
Effects of Temperature, Scarification, Stratification, Phytohormones, and After-Ripening on the Dormancy and Germination of Eucommia ulmoides Oliv. Seeds


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seeds
2.2. Determination of the Moisture Content
2.3. Determination of the Seed Viability
2.4. Germination of the Isolated Embryos
2.5. Water Absorption by the Intact Seeds, Scarified Seeds, and Isolated Embryos
2.6. After-Ripening Treatment
2.7. Scarification Treatment
2.8. Stratification Treatment
2.9. GA3 Treatment
2.10. Germination Test
2.11. Statistical Analysis
3. Results
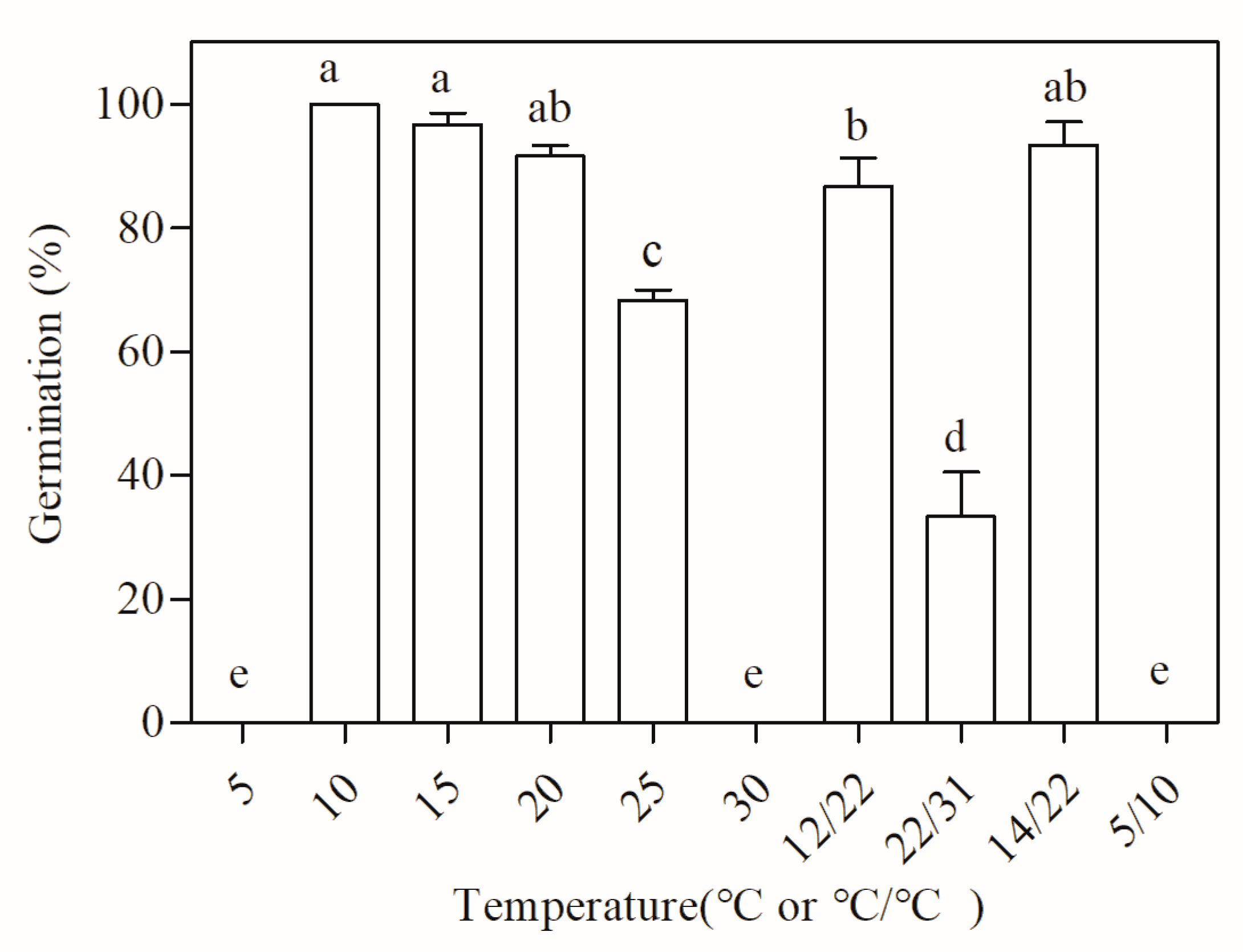
3.1. Viability and Germination of the Intact Seeds
3.2. Germination of the Isolated Embryos
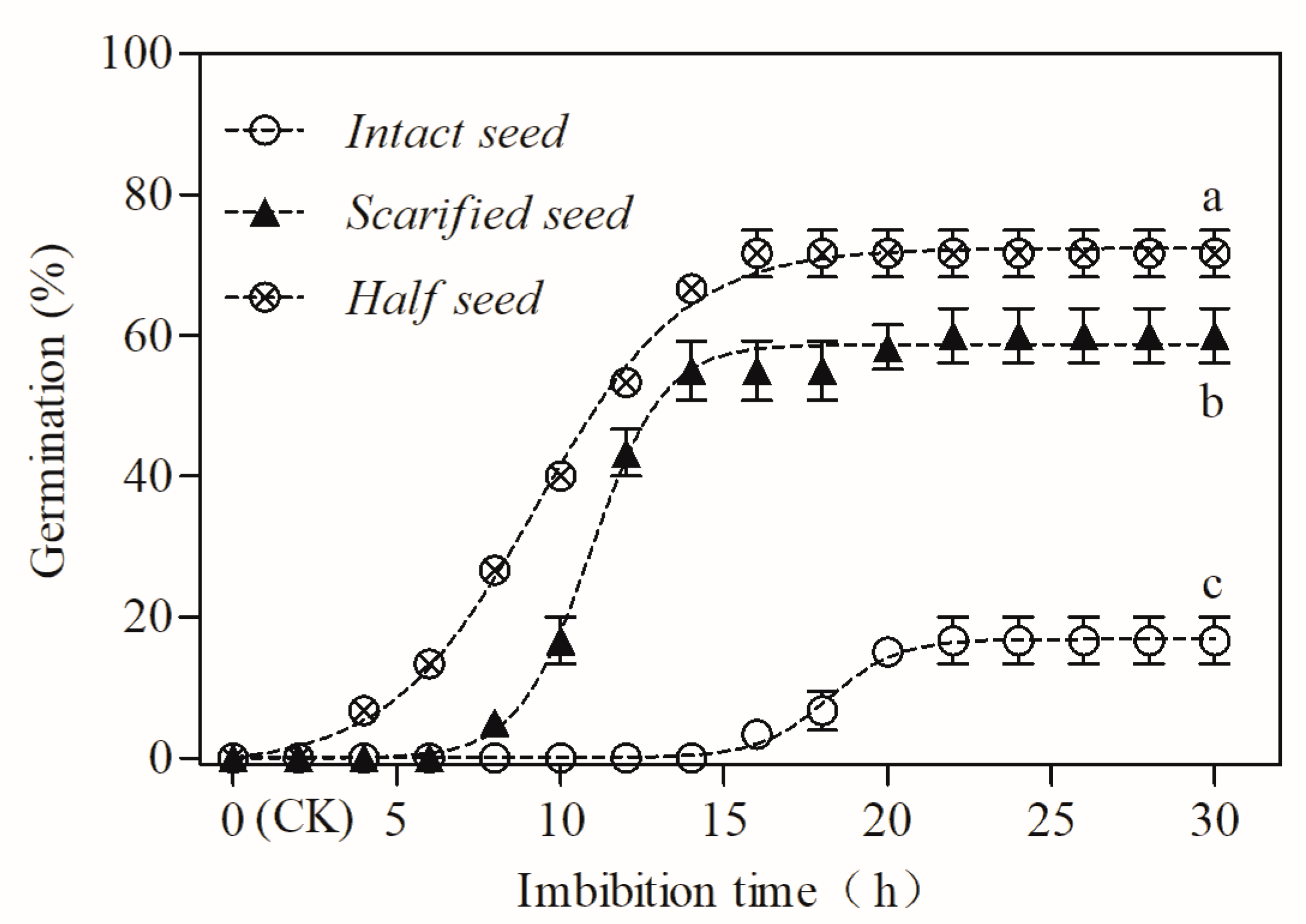
3.3. Effect of Scarification on the Dormancy Release and Germination
3.4. Water Absorption by the Intact Seeds, Scarified Seeds, and Isolated Embryos
3.5. Effects of Stratification on the Dormancy Release and Germination
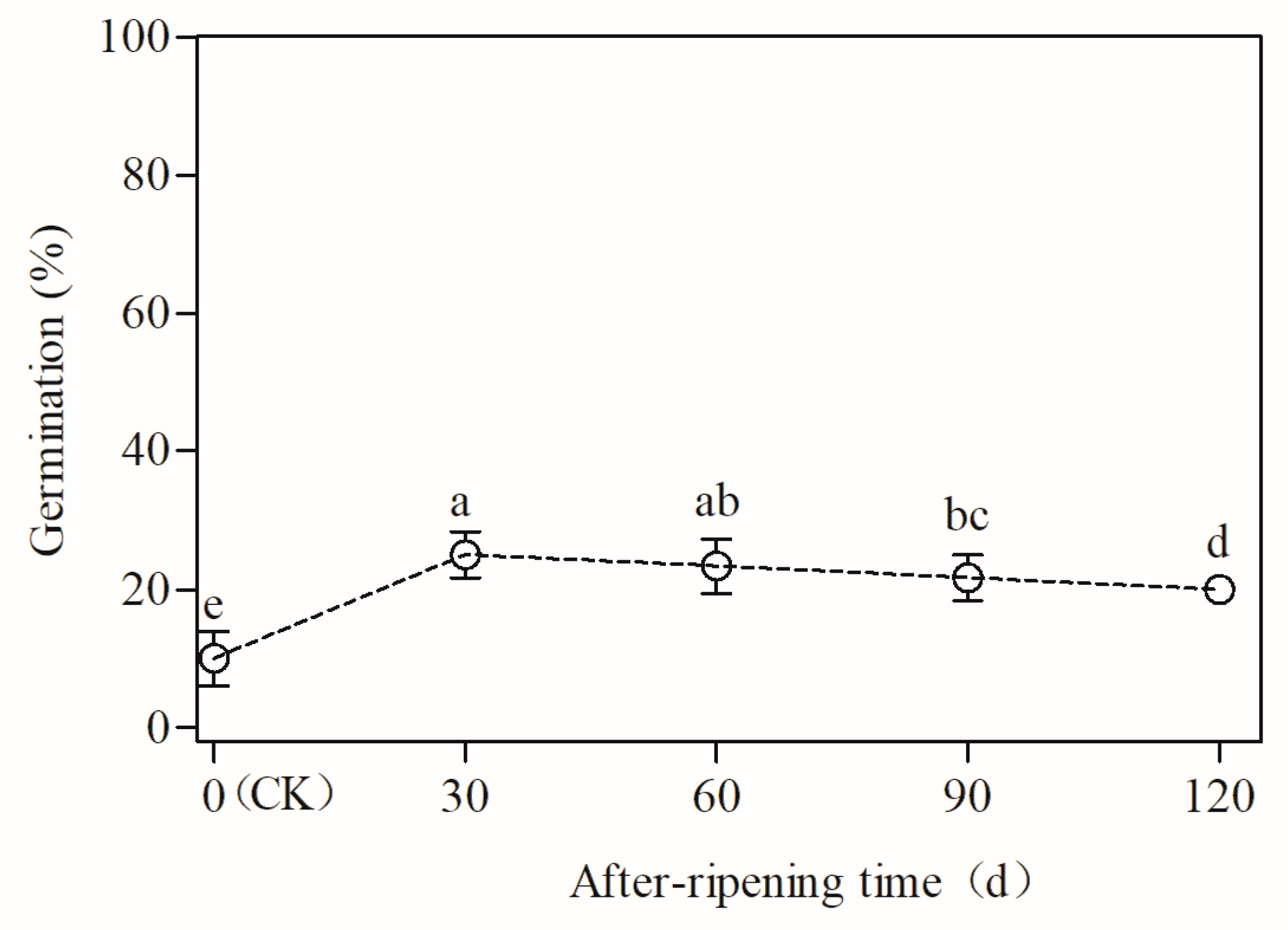
