
Opposite Tree-Tree Interactions Jointly Drive the Natural Fir Treeline Population on the Southeastern Tibetan Plateau



Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area and Climate
2.2. Tree Species and Field Sampling
2.3. Data Analysis
2.3.1. Analysis of Population Traits
2.3.2. Surface Pattern Analysis
2.3.3. Spatial Point Pattern Analysis
2.3.4. Comparative Analysis
3. Results
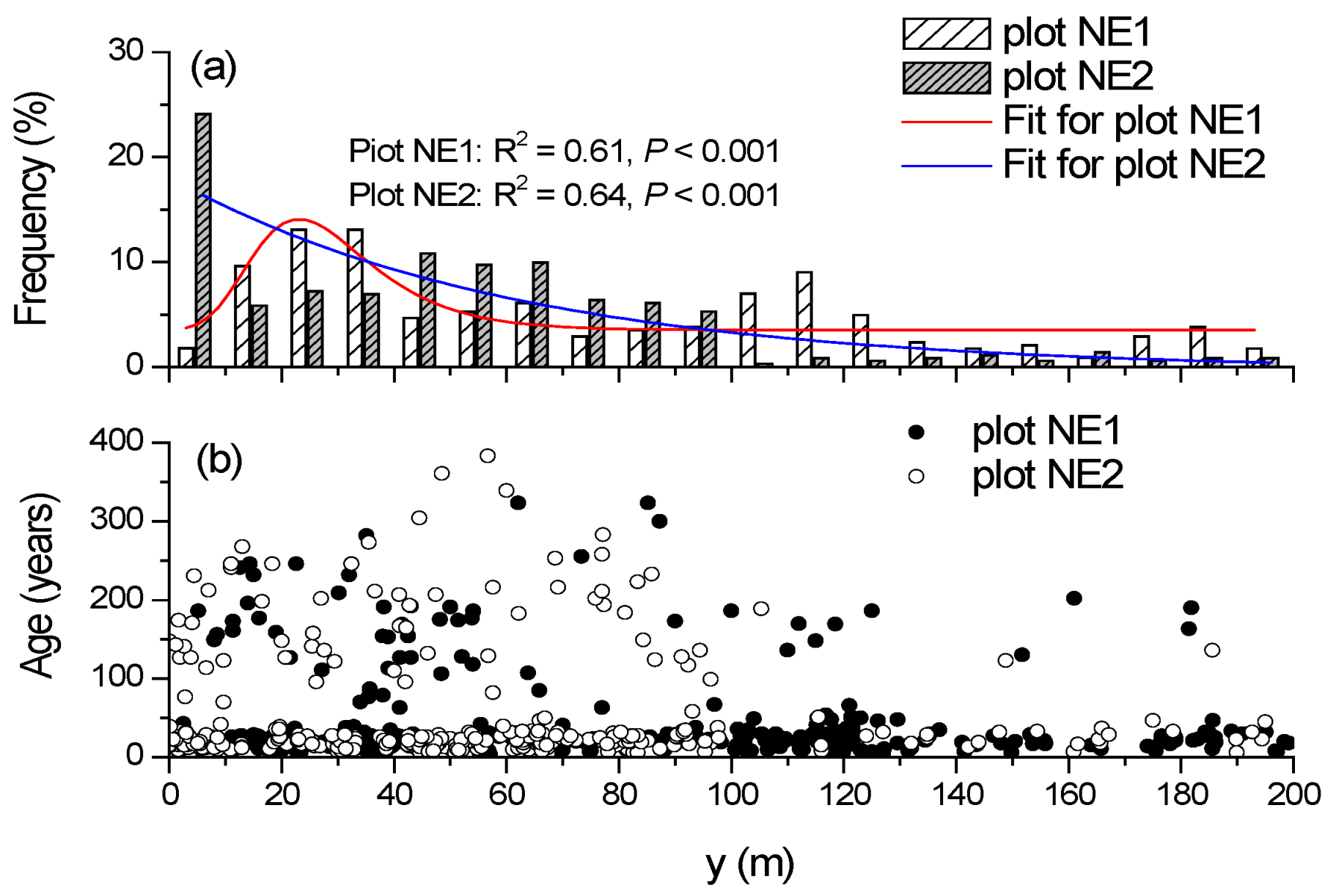
3.1. Trait Relationships and Population Structure
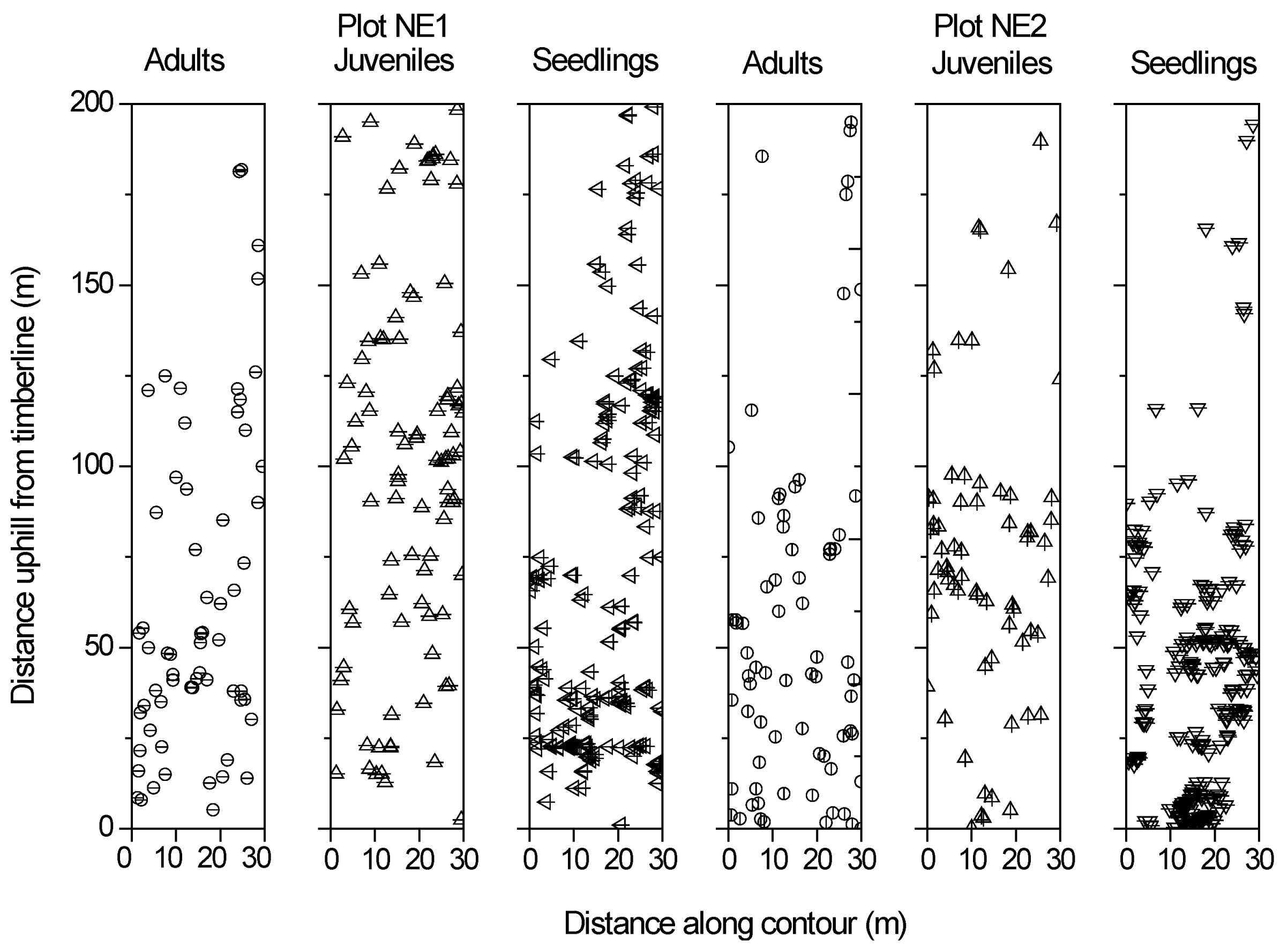
3.2. Relationships between Tree Age and Altitude
3.3. Spatial Autocorrelation of Trees
3.4. Spatial Distribution Patterns of Tree Size Classes
3.5. Spatial Associations between Different Tree Size Classes
3.6. Tree Spatial Patterns in Other Treeline Sites on the TP
4. Discussion
4.1. Stand Structure and Tree Trait Relations
4.2. Tree Spatial Patterns
4.3. Comparisons with Previous Studies on the TP
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McIntire, J.B.; Fajardo, A. Beyond Description: The Active and Effective Way to Infer Processes from Spatial Patterns. Ecology 2009, 90, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Lyu, L.X.; Zhang, Q.B.; Deng, X.; Mäkinen, H. Fine-scale distribution of treeline trees and the nurse plant facilitation on the eastern Tibetan Plateau. Ecol. Indic. 2016, 66, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, M.; Kruse, S.; Epp, L.S.; Kolmogorov, A.; Nikolaev, A.N.; Heinrich, I.; Jeltsch, F.; Pestryakova, L.A.; Zibulski, R. Dissimilar responses of larch stands in northern Siberia to increasing temperatures-a field and simulation based study. Ecology 2017, 98, 2343–2355. [Google Scholar] [CrossRef] [Green Version]
- Holtmeier, F.K.; Broll, G. Treeline advance-driving processes and adverse factors. Landsc. Online 2007, 1, 1–33. [Google Scholar] [CrossRef]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T.; et al. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dynam. 2016, 6, 245–265. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Q.; Xie, F.L.; He, N.J.; He, R.; Zhang, K.R.; Zhang, Q.F.; Dang, H.S. Disparity in elevational shifts of upper species limits in response to recent climate warming in the Qinling Mountains, North-central China. Sci. Total Environ. 2020, 706, 135718. [Google Scholar] [CrossRef]
