
Projections for Mexico’s Tropical Rainforests Considering Ecological Niche and Climate Change

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Selection
2.2. Geographic Records and Bioclimatic Variables
2.3. Niche and Species Distribution Modeling
2.4. Mapping Potential Distribution and Percent Changes
2.5. Proposed Conservation Areas
3. Results
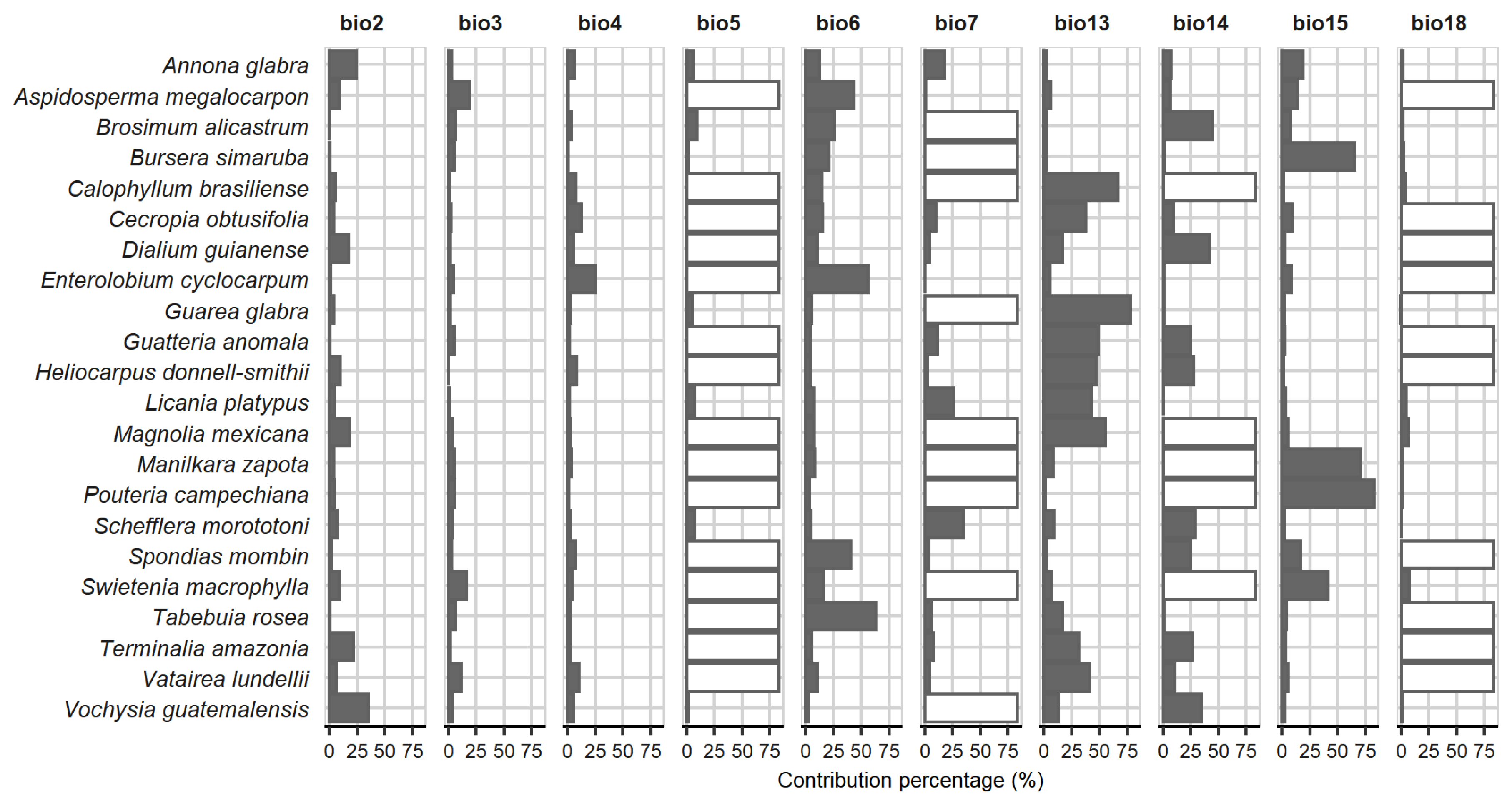
3.1. Geographic Records and Bioclimatic Variables
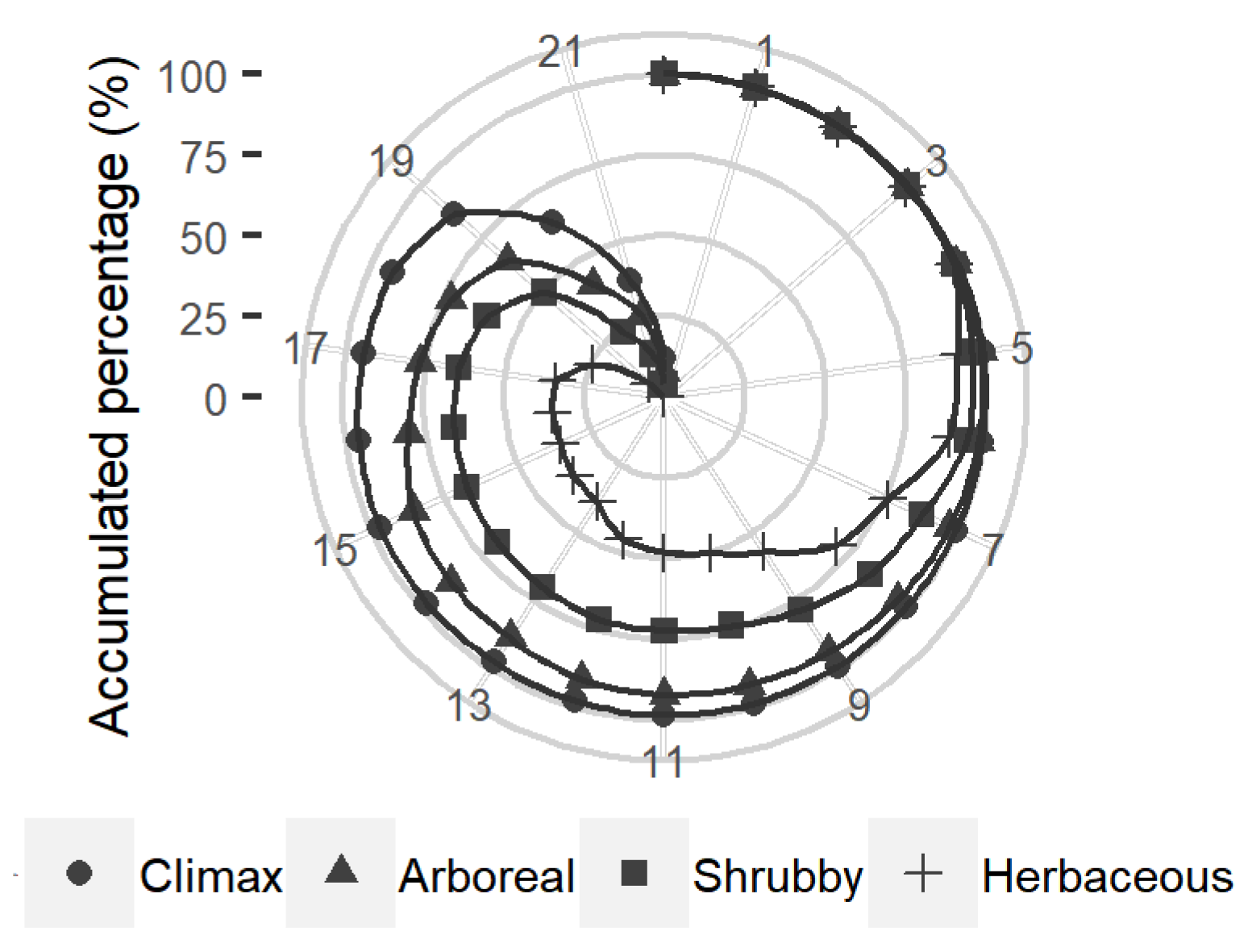
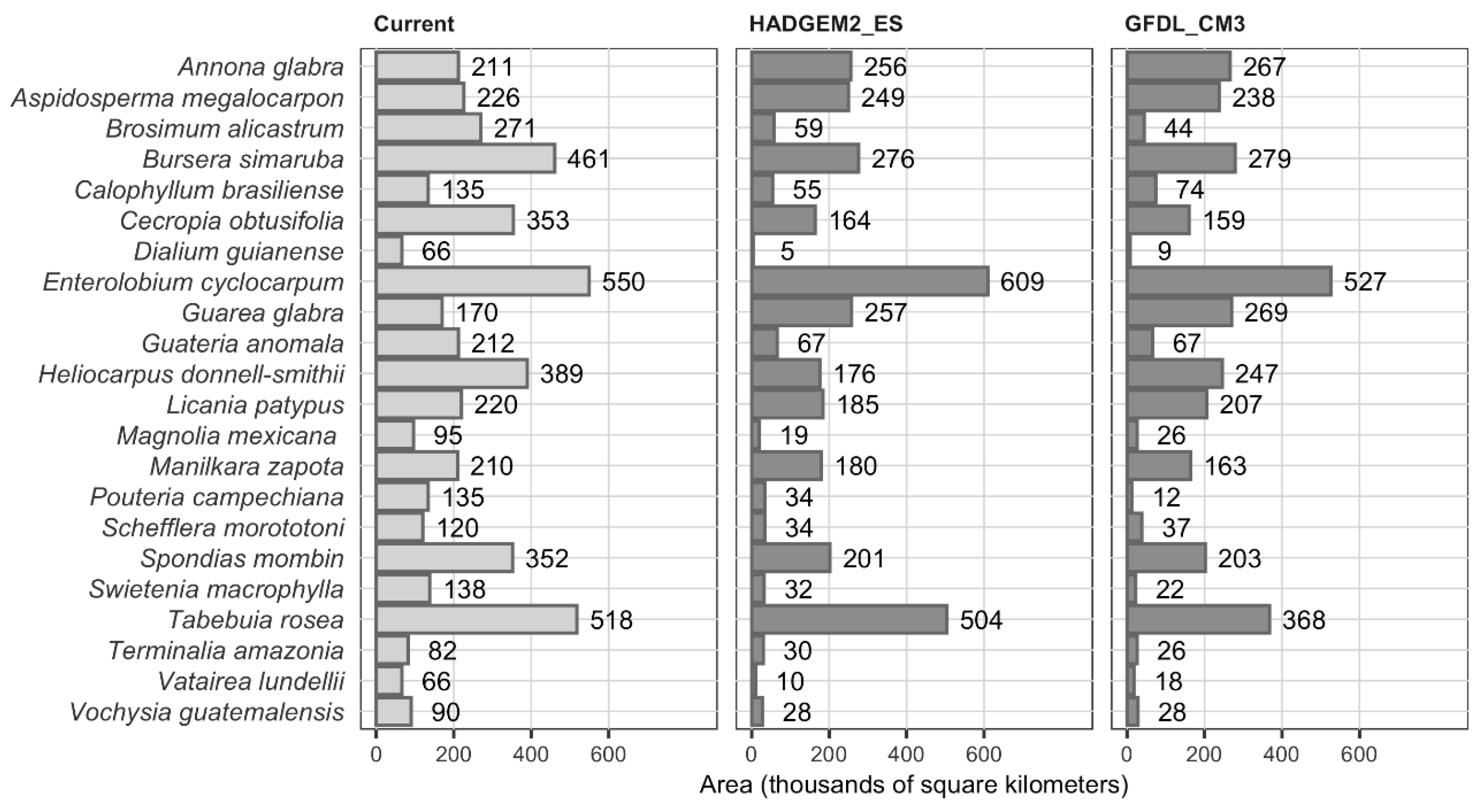
3.2. Niche Modeling and Species Distribution
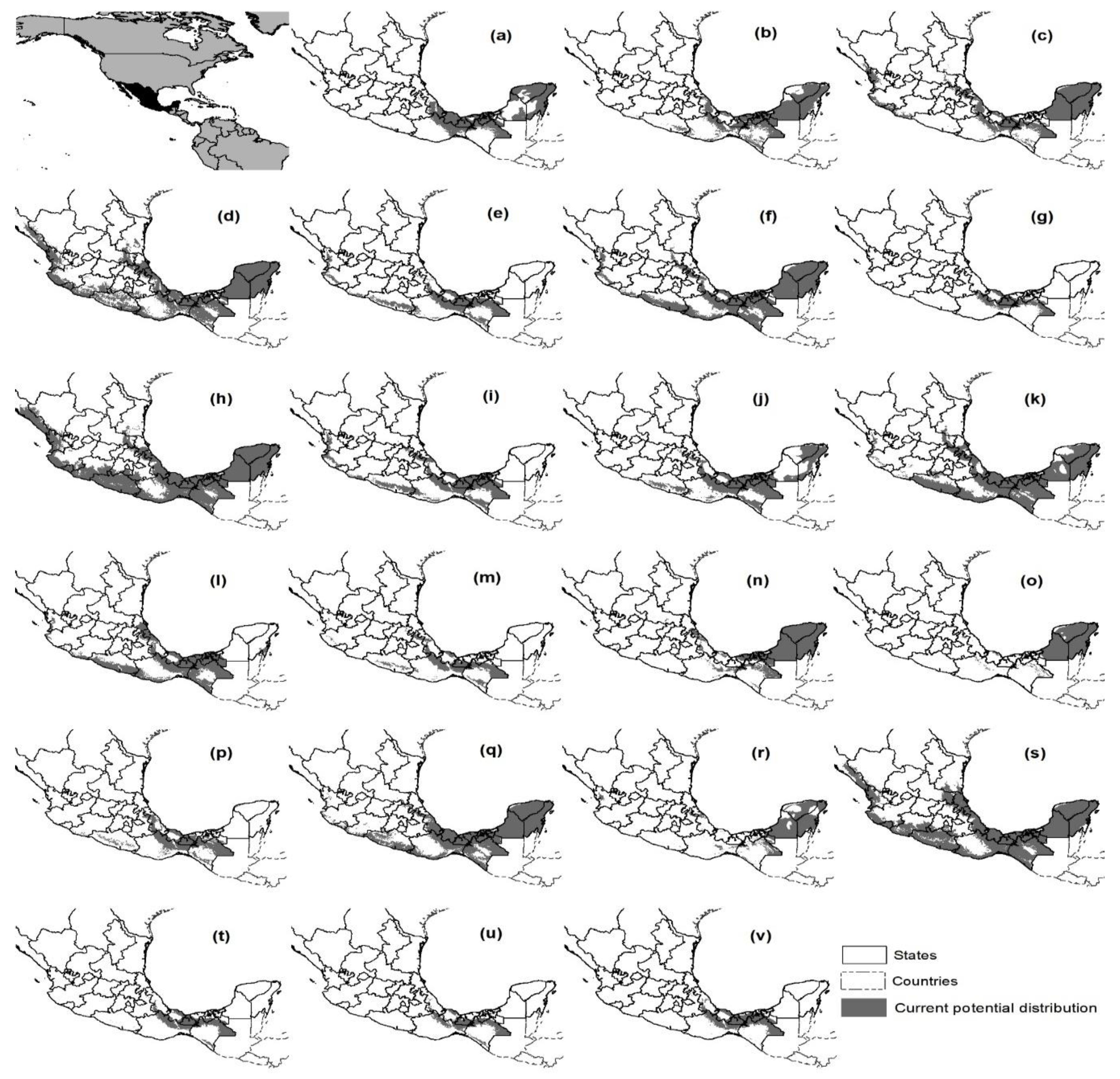
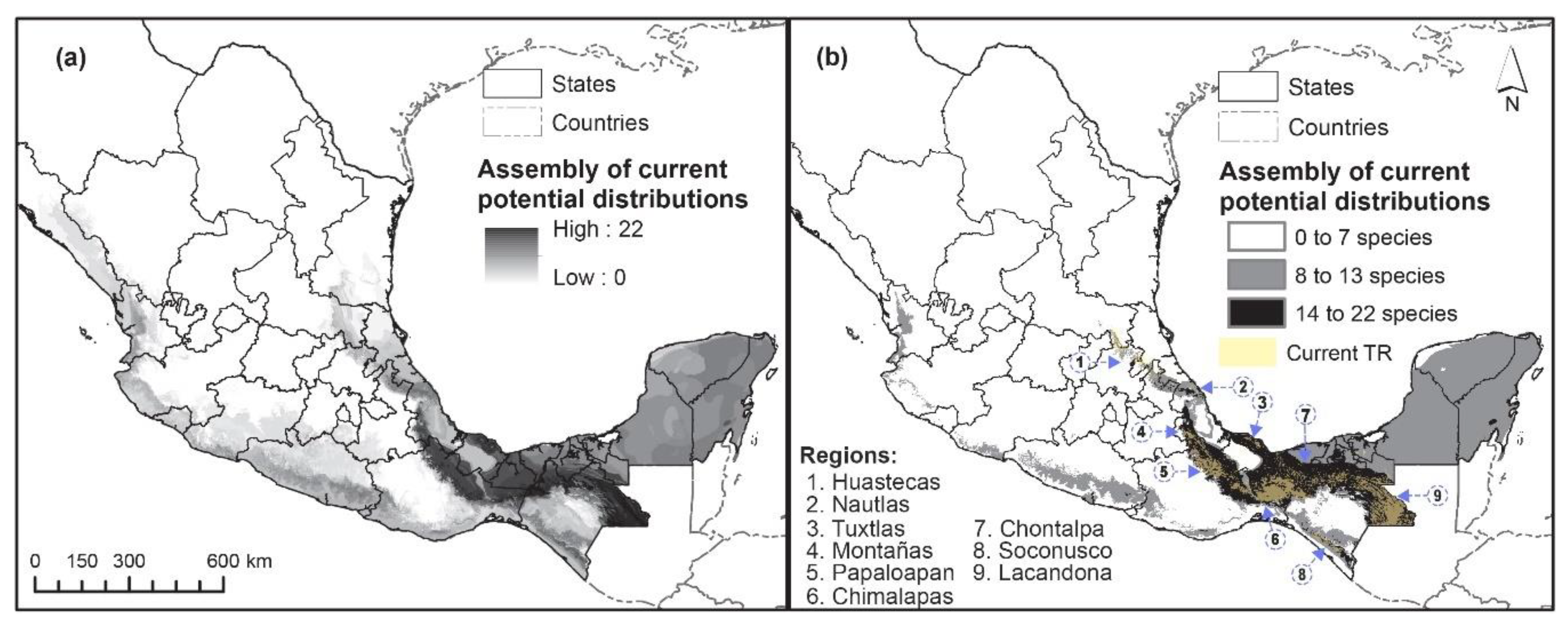
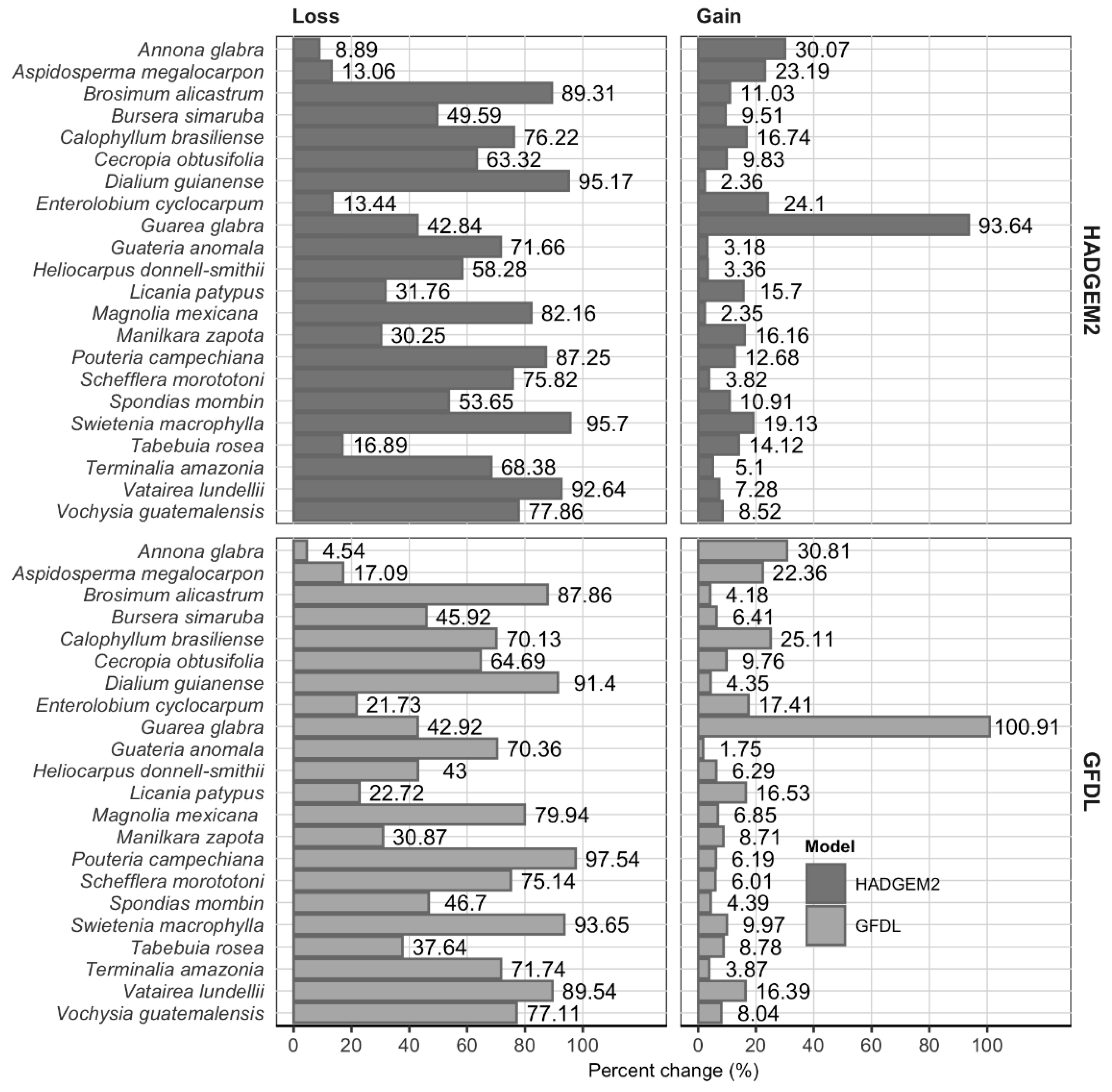