3.6. Effect of After-Ripening on the Dormancy Release and Germination
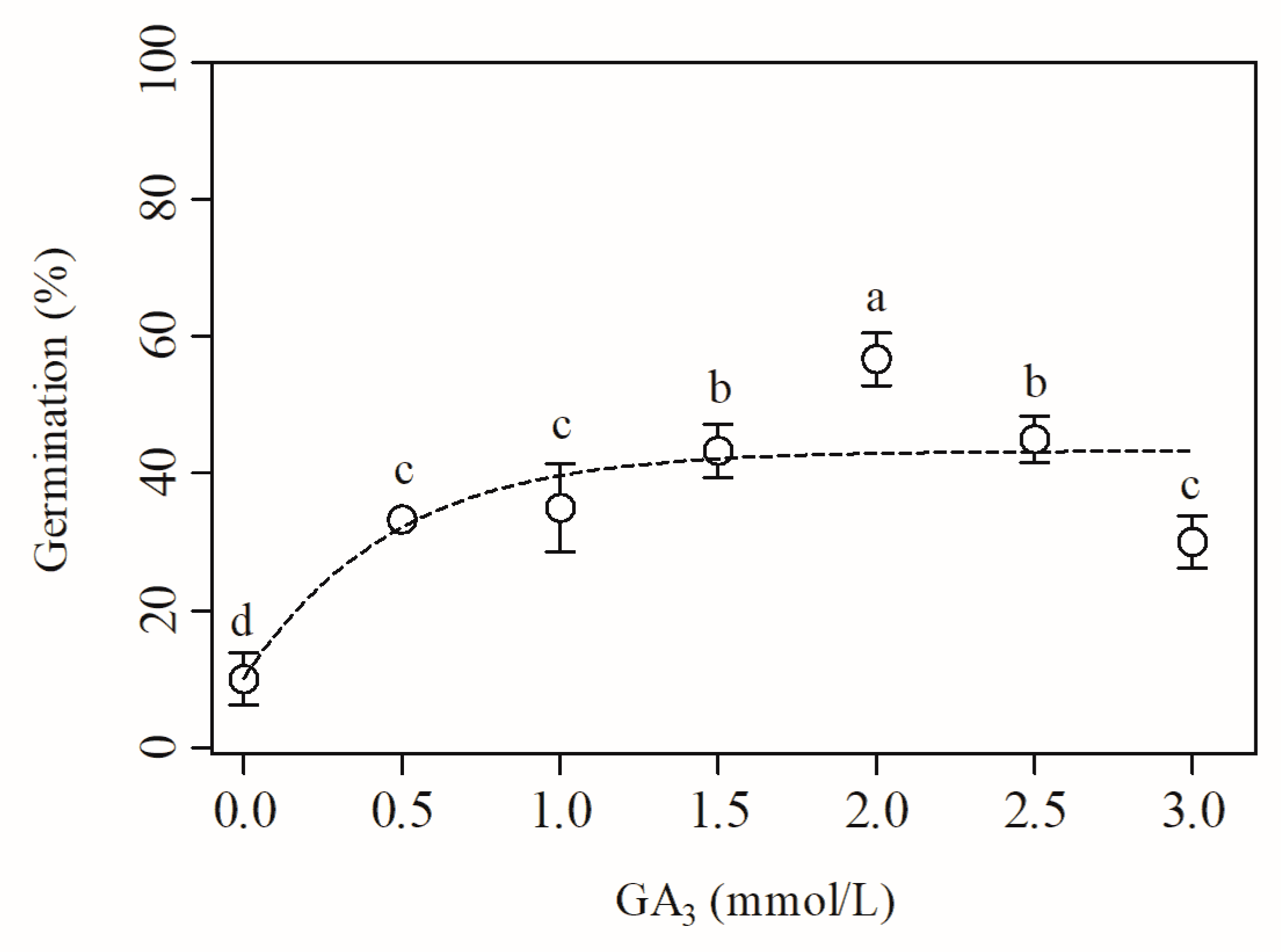
3.7. Effect of GA3 Treatment on the Dormancy Release and Germination
4. Discussion
4.1. Determination of the Dormancy Type
4.2. Effective Methods for Releasing Dormancy
4.3. Seed Germination and Phylogeny of E. ulmoides
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bewley, J.D. Seed Germination and Dormancy. Plant Cell 1997, 9, 1055–1066. Available online: https://academic.oup.com/plcell/article/9/7/1055/5986415 (accessed on 18 October 2021). [CrossRef] [PubMed] [Green Version]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. Available online: http://www.seedbiology.eu/pdf/NPH-TR-2006-04168_applied.pdf (accessed on 18 October 2021). [CrossRef] [PubMed]
- Bradford, K.; Nonogaki, H. Seed Development, Dormancy and Germination; Blackwell Publishing Press: Oxford, UK, 2007; Available online: https://lib.ugent.be/catalog/rug01:001223298 (accessed on 18 October 2021).