- Sigdel, S.R.; Liang, E.; Wang, Y.F.; Dawadi, B.; Camarero, J.J. Tree-to-tree interactions slow down Himalayan treeline shifts as inferred from tree spatial patterns. J. Biogeogr. 2020, 47, 1816–1826. [Google Scholar] [CrossRef]
- Astudillo-Sanchez, C.C.; Fowler, M.S.; Villanueva-Diaz, J.; Villanueva-Díaz, J.; Endara-Agramont, A.R.; Soria-Díaz, L. Recruitment and facilitation in Pinus hartwegii, a Mexican alpine treeline ecotone, with potential responses to climate warming. Trees 2019, 33, 1087–1100. [Google Scholar] [CrossRef] [Green Version]
- Elliot, G.P. Influences of 20th-century warming at the upper tree line contingent on local-scale interactions: Evidence from a latitudinal gradient in the Rocky Mountains, USA. Glob. Ecol. Biogeogr. 2011, 20, 46–57. [Google Scholar] [CrossRef]
- Wang, Y.; Pederson, N.; Ellison, A.M.; Buckley, H.L.; Case, B.S.; Liang, E.Y.; Julio, C.; Camarero, J.J. Increased stem density and competition may diminish the positive effects of warming at alpine treeline. Ecology 2016, 97, 1668–1679. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Scherrer, D.; Vitasse, Y.; Guisan, A.; Wohlgemuth, T.; Lischke, H. Competition and demography rather than dispersal limitation slow down upward shifts of trees’s upper elevation limits in the Alps. J. Ecol. 2020, 108, 2416–2430. [Google Scholar] [CrossRef]
- Liu, B.; Liang, E.; Zhu, L.P. Microclimatic conditions for Juniperus saltuaria treeline in the Sygera Mountains, southeastern Tibetan Plateau. Mt. Res. Dev. 2011, 31, 45–53. [Google Scholar] [CrossRef]
- Li, X.; Liang, E.; Gričar, J.; Rossi, S.; Čufaret, K.; Ellison, A.M. Critical temperature and precipitation thresholds for the onset of xylogenesis and maintains treelines on the southeastern Tibetan Plateau. Sci. Bull. 2017, 62, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Arekhi, M.; Yesil, A.; Ozkan, Y.; Sanli, F.B. Detecting treeline dynamics in response to climate warming using forest stand maps and Landsat data in a temperate forest. For. Ecosyst. 2018, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Gatti, R.C.; Callaghan, T.; Velichevskaya, A.; Dudko, A.; Fabbio, L.; Battipaglia, G.; Liang, J. Accelerating upward treeline shift in the Altai Mountains under last-century climate change. Sci. Rep. 2019, 9, 768. [Google Scholar]