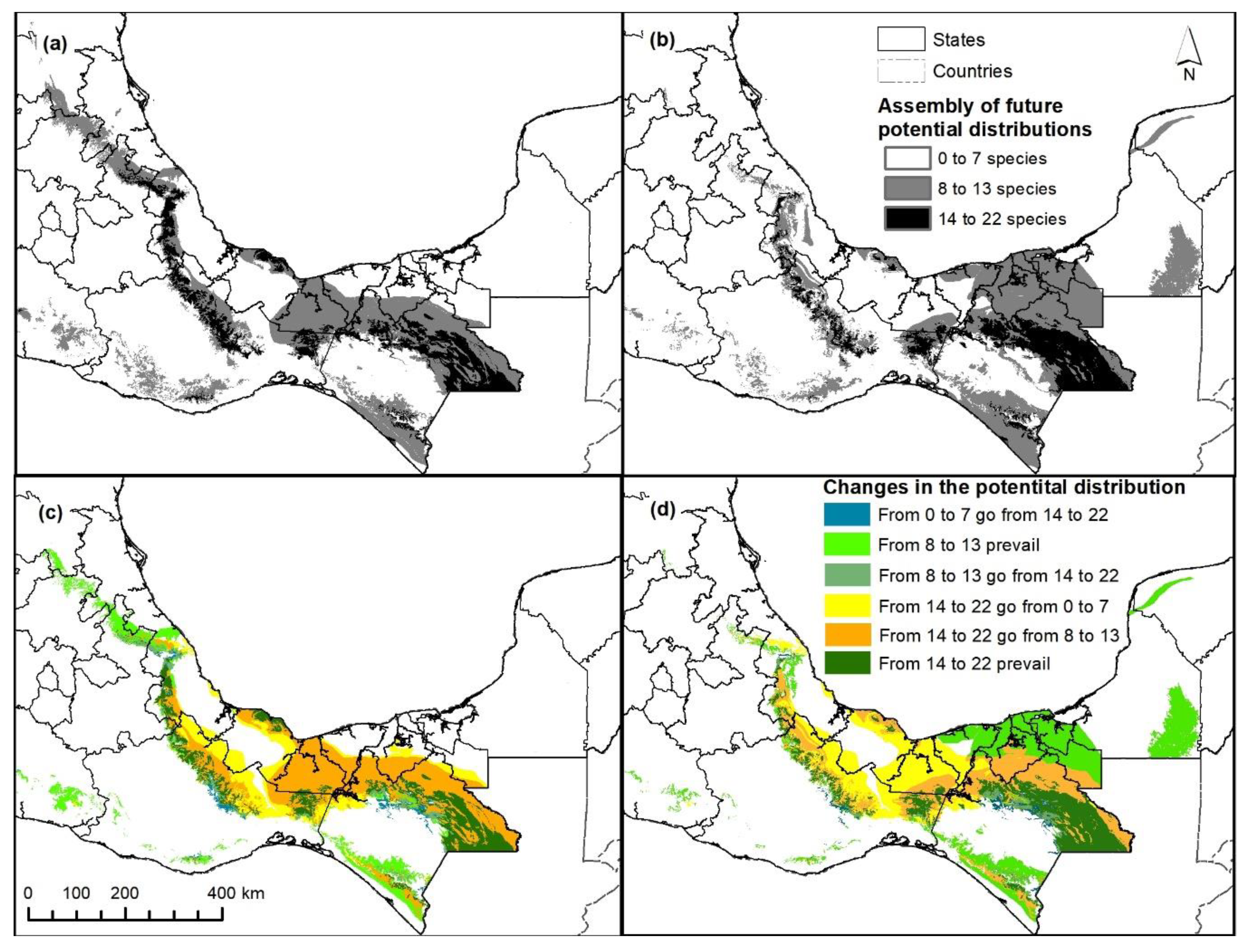
3.3. Mapping Potential Distribution and Changes
3.4. Proposed Conservation Areas
4. Discussion
4.1. Niche Modeling and Species Distribution
4.2. Projected Changes and Proposed Conservation Areas
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pennington, T.; Sarukán, J. Árboles Tropicales de México. Manual Para la Identificación de Las Principales Especies (Ediciones Cientficas Universitarias), Tercera ed.; UNAM, FCE, Eds.; Fondo de Cultura Económica: México DF, Mexico, 2005; ISBN 9681678559. [Google Scholar]
- Miranda, F.; Hernández-X., E. Los Tipos de Vegetación de México y Su Clasificación. Bol. Soc. Bot. México 1963, 28, 29–179. [Google Scholar] [CrossRef]