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2014; Available online: http://www.irgrid.ac.cn/handle/1471x/915815 (accessed on 18 October 2021).
- Nikolaeva, M. On criteria to use in studies of seed evolution. Seed Sci. Res. 2004, 14, 315–320. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998; Available online: https://www.sciencedirect.com/science/article/abs/pii/S017616179980159X (accessed on 18 October 2021).
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Curr. Opin. Plant Biol. 2002, 5, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Blanco, C.; Bentsink, L.; Hanhart, C.J.; Vries, H.B.D.; Koornneef, M. Analysis of natural allelic variation at seed dormancy loci of Arabidopsis thaliana. Genetics 2003, 164, 711–729. Available online: https://www.genetics.org/content/164/2/711.abstract (accessed on 18 October 2021). [CrossRef]
- Jing, Y.; Liu, W.; Zhang, Y.H. Seed germination characteristics of Spartina alterniflora from high and low latitude populations in relation to temperature. Chin. J. Ecol. 2021, 40, 2763–2772. [Google Scholar] [CrossRef]
- Ņečajeva, J.; Bleidere, M.; Jansone, Z.; Gailīte, A.; Ruņģis, D. Variability of seed germination and dormancy characteristics and genetic analysis of Latvian Avena fatua populations. Plants 2021, 10, 235. Available online: https://www.researchgate.net/publication/348803707_Variability_of_Seed_Germination_and_Dormancy_Characteristics_and_Genetic_Analysis_of_Latvian_Avena_fatua_Populations (accessed on 18 October 2021). [CrossRef]
- Huang, Z.; Ölçer-Footitt, H.; Footitt, S.; Finch-Savage, W.E. Seed dormancy is a dynamic state: Variable responses to pre- and post-shedding environmental signals in seeds of contrasting Arabidopsis ecotypes. Seed Sci. Res. 2015, 25, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Cheng, J.; Wang, J.; Cheng, Y.; He, Y.; Zhang, H.; Wang, Z. Physiological characteristics of cold stratification on seed dormancy release in rice. Plant Growth Regul. 2019, 89, 131–141. [Google Scholar] [CrossRef]
- Benjelloun, J.; Bouzroud, S.; Triqui, Z.E.A.; Alami, Q.L.; Layachi, R.; Smouni, A.; Guedira, A. Warm stratification improves embryos development and seed germination of Cycas revoluta. Adv. Hortic. Sci. 2021, 35, 91–96. [Google Scholar] [CrossRef]
- Xue, T.T.; She, Y.B.; Liu, J.; Shi, F. Advances in research of seed physical dormancy. Chin. Bull. Bot. 2016, 51, 863–871. [Google Scholar] [CrossRef]
- Nawrot-Chorabik, K.; Osmenda, M.; Słowiński, K.; Latowski, D.; Tabor, S.; Woodward, S. Stratification, Scarification and Application of Phytohormones Promote Dormancy Breaking and Germination of Pelleted Scots Pine (Pinus sylvestris L.) Seeds. Forest 2021, 12, 621. [Google Scholar] [CrossRef]
- Liang, J.; Li, X.; Liu, J.; Zhang, Q.; Song, S.; Xu, H.; Huang, H.; Wu, X. Gibberellin metabolism and signaling and its molecular mechanism in regulating seed germination and dormancy. Sci. Sin. Vitae 2020, 50, 599–615. [Google Scholar] [CrossRef]