- Lu, X.; Liang, E.; Wang, Y.; Babst, F.; Camarero, J.J. Mountain treelines climb slowly despite rapid climate warming. Glob. Ecol. Biogeogr. 2021, 30, 305–315. [Google Scholar] [CrossRef]
- Batllori, E.; Gutierrez, E. Regional tree line dynamics in response to global change in the Pyrenees. J. Ecol. 2008, 96, 1275–1288. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gutierrez, E. Pace and pattern of recent treeline dynamics: Response of ecotones to climatic variability in the Spanish Pyrenees. Clim. Chang. 2004, 63, 181–200. [Google Scholar] [CrossRef]
- Bailey, S.N.; Elliot, G.P.; Schliep, E.M. Seasonal temperature-moisture interactions limit seedling establishment at upper treeline in the Southern Rockies. Ecosphere 2021, 12, e03568. [Google Scholar] [CrossRef]
- Sigdel, S.R.; Wang, Y.; Camarero, J.J.; Zhu, H.; Liang, E.; Peuelas, J. Moisture-mediated responsiveness of treeline shifts to global warming in the Himalayas. Glob. Chang. Bio 2018, 24, 5549–5559. [Google Scholar] [CrossRef] [Green Version]
- Harper, K.A.; Danby, R.K.; From Fields, D.L.; Lewis, K.P.; Trant, A.J.; Starzomski, B.M.; Savidge, R.; Hermanutz, L. Tree spatial pattern within the forest-tundra ecotone: A comparison of sites across Canada. Can. J. For. Res. 2011, 41, 479–489. [Google Scholar] [CrossRef]
- Feuillet, T.; Birre, D.; Milian, J.; Godard, V.; Clauzel, C.; Serrano-Notivoli, R. Spatial dynamics of alpine tree lines under global warming: What explains the mismatch between tree densification and elevational upward shifts at the tree line ecotone? J. Biogeogr. 2020, 47, 1056–1068. [Google Scholar] [CrossRef]
- Callaway, R.M. Positive Interactions and Interdependence in Plant Communities; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Smith, W.K.; Germino, M.J.; Hancock, T.E.; Johnson, D.M. Another perspective on altitudinal limits of alpine timberlines. Tree Physiol. 2003, 23, 1101–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miehe, G.; Miehe, S.; Vogel, J.; Co, s.; Duo, L. Highest treeline in the northern hemisphere found in southern Tibet. Mt. Res. Dev. 2007, 27, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Wang, Y.; Piao, S.; Lu, X.; Camarero, J.J.; Zhu, L.; Zhu, L.; Ellison, A.M.; Ciais, P.; Josep, P. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2016, 113, 4380–4385. [Google Scholar] [CrossRef] [Green Version]