- Challenger, A. Utilización y Conservación de Los Ecosistemas Terrestres de México: Pasado, Presente y Futuro; Comisión Nacional para el Uso y Conocimiento de la Biodiversidad, Instituto de Biología de la UNAM y Agrupación Sierra Madre S.C: México DF, Mexico, 1988; ISBN 970-9000-02-0. [Google Scholar]
- INEGI. Conjunto de Datos Vectoriales de Uso Del Suelo y Vegetación Escala 1:250,000, Serie V (Capa Unión); Instituto Nacional de Estadística y Geografía (INEGI): Aguascalientes, Mexico, 2016.
- Moreno-Sanchez, R.; Moreno-Sanchez, F.; Torres-Rojo, J.M. National Assessment of the Evolution of Forest Fragmentation in Mexico. J. For. Res. 2011, 22, 167–174. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Rös, M.; Escobar, F.; Melo, F.P.L.; Santos, B.A.; Tabarelli, M.; Chazdon, R. Plant β-Diversity in Fragmented Rain Forests: Testing Floristic Homogenization and Differentiation Hypotheses. J. Ecol. 2013, 101, 1449–1458. [Google Scholar] [CrossRef] [Green Version]
- Carrara, E.; Arroyo-Rodríguez, V.; Vega-Rivera, J.H.; Schondube, J.E.; de Freitas, S.M.; Fahrig, L. Impact of Landscape Composition and Configuration on Forest Specialist and Generalist Bird Species in the Fragmented Lacandona Rainforest, Mexico. Biol. Conserv. 2015, 184, 117–126. [Google Scholar] [CrossRef]