- Song, S.Q.; Liu, J.; Xu, H.H.; Liu, X.; Huang, H. ABA metabolism and signaling and their molecular mechanism regulating seed dormancy and germination. Sci. Agric. Sin. 2020, 50, 599–615. [Google Scholar] [CrossRef]
- Holdsworth, M.J.; Bentsink, L.; Soppe, W.J.J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytol. 2008, 179, 33–54. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Liu, S.; Yuan, G.; Liu, P.; Qi, D.; Dong, X.; Liu, H.; Liu, G.; Li, X. Germination characteristics among different sheepgrass (Leymus chinensis) germplasm during the seed development and after-ripening stages. PeerJ 2019, 7, e6688. [Google Scholar] [CrossRef]
- Feng, F.; Liang, Z.R. Understanding and utilization of Eucommia ulmoides in Chinese history. J. Northwest For. Coll. 1996, 11, 84–89. Available online: https://www.doc88.com/p-8199696804092.html?r=1 (accessed on 18 October 2021).
- Wang, D.W.; Wang, Y.C.; Li, G.Q.; Li, Z.Q. Research progress on breeding of Eucommia ulmoides Oliv. Jiangsu Agric. Sci. 2014, 42, 7–10. Available online: https://www.docin.com/p-1058899047.html (accessed on 18 October 2021).
- Wu, K.X.; Wu, B.; Peng, X.X.; Tang, Z.H.; Chen, L.W. Active constituents and eucommia gum content of Eucommia ulmoides Oliver from multiple origins. J. Northeast. For. Univ. 2019, 47, 40–43, 74. Available online: http://dlxb.nefu.edu.cn/#/digest?ArticleID=2700 (accessed on 18 October 2021).
- Fu, Y.C. The properties and uses of chinese gutia-percha. Polym. Mater. Sci. Eng. 1992, 21, 123–126. Available online: https://www.doc88.com/p-5731018625852.html (accessed on 18 October 2021).
- Wang, F.S.; Du, A.H.; Huang, B.C. Properties of TPI and its application in tire. China Rubber Ind. 2005, 12, 762–764. Available online: https://www.doc88.com/p-961176375661.html (accessed on 18 October 2021).
- Zhang, J.; Xue, Z. Study on under-water sound absorption properties of Eucommia ulmoides gum and its blends. Polym. Bull. 2011, 67, 511–525. [Google Scholar] [CrossRef]
- Zhong, R. Studies on seed germination of Eucommia ulmoides. J. Gansu For. Sci. Technol. 2018, 43, 13–15. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-GSLK201801005.html (accessed on 18 October 2021).
- Xu, B.M.; Bai, K.Z. Effects of low temperature on seed germination and seedling establishment of Eucommia ulmoides. For. Sci. Technol. 1993, 9, 5–6. Available online: http://lib.cqvip.com/Qikan/Article/Detail?id=1116328 (accessed on 18 October 2021).
- Cheng, J.F.; Tian, Q.J.; Wang, S.L.; He, X.B.; Peng, X.M. The effect of storage in sand and seeding time on seed germination and seedling growth of Eucommia ulmoides. Seed 2013, 32, 93–99. Available online: http://qikan.cqvip.com/Qikan/Article/Detail?id=46904537 (accessed on 18 October 2021).
- Ning, H.; Qi, Y.Y. Fast test of vitality and germination rate of Eucommia Seeds. J. Hubei Univ. Educ. 2009, 26, 44–46. [Google Scholar] [CrossRef]
- Moore, R.P. Tetrazolium staining for assessing seed quality. In Seed Ecology; Heydecker, W., Ed.; Butterworths: London, UK, 1973; pp. 347–366. Available online: http://citeseerx.ist.psu.edu/showciting?cid=31299499 (accessed on 18 October 2021).