- Miao, N.; Liu, S.; Shi, Z.; Xue, Z.B.; Yu, H.; He, F. Spatial pattern analysis of a Rhododendron-Abies virginal forest near timberline on the eastern edge of Qinghai-Tibetan Plateau, China. Acta Ecol. Sin. 2011, 31, 1–9. [Google Scholar]
- Wang, Y.; Camarero, J.J.; Luo, T.; Liang, E. Spatial patterns of Smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns. Plant Ecol. Divers. 2012, 5, 311–321. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, B.; Wang, Y. Size structure and spatial patterns of Balfour spruce treeline population in Yushu, Qinghai. J. Earth Environ. 2020, 11, 660–671. [Google Scholar] [CrossRef]
- Zhang, Q.; Luo, P.; Zhang, Y.; Shi, F.; Wu, N. Ecological characteristics of Abies georgi population at timberline on the north-facing slope of Baima Snow mountains, southwest China. Acta Ecol. Sin. 2008, 28, 129–135. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Luo, P.; Wang, Q.; Wu, N. Ecological characteristics of a Sabina saltuaria population at timberline on the south facing slope of Baima Snow Mountain, southwest China. J. Plant Ecol. 2007, 31, 857–864. [Google Scholar] [CrossRef]
- Bader, M.Y.; Llambi, L.D.; Case, B.S.; Buckley, H.; Toivon, J.M.; Camarero, J.J.; Cairns, D.M.; Brown, C.D.; Wiegand, T.; Resler, L.M. A global framework for linking alpine-treeline ecotone patterns to underlying processes. Ecography 2021, 44, 265–292. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations- the CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.F.; Shao, X.M.; Yin, Z.Y.; Xu, P.; Tian, H. August temperature variability in the southeastern Tibetan Plateau since AD 1385 inferred from tree rings. Palaeogeogr. Palaeoclim. Palaeocl 2011, 305, 84–92. [Google Scholar] [CrossRef]
- Liang, E.; Wang, Y.; Xu, Y.; Liu, B.; Shao, X. Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan Plateau. Trees 2010, 24, 363–373. [Google Scholar] [CrossRef]
- Liang, E.; Wang, F.; Eckstein, D.; Luo, T. Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytol. 2011, 190, 760–769. [Google Scholar] [CrossRef]