- Zelazowski, P.; Malhi, Y.; Huntingford, C.; Sitch, S.; Fisher, J.B. Changes in the Potential Distribution of Humid Tropical Forests on a Warmer Planet. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 137–160. [Google Scholar] [CrossRef] [PubMed]
- López-Arce, L.; Ureta-Sánchez Cordero, C.; Granados-Sánchez, D.; Rodríguez-Esparza, L.; Monterroso-Rivas, A. Identifying Cloud Forest Conservation Areas in Mexico from the Potential Distribution of 19 Representative Species. Heliyon 2019, 5. [Google Scholar] [CrossRef]
- Christensen, J.H.; Kumar, K.K.; Aldria, E.; An, S.-I.; Cavalcanti, I.F.A.; De Castro, M.; Dong, W.; Goswami, P.; Hall, A.; Kanyanga, J.K.; et al. IPCC 2013 Chapter 14: Climate Phenomena and Their Relevance for Future Regional Climate Change Supplementary Material; Stocker, T.F., Qin, D., Plattner., G.K., Tignor, M., Allen, S.K., Boschung, A., Nauels, Y., Xia, V., Midgley, B., Eds.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- IPCC. AR5 Climate Change 2014: Impacts, Adaptation, and Vulnerability; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014; Volume 62. [Google Scholar]
- SEMARNAT-INECC. Sexta Comunicación Nacional y Segundo Informe Bienal de Actualización Ante La Convención Marco de Las Naciones Unidas Sobre El Cambio Climático; Secretaría de Medio Ambiente y Recursos Naturales: Ciudad de México, Mexico, 2018.
