Huber, D.A.; Apsley, D.K. Environmental and genetic influences on short-rotation biomass production of black locust (Robinia pseudoacacia L.) in the Georgia Piedmont. For. Ecol. Manag. 1992, 55, 315–331. [Google Scholar] [CrossRef]
- Li, S.W.; Zhang, Z.Y.; He, C.Z.; An, X.M. Progress on hybridization breeding of poplar in China. World For. Res. 2004, 17, 37–41. [Google Scholar]
- Zhang, Z.Y.; Li, S.W.; He, Z.G. Research status and development strategy of poplar resources and hybrid breeding in China. J. Hebei For. Sci. Technol. 2006, S1, 20–24. [Google Scholar] [CrossRef]
- Wei, Y.C.; Li, Z.Q.; Li, Y.; Chang, L. Genetic analysis of morphological traits of Eucommia ulmoides Oliver’s hybrid offspring. J. Northwest A F Univ. (Nat. Sci. Ed.) 2012, 40, 137–143. [Google Scholar]
- Wu, K.M.; Wu, J.Y.; Gan, S.M. Hybridization and Heterosis in Eucalyptus. For. Environ. Sci. 2001, 17, 10–15. [Google Scholar]
- Mao, X.H. Analysis of Phenotypic Variation and Genetic Diversity for Robinia Pseudoacacia Clonal Germplasms from Different Sources; Chinese Academy of Forestry: Beijing, China, 2017. [Google Scholar]
- Gu, J.T. Studies on Genetic Structure and Genetic Diversity in Provenances of Black Locust (Robinia pseudoacacia L.) in China; Hebei Agricultural University: Baoding, China, 2006. [Google Scholar]
- Susko, D.J. Effect of ovule position on patterns of seed maturation and abortion in Robinia pseudoacacia (Fabaceae). Botany 2006, 84, 1259–1265. [Google Scholar] [CrossRef]
- Sun, P. Effects of Emasculation on Flower Development and Fruit Set of Controlled Pollination in Robinia pseudoacacia; Beijing Forestry University: Beijing, China, 2012. [Google Scholar]
- Surles, S.E.; Hamrick, J.L.; Bongarten, B.C. Mating systems in open-pollinated families of black locust (Robinia pseudoacacia). Silvae Genet. 1990, 39, 35. [Google Scholar]
- Yuan, C.Q. The sexual Reproduction and Mating System of Robinia pseudoacacia L. (Fabaceae); Beijing Forestry University: Beijing, China, 2013. [Google Scholar]
- Dow, B.D.; Ashley, M.V. High levels of gene flow in bur oak revealed by paternity analysis using microsatellites. J. Hered. 1998, 89, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Charlesworth, B. Inbreeding depression and its evolutionary consequences. Annu. Rev. Ecol. Syst. 1987, 18, 237–268. [Google Scholar] [CrossRef]
- Barrett, S.C.H. Mating strategies in flowering plants: The outcrossing–selfing paradigm and beyond. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 991–1004. [Google Scholar] [CrossRef]
- Lande, R.; Schemske, D.W. The evolution of self-fertilization and inbreeding depression in plants. I. Genetic models. Evolution 1985, 39, 24–40. [Google Scholar] [PubMed]
- Ansaldi, B.H.; Weber, J.J.; Franks, S.J. The role of phenotypic plasticity and pollination environment in the cleistogamous, mixed mating breeding system of Triodanis perfoliata. Plant Biol. 2018, 20, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Goodwillie, C.; Kalisz, S.; Eckert, C.G. The evolutionary enigma of mixed mating systems in plants: Occurrence, theoretical explanations, and empirical evidence. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 47–79. [Google Scholar] [CrossRef] [Green Version]