- Applequist, M.B. A simple pith locator for use with off-center increment cores. J. For. 1958, 56, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Gutiérrez, E.; Fortin, M. Spatial pattern of subalpine forest-alpine grassland ecotones in the Spanish Central Pyrenees. For. Ecol. Manag. 2000, 134, 1–16. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gutiérrez, E.; Fortin, M.; Ribbens, E. Spatial patterns of tree recruitment in a relict population of Pinus uncinata: Forest expansion through stratified diffusion. J. Biogeogr. 2005, 32, 1979–1992. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Peng, S.; Yirdaw, E.; Wu, N. Spatial structure of alpine trees in Mountain Baima Xueshan. on the southeast Tibetan Plateau. Silva Fenn. 2009, 43, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Batllori, E.; Camarero, J.J.; Gutiérrez, E. Current regeneration patterns at the tree line in the Pyrenees indicate similar recruitment processes irrespective of the past disturbance regime. J. Biogeogr. 2010, 37, 1938–1950. [Google Scholar] [CrossRef]
- Kenkel, N.C. Pattern of self-thinning in Jack pine: Testing the random mortality hypothesis. Ecology 1988, 69, 1017–1024. [Google Scholar] [CrossRef]
- Rietkerk, M.; Van de Koppel, J. Regular pattern formation in real ecosystems. Trends Ecol. Evol. 2008, 23, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Batllori, E.; Camarero, J.J.; Ninot, J.M.; Gutiérrez, E. Seedling recruitment, survival and facilitation in alpine Pinus uncinata tree line ecotones. Implications and potential responses to climate warming. Glob. Ecol. Biogeogr. 2009, 18, 460–472. [Google Scholar] [CrossRef]
- He, Q.; Bertness, M.D.; Altieri, A.H.; Monstserrat, V. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sylvester, S.P.; Lu, X.; Dawadi, B.; Sigdel, S.R.; Liang, E.; Camarero, J.J. The stability of spruce treelines on the eastern Tibetan Plateau over the last century is explained by pastoral disturbance. For. Ecol. Manag. 2019, 442, 34–45. [Google Scholar] [CrossRef]
- Naudiyal, N.; Wang, J.N.; Ning, W.; Gaire, N.P.; Peili, S.; Wei, Y.D.; He, J.A.; Shi, N.A. Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecol. Indic. 2021, 121, 107131. [Google Scholar] [CrossRef]
- Dai, Y.; Yao, T.; Li, X.; Ping, F. The impact of lake effect on the temporal and spatial distribution of precipitation in the Nam Co basin, Tibetan Plateau. Quat. Int. 2018, 475, 63–69. [Google Scholar] [CrossRef]