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; van Ypserle, J.P. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014; Volume 91, ISBN 9789291691432. [Google Scholar]
- Sierra, R.; Campos, F.; Chamberlin, J. Assessing Biodiversity Conservation Priorities: Ecosystem Risk and Representativeness in Continental Ecuador. Landsc. Urban Plan. 2002, 59, 95–110. [Google Scholar] [CrossRef]
- Peterson, A.; Soberón, J.; Pearson, R.; Anderson, R.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M. Ecological Niches and Geographic Distributions (MPB-49); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Aarts, G.; Fieberg, J.; Matthiopoulos, J. Comparative Interpretation of Count, Presence-Absence and Point Methods for Species Distribution Models. Methods Ecol. Evol. 2012, 3, 177–187. [Google Scholar] [CrossRef]
- Gomes, V.H.F.; Ijff, S.D.; Raes, N.; Amaral, I.L.; Salomão, R.P.; Coelho, L.D.S.; Matos, F.D.D.A.; Castilho, C.V.; Filho, D.D.A.L.; López, D.C.; et al. Species Distribution Modelling: Contrasting Presence-Only Models with Plot Abundance Data. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Soberón, J.; Osorio-Olvera, L.; Peterson, T. Diferencias Conceptuales Entre Modelación de Nichos y Modelación de Áreas de Distribución. Rev. Mex. Biodivers. 2017, 88, 437–441. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding Remark. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The First Species Distribution Modelling Package, Its Early Applications and Relevance to Most Current MaxEnt Studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of Consensus Methods in Predictive Species Distribution Modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Ferrier, S.; Dudík, M.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the Black Box: An Open-Source Release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Rzedowski, J. Vegetación de México; Primera, Ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, Mexico, 2006.
- Curtis, J.F.; Mclntosh, R. An Upload Forest Continuum in the Pariré-Forest Border Region of Wisconsin. Ecology 1951, 32, 476–696. [Google Scholar] [CrossRef]
- Corella, J.F.; Valdez, H.J.I.; Cetina, A.V.M.; González, C.F.V.; Trinidad, S.A.; Aguirre, R.J. Estructura Forestal de Un Bosque de Mangles Al Noroeste Del Estado de Tabasco, México. Cienc. For. México 2002, 26, 73–102. [Google Scholar]
- Maldonado-Sánchez, E.A.; Maldonado-Mares, F. Estructura y Diversidad Arborea de Una Selva Alta Perennifolia En Tlacotalpa, Tabasco, México. Univ. Cienc. Tróp. Humed. 2010, 26, 235–245. [Google Scholar]
- CONAFOR. UACh Inventario Nacional Forestal y de Suelos 2004–2009. 2009. Available online: http://www.conafor.gob.mx/biblioteca/Inventario-Nacional-Forestal-y-de-Suelos.pdf (accessed on 22 September 2019).
- CONAFOR. Metodología Del Inventario Nacional Forestal y de Suelos 2004—2009; Comisión Nacional Forestal: Guadalajara, Mexico, 2009.
- Vázquez-Negrín, I.; Castillo-Acosta, O.; Valdez-Hernández, J.I.; Zavala-Cruz, J.; Martínez-Sánchez, J.L. Estructura y Composición Florística de La Selva Alta Perennifolia En El Ejido Niños Héroes Tenosique, Tabasco, México. Polibotánica 2011, 32, 41–61. [Google Scholar]
- Martínez-Sánchez, J.L. Comparación de La Diversidad Estructural de Una Selva Alta Perennifolia y Una Mediana Subperennifolia En Tabasco, México. Madera Bosques 2016, 22, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Ibarra-Manríquez, G.; Cornejo-Tenorio, G. Diversidad de Frutos de Los Árboles Del Bosque Tropical Perennifolio de México. Acta Bot. Mex. 2010, 90, 51–104. [Google Scholar] [CrossRef] [Green Version]
- Arce-Romero, A.R.; Monterroso-Rivas, A.I.; Gómez-Díaz, J.D.; Cruz-León, A. Ciruelas Mexicanas (Spondias spp.): Su Aptitud Actual y Potencial Con Escenarios de Cambio Climático Para México. Rev. Chapingo Ser. Hortic. 2017, 23, 5–19. [Google Scholar] [CrossRef]
- SEMARNAT. NORMA Oficial Mexicana NOM-059-SEMARNAT-2010, Protección Ambiental-Especies Nativas de México de Flora y Fauna Silvestres-Categorías de Riesgo y Especificaciones Para Su Inclusión, Exclusión o Cambio-Lista de Especies En Riesgo; Secretaría de Medio Ambiente y Recursos Naturales: Ciudad de México, Mexico, 2010; p. 77.
- GBIF [Global Information Facility] Free and Open Access to Biodiversity Data. Available online: GBIF.org (accessed on 22 September 2019).
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. SpThin: An R Package for Spatial Thinning of Species Occurrence Records for Use in Ecological Niche Models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R Package for Conducting Spatially Independent Evaluations and Estimating Optimal Model Complexity for Maxent Ecological Niche Models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic Predictors for Supporting Ecological Applications in the Conterminous United States; U.S. Geological Survey (USGS): Fort Collins, CO, USA, 2012.