- Kruszewski, L.J.; Galloway, L.F. Explaining outcrossing rate in Campanulastrum americanum (Campanulaceae): Geitonogamy and cryptic self-incompatibility. Int. J. Plant Sci. 2006, 167, 455–461. [Google Scholar] [CrossRef]
- Marshall, D.L.; Shaner, M.G.; Oliva, J.P. Effects of pollen load size on seed paternity in wild radish: The roles of pollen competition and mate choice. Evolution 2007, 61, 1925–1937. [Google Scholar] [CrossRef] [PubMed]
- Korbecka, G.; Klinkhamer, P.G.L.; Vrieling, K. Selective embryo abortion hypothesis revisited-a molecular approach. Plant Biol. 2002, 4, 298–310. [Google Scholar] [CrossRef]
- Lian, C.; Hogetsu, T. Development of microsatellite markers in black locust (Robinia pseudoacacia) using a dual-supression-PCR technique. Mol. Ecol. Notes 2002, 2, 211–213. [Google Scholar]
- Mishima, K.; Hirao, T.; Urano, S.; Watanabe, A.; Takata, K. Isolation and characterization of microsatellite markers from Robinia pseudoacacia L. Mol. Ecol. Resour. 2009, 9, 850–852. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, J.X.; Su, L.Z.; Lv, W.; Sun, Y.H.; Li, Y. Development and evaluation of a novel set of EST-SSR markers based on transcriptome sequences of black locust (Robinia pseudoacacia L.). Genes 2017, 8, 177. [Google Scholar] [CrossRef] [Green Version]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System, Version 2.1; Applied Biostatistics: Setauket, NY, USA, 2000. [Google Scholar]
- Haldane, J.B. The Causes of Evolution, Volume 5; Princeton University Press: Princeton, NJ, USA, 1990. [Google Scholar]
- Pélabon, C.; Hennet, L.; Bolstad, G.H.; Albertsen, E.; Opedal, Ø.H.; Ekrem, R.K.; Armbruster, W.S. Does stronger pollen competition improve offspring fitness when pollen load does not vary? Am. J. Bot. 2016, 103, 522–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulcahy, D.L. The rise of the angiosperms: A genecological factor. Science 1979, 206, 20–23. [Google Scholar] [CrossRef]
- Walsh, N.E.; Charlesworth, D. Evolutionary interpretations of differences in pollen tube growth rates. Q. Rev. Biol. 1992, 67, 19–37. [Google Scholar] [CrossRef]
- Lankinen, Å.; Skogsmyr, I.O. Evolution of pistil length as a choice mechanism for pollen quality. Oikos 2001, 92, 81–90. [Google Scholar] [CrossRef]
- Armbruster, W.S.; Debevec, E.M.; Willson, M.F. Evolution of syncarpy in angiosperms: Theoretical and phylogenetic analyses of the effects of carpel fusion on offspring quantity and quality. J. Evol. Biol. 2002, 15, 657–672. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, W.S. Evolution of floral morphology and function: An integrative approach to adaptation, constraint, and compromise in Dalechampia (Euphorbiaceae). In Floral Biology; Springer: Boston, MA, USA, 1996; pp. 241–272. [Google Scholar]
- Galen, C.; Shykoff, J.A.; Plowright, R.C. Consequences of stigma receptivity schedules for sexual selection in flowering plants. Am. Nat. 1986, 127, 462–476. [Google Scholar] [CrossRef]