- Johnson, A.C.; Yeakley, J.A. Microsites and Climate Zones: Seedling Regeneration in the Alpine Treeline Ecotone Worldwide. Forests 2019, 10, 864. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liang, E.; Sigdel, S.R.; Liu, B.; Camarero, J. The coupling of treeline elevation and temperature is mediated by non-thermal factors on the Tibetan Plateau. Forests 2017, 8, 109. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Region | Species | EC | Spatial Distributions of Tree Classes (Scale: m) | Spatial Associations between Tree Classes (Scale: m) | Data Source | ||||
|---|---|---|---|---|---|---|---|---|---|
| SE | JV | AD | SE-JV | SE-AD | JV-AD | ||||
| AB, ETP | S1 | SHT | CD: 1–5 | CD: 1–2 | RGD: 1–2; RD: 3–15 | PA: 1–5 | NA: 1–6; PA: 8–13 | NA: 1–15 | [28] |
| YS, ETP | S2 | DP | CD: 1–4 | RD: 1–15 | RD: 1–15 | PA: 2–6 | PA: 9–14 | UC: 1–15 | [30] |
| CD, ETP | S2 | DS | CD: 1–15 | CD: 2–15 | RD: 2–15 | PA: 1–15 | PA: 1–15 | PA: 1–15 | [2] |
| LZ, SETP | S3 | SHT | CD: 1–7 | CD: 1–7 | RD: 1–15 | PA: 1–7 | NA: 1–5 | NA: 1–5 | [29] |
| DQ, SETP | S4 | WS | CD: 1–15 | CD: 1–15 | CD: 1–15 | PA: 1–15 | PA: 1–15 | PA: 1–15 | [31] |
| DQ, SETP | S5 | WS | RD: 1–15 | CD: 1–15 | CD: 1–15 | PA: 1–15 | PA: 1–15 | PA: 1–15 | [32] |
| Plot NE1 in RW, SETP | S3 | SHT | CD: 1–5 | CD: 1–2 | RD: 1–15 | PA: 1–4 | NA: 1–2, 10 | NA: 8–9, 12, 14 | This study |
| Plot NE2 in RW, SETP | S3 | SHT | CD: 1–7 | CD: 1–2 | RD: 1–15 | PA: 1–4; NA: 7–15 | PA: 7–14 | NA: 3–4, 6–8 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Mao, Q.; Ren, P.; Sigdel, S.R. Opposite Tree-Tree Interactions Jointly Drive the Natural Fir Treeline Population on the Southeastern Tibetan Plateau. Forests 2021, 12, 1417. https://doi.org/10.3390/f12101417
Wang Y, Mao Q, Ren P, Sigdel SR. Opposite Tree-Tree Interactions Jointly Drive the Natural Fir Treeline Population on the Southeastern Tibetan Plateau. Forests. 2021; 12(10):1417. https://doi.org/10.3390/f12101417
Chicago/Turabian StyleWang, Yafeng, Qing Mao, Ping Ren, and Shalik Ram Sigdel. 2021. "Opposite Tree-Tree Interactions Jointly Drive the Natural Fir Treeline Population on the Southeastern Tibetan Plateau" Forests 12, no. 10: 1417. https://doi.org/10.3390/f12101417
APA StyleWang, Y., Mao, Q., Ren, P., & Sigdel, S. R. (2021). Opposite Tree-Tree Interactions Jointly Drive the Natural Fir Treeline Population on the Southeastern Tibetan Plateau. Forests, 12(10), 1417. https://doi.org/10.3390/f12101417