- Hijmans, R.J.; Cameron, S.E.; Parram, J.L.; Jones, P.G.; Jarvis, A. Very High Resolution Interpolated Climate Surfaces for Land Areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- WorldClim Global Climate Data. Available online: https://www.worldclim.org/ (accessed on 22 September 2019).
- Gómez-Díaz, J.; Etchevers-Barra, J.; Monterroso-Rivas, A.; Gay-García, C.; Campo-Alves, J.; Martínez-Menes, M. Spatial Estimation of Mean Temperature and Precipitation in Areas of Scarce Meteorological Information. Atmósfera 2008, 21, 35–56. [Google Scholar]
- Fernández Eguiarte, A.; Zavala Hidalgo, J.; Romero Centeno, R. Atlas Climático Digital de México. Available online: http://uniatmos.atmosfera.unam.mx/ACDM/ (accessed on 22 September 2019).
- Estrada, F.; Guerrero, V.M.; Gay-García, C.; Martínez-López, B. A Cautionary Note on Automated Statistical Downscaling Methods for Climate Change. Clim. Chang. 2013, 120, 263–276. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Interpretation of Models of Fundamental Ecological Niches and Species’ Distributional Areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- INEGI; CONABIO. INE Ecorregiones Terrestres de México 2008. Available online: http://www.conabio.gob.mx/informacion/gis/maps/geo/ecort08gw.zip (accessed on 22 September 2019).
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, T.A.; Barve, N.; Osorio-Olvera, L. Kuenm: An R Package for Detailed Development of Ecological Niche Models Using Maxent. PeerJ 2019, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanaugh, J.E. Unifying the Derivations for the Akaike and Corrected Akaike Information Criteria. Stat. Probab. Lett. 1997, 33, 201–208. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking Receiver Operating Characteristic Analysis Applications in Ecological Niche Modeling. Ecol. Modell. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Anderson, R.P.; Lew, D.; Peterson, A.T. Evaluating Predictive Models of Species’ Distributions: Criteria for Selecting Optimal Models. Ecol. Modell. 2003, 162, 211–232. [Google Scholar] [CrossRef]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Automat. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological Niche Modeling in Maxent: The Importance of Model Complexity and the Performance of Model Selection Criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Granados Sánchez, D.; Tapia Vargas, R. Comunidades Vegetales, Primera ed.; Colección Cuadernos Universitarios—Serie de Agronomía No. 19; Universidad Autónoma Chapingo: México DF, Mexico, 1990; ISBN 968-884-097-1. [Google Scholar]
- Granados Sánchez, D.; López Rios, G.F. Sucesión Ecológica, Dinámica Del Ecosistema, Primera ed.; Universidad Autónoma Chapingo: Texcoco, Mexico, 2000; ISBN 968-884-450-0. [Google Scholar]
- Whittaker, R.H. A Consideration of Climax Theory: The Climax as a Population and Pattern. Ecol. Monogr. 1953, 23, 41–78. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Levin, S.A. The Role of Mosaic Phenomena in Natural Communities. Theor. Popul. Biol. 1977, 12, 117–139. [Google Scholar] [CrossRef]
- Bezaury-Creel, J.E.; Torres, J.F.; Ochoa-Ochoa, L.M.; Castro-Campos, M.; Moreno, N. Base de Datos Geográfica de Áreas Naturales Protegidas Estatales y Del Distrito Federal de México (2009). 2010. Available online: http://www.conabio.gob.mx/informacion/gis/maps/geo/anpe09gw.zip (accessed on 22 September 2019).
- Comisión Nacional de Áreas Naturales Protegidas Áreas Naturales Protegidas Federales de La República. Available online: http://sig.conanp.gob.mx/website/pagsig/datos_anp.htm (accessed on 22 September 2019).
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The Effect of Sample Size and Species Characteristics on Performance of Different Species Distribution Modeling Methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Vignette of Package ‘Dismo’. Species Distribution Modeling Package. Circles 2020, 9, 1–68. [Google Scholar]
- Elith, J.; Kearney, M.; Phillips, S. The Art of Modelling Range-Shifting Species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Flores-Tolentino, M.; Ortiz, E.; Villaseñor, J.L. Ecological Niche Models as a Tool for Estimating the Distribution of Plant Communities. Rev. Mex. Biodivers. 2019, 90. [Google Scholar] [CrossRef]
- Maldonado-Sánchez, E.A.; Ochoa-Gaona, S.; Ramos-Reyes, R.; Guadarrama-Olivera, M.D.; González-Valdivia, N.; de Jong, B.H. La Selva Inundable de Canacoite En Tabasco, México, Una Comunidad Vegetal Amenazada. Acta Bot. Mex. 2016, 115, 75–110. [Google Scholar] [CrossRef] [Green Version]
- Roberge, J.-M.; Angelstam, P. Usefulness of the Umbrella Species Concept as a Conservation Tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Caro, T.M.; O’Doherty, G. On the Use of Surrogate Species in Conservation Biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Challenger, A. Utilzación y Conservación de Los Ecosistemas Terrestres de México; UNAM, Instituto de