- Lankinen, Å.; Armbruster, W.S.; Antonsen, L. Delayed stigma receptivity in Collinsia heterophylla (Plantaginaceae): Genetic variation and adaptive significance in relation to pollen competition, delayed self-pollination, and mating-system evolution. Am. J. Bot. 2007, 94, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Lankinen, Å.; Madjidian, J.A. Enhancing pollen competition by delaying stigma receptivity: Pollen deposition schedules affect siring ability, paternal diversity, and seed production in Collinsia heterophylla (Plantaginaceae). Am. J. Bot. 2011, 98, 1191–1200. [Google Scholar] [CrossRef]
- Skogsmyr, I.O.; Lankinen, Å. Sexual selection: An evolutionary force in plants? Biol. Rev. 2002, 77, 537–562. [Google Scholar] [CrossRef]
- Marshall, D.L.; Ellstrand, N.C. Proximal causes of multiple paternity in wild radish, Raphanus sativus. Am. Nat. 1985, 126, 596–605. [Google Scholar] [CrossRef]
- Dini-Papanastasi, O.; Aravanopoulos, F.A. Artificial hybridization between Robinia pseudoacacia L. and R. pseudoacacia var. monophylla Carr. Forestry 2008, 81, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan, D.I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2006, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Aizen, M.A.; Morales, C.L.; Morales, J.M. Invasive mutualists erode native pollination webs. PLoS Biol. 2008, 6, e31. [Google Scholar] [CrossRef]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. Long-term global trends in crop yield and production reveal no current pollination shortage but increasing pollinator dependency. Curr. Biol. 2008, 18, 1572–1575. [Google Scholar] [CrossRef] [Green Version]
- Chu, Z.Q.; Wang, B.; Luo, Y.G. Research on increasing yield by honeybee pollination of Helianthus annuus L. in Ningxia. Apic. China 2014, 65, 29–31. [Google Scholar]
- Ge, F.C.; Li, Y.F.; Yan, D.B.; Song, Y.M.; Zhao, L.M.; Sun, H.; Yao, Z.L. Report of improving soybean yield in field by using honeybee as pollinator. Apic. China 2012, 9, 24–26. [Google Scholar]
- Shi, Y.Y.; Guan, C.; Zeng, Z.J.; An, J.D.; Luo, S.D. Yield-increasing effect mechanism of honeybee on rape pollination. Acta Agric. Univ. Jiangxiensis 2009, 31, 994–999+1005. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Female Parents (♀) | Male Parents (♂) | Mating Combinations |
|---|---|---|---|
| 2016 | 2016-C1 | 2016-C1 + 2016-C4 + 2016-C5 | 2016-C1 × (2016-C1 + 2016-C4 + 2016-C5) |
| 2016-C2 | 2016-C2 + 2016-C4 + 2016-C5 | 2016-C2 × (2016-C2 + 2016-C4 + 2016-C5) | |
| 2016-C3 | 2016-C3 + 2016-C4 + 2016-C5 | 2016-C3 × (2016-C3 + 2016-C4 + 2016-C5) | |
| 2017 | 2017-C1 | 2017-♂1 + 2017-♂2 + 2017-♂3 | 2017-C1 × (2017-♂1 + 2017-♂2 + 2017-♂3) |
| 2017-C2 | 2017-C2 × (2017-♂1 + 2017-♂2 + 2017-♂3) | ||
| 2017-C3 | 2017-C3 × (2017-♂1 + 2017-♂2 + 2017-♂3) | ||
| 2017-C4 | 2017-♂4 + 2017-♂5 + 2017-♂6 | 2017-C4 × (2017-♂4 + 2017-♂5 + 2017-♂6) | |
| 2017-C5 | 2017-C5 × (2017-♂4 + 2017-♂5 + 2017-♂6) | ||
| 2017-C6 | 2017-C6 × (2017-♂4 + 2017-♂5 + 2017-♂6) | ||
| 2017-C7 | 2017-♂7 + 2017-♂8+2017-♂9 | 2017-C7 × (2017-♂7 + 2017-♂8 + 2017-♂9) | |
| 2017-C8 | 2017-C8 × (2017-♂7 + 2017-♂8 + 2017-♂9) | ||
| 2017-C9 | 2017-C9 × (2017-♂7 + 2017-♂8 + 2017-♂9) | ||