- Li, T.H.; Min, X.H. Dormancy characteristics and germination requirements of Phoebe bournei seed. Sci. Hortic. 2020, 26, 108903. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 18 October 2021).
- Pipinis, E.; Stampoulidis, A.; Milios, E.; Kitikidou, K.; Radoglou, K. Effects of Cold Stratification and GA3 on Germination of Arbutus Unedo Seeds of Three Provenances. Afr. J. Tradit. Complement. Altern. Med. 2016, 14, 318–323. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, S.H.; Jia, N. Effects of different concentrations of gibberellin on seed germination in Eucommia ulmoide. J. Anhui Agric. Sci. 2019, 47, 144–146, 185. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-AHNY201903046.html (accessed on 18 October 2021).
- Yang, L.-E.; Peng, D.-L.; Li, Z.-M.; Huang, L.; Yang, J.; Sun, H. Cold stratification, temperature, light, GA3, and KNO3 effects on seed germination of Primula beesiana from Yunnan, China. Plant Divers. 2020, 42, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Forbis, T.A.; Floyd, S.K.; De Queiroz, A. The Evolution of Embryo Size in Angiosperms and Other Seed Plants: Implications for the Evolution of Seed Dormancy. Evolution 2002, 56, 2112–2125. [Google Scholar] [CrossRef]
- Van Assche, J.A.; Vandelook, F.E. Germination ecology of eleven species of Geraniaceae and Malvaceae, with special reference to the effects of drying seeds. Seed Sci. Res. 2006, 16, 283–290. Available online: http://www.editorialmanager.com/ssr/default.asp (accessed on 18 October 2021). [CrossRef]
- Owens, S.A.; Fields, P.F.; Ewers, F.W. Degradation of the upper pulvinus in modern and fossil leaves of Cercis (Fabaceae). Am. J. Bot. 1998, 85, 273–284. [Google Scholar] [CrossRef]
- Manchester, S.R. Inflorescence bracts of fossil and extant Tilia in North America, Europe, and Asia: Patterns of morphologic divergence and biogeographic history. Am. J. Bot. 1994, 81, 1176–1185. [Google Scholar] [CrossRef]
- Prothero, D.R. The Eocene-Oligocene Transition: Paradise Lost; Columbia University Press: New York, NY, USA, 1994; p. 291. Available online: https://xueshu.baidu.com/usercenter/paper/show?paperid=39d8c22a09d5e8c09a2e2aae89efc6d1&site=xueshu_se (accessed on 18 October 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Sum of Squares | Df | Mean Squares | f-Value | p-Value |
|---|---|---|---|---|---|
| Seed integrity | 43,575.729 | 3 | 21,787.87 | 992.60 | 0.00 |
| Temperature | 55,389.446 | 9 | 6154.387 | 280.38 | 0.00 |
| Seed integrity × temperature | 22,433.146 | 27 | 1246.29 | 56.78 | 0.00 |
| Stratification Temperature (°C) | Stratification Duration (Day) | Germination Percentage (%) | |||||
|---|---|---|---|---|---|---|---|
| 5 °C | 10 °C | 15 °C | 20 °C | 25 °C | 30 °C | ||