| 2018 | 2018-C1 | 2018-♂4 | 2018-C1 × 2018-♂4 |
| 2018-C4 | 2018-C4 × 2018-♂4 | ||
| 2018-C5 | 2018-C5 × 2018-♂4 | ||
| 2018-C2 | 2018-♂2 | 2018-C2 × 2018-♂2 | |
| 2018-C3 | 2018-C3 × 2018-♂2 | ||
| 2018-C6 | 2018-♂1 + 2018-♂3 + 2018-♂5 | 2018-C6 × (2018-♂1 + 2018-♂3 + 2018-♂5) | |
| 2018-C7 | 2018-C7 × (2018-♂1 + 2018-♂3 + 2018-♂5) | ||
| 2018-C8 | 2018-C8 × (2018-♂1 + 2018-♂3 + 2018-♂5) |
| Locus | Repeat Motif | Primer Sequences (5’ to 3’) | Size (bp) | Source |
|---|---|---|---|---|
| Rply3 | (GTGGTA)4 | F: GCCTCATAAATAAAAGGAACG | 138–164 | Our group |
| R: CTGCCATTGGTAACTGGTAAA | ||||
| Rply5 | (ATG)8 | F: GAGTCATGCCCTTTGTATGTT | 122–143 | Our group |
| R: TGTCACCTTCAAGTCCCTATT | ||||
| Rply8 | (CT)14 | F: TCCCTACATAAAACTCCAAA | 239–271 | Our group |
| R: TCATTAAGTCAGCACTCACAG | ||||
| Rply49 | (CT)8 | F: CCCCGTACAGTTCCATCT | 118–144 | Our group |
| R: GACCTCGTAAAAGCCACC | ||||
| Rops08 | (CA)8TA(CA)3 | F: TTCTGAGGAAGGGTTCCGTGG | 191–205 | Lian et al., 2002 |
| R: GTTAAAGCAACAGGCACATGG | ||||
| Rp109 | (AG)17 | F: GAGGAATCACAAAACCGTTTGG | 136–160 | Mishima et al., 2009 |
| R: TGGGATTTGAGAGAGTGGTGGTG | ||||
| Rp200 | (AG)23 | F: GGTTTCTTTGTTCACCTGCTCTGG | 160–198 | Mishima et al., 2009 |
| R: ACCTACGTGTCCACGGCTCT | ||||
| Rp206 | (GT)9 | F: GCCAAATCCCATTAGATCACAGTTGA | 222–246 | Mishima et al., 2009 |
| R: AGAAGTTAGACTTACGTGCTGC |
| Stages | Number of Samples | Correlation | Genetic Similarity | Hybrid Affinity |
|---|---|---|---|---|
| 7 days after pollination | 69 | Pearson correlation | 1 | 0.272 * |
| Significance (two-tailed test) | 0.024 | |||
| 30 days after pollination | 69 | Pearson correlation | 1 | 0.676 ** |
| Significance (two-tailed test) | 0.000 | |||
| Seed maturity | 36 | Pearson correlation | 1 | 0.446 ** |
| Significance (two-tailed test) | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Hu, R.; Dong, L.; Li, X.; Zhang, Z.; Guo, Q.; Cao, S.; Li, J.; Han, P.; Han, C.; et al. Pollen Competition and Paternal Contribution during Artificially Controlled Pollination of Black Locust (Robinia pseudoacacia L.) without Castration. Forests 2021, 12, 1416. https://doi.org/10.3390/f12101416
Sun Y, Hu R, Dong L, Li X, Zhang Z, Guo Q, Cao S, Li J, Han P, Han C, et al. Pollen Competition and Paternal Contribution during Artificially Controlled Pollination of Black Locust (Robinia pseudoacacia L.) without Castration. Forests. 2021; 12(10):1416. https://doi.org/10.3390/f12101416
Chicago/Turabian StyleSun, Yuhan, Ruiyang Hu, Li Dong, Xiuyu Li, Zijie Zhang, Qi Guo, Sen Cao, Jiankang Li, Peiyao Han, Chao Han, and et al. 2021. "Pollen Competition and Paternal Contribution during Artificially Controlled Pollination of Black Locust (Robinia pseudoacacia L.) without Castration" Forests 12, no. 10: 1416. https://doi.org/10.3390/f12101416
APA StyleSun, Y., Hu, R., Dong, L., Li, X., Zhang, Z., Guo, Q., Cao, S., Li, J., Han, P., Han, C., Uddin, S., Long, C., Fan, Y., & Li, Y. (2021). Pollen Competition and Paternal Contribution during Artificially Controlled Pollination of Black Locust (Robinia pseudoacacia L.) without Castration. Forests, 12(10), 1416. https://doi.org/10.3390/f12101416