| 5 | 0 | 0.0 Dc | 13.3 ± 0.7 DEa | 5.0 ± 1.7 DEFb | 1.7 ± 1.7 Dc | 0.0 Cc | 0.0 Ac |
| 15 | 0.0 Db | 13.3 ± 0.7 Ea | 3.3 ± 3.3 EFb | 0.0 Db | 0 Cb | 0.0 Ab | |
| 30 | 0.0 Dd | 28.3 ± 6.9 Da | 18.3 ± 7.4 CDab | 10.0 ± 4.3 Cbc | 1.7 ± 1.7 BCcd | 0.0 Ad | |
| 45 | 0.0 Dd | 48.3 ± 9.2 Ca | 20 ± 3.9 Cb | 10.0 ± 3.3 Cbc | 5.0 ± 3.1 ABcd | 0.0 Ad | |
| 60 | 5.0 ± 1.67 Cb | 51.7 ± 5.0 Ca | 30.7 ± 5.7 CEb | 13.3 ± 2.7 Cb | 0.0 Cc | 0.0 Ac | |
| 75 | 6.6 ± 0.0 Cc | 72 ± 4.7 BCa | 58.3 ± 6.8 Ba | 35.0 ± 3.2 Bb | 8.3 ± 1.7 Ac | 0.0 Ad | |
| 90 | 18.33 ± 1.67 Bd | 93.7 ± 0.3 ABa | 61.7 ± 6.3 Bb | 36.7 ± 3.3 Bc | 8.3 ± 1.7 Ac | 0.0 Ae | |
| 120 | 26.7 ± 1.92 Ac | 92.7 ± 0.7 Aa | 83.3 ± 7.2 Aa | 46.7 ± 6.4 Ab | 10.0 ± 1.7 Ac | 0.0 Ad | |
| 15 | 0 | 0.0 Da | 0.0 Fa | 0.0 Fa | 0.0 Da | 0.0 Ca | 0.0 Aa |
| 15 | 0.0 Db | 0.0 Fb | 5.7 ± 3.2 EFa | 0.0 Db | 0.0 Cb | 0.0 Ab | |
| 30 | 0.0 Dc | 20.0 ± 13.6 DEa | 6.7 ± 2.7 DEFa | 1.7 ± 1.7 Dab | 0.0 Cc | 0.0 Ac | |
| 45 | 0.0 Db | 11.7 ± 4.2 Ea | 0.0 Fb | 0.0 Db | 3.3 ± 3.3 BCb | 0.0 Ab | |
| 60 | 0.0 Da | 0.0 Fa | 0.0 Fa | 0.0 Da | 0.0 Ca | 0.0 Aa | |
| 75 | 0.0 Db | 10.0 ± 0.0 Ea | 3.3 ± 3.3 EFb | 1.7 ± 1.7 Db | 1.7 ± 1.7 BCb | 0.0 Ab | |
| 90 | 0.0 Da | 6.7 ± 4.7 EFa | 0.0 Fa | 0.0 Da | 0.0 Ca | 0.0 Aa | |
| 120 | 0.0 Da | 1.7 ± 1.7 Fa | 0.0 Fa | 0.0 Da | 0.0 Ca | 0.0 Aa | |
| Source of Variation | Df | Mean Squares | f-Value | p-Value |
|---|---|---|---|---|
| Cold-moist stratification duration | 9 | 3266.52 | 82.75 | 0.00 |
| Germination temperature | 5 | 10,079.32 | 255.34 | 0.00 |
| Cold-moist stratification duration × germination temperature | 45 | 315.15 | 7.94 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, S.; Deng, Z.; Wang, X.; Lu, H.; Xue, H. Effects of Temperature, Scarification, Stratification, Phytohormones, and After-Ripening on the Dormancy and Germination of Eucommia ulmoides Oliv. Seeds. Forests 2021, 12, 1593. https://doi.org/10.3390/f12111593
Deng S, Deng Z, Wang X, Lu H, Xue H. Effects of Temperature, Scarification, Stratification, Phytohormones, and After-Ripening on the Dormancy and Germination of Eucommia ulmoides Oliv. Seeds. Forests. 2021; 12(11):1593. https://doi.org/10.3390/f12111593
Chicago/Turabian StyleDeng, Shiming, Zhijun Deng, Xiaofeng Wang, Hai Lu, and Hua Xue. 2021. "Effects of Temperature, Scarification, Stratification, Phytohormones, and After-Ripening on the Dormancy and Germination of Eucommia ulmoides Oliv. Seeds" Forests 12, no. 11: 1593. https://doi.org/10.3390/f12111593
APA StyleDeng, S., Deng, Z., Wang, X., Lu, H., & Xue, H. (2021). Effects of Temperature, Scarification, Stratification, Phytohormones, and After-Ripening on the Dormancy and Germination of Eucommia ulmoides Oliv. Seeds. Forests, 12(11), 1593. https://doi.org/10.3390/f12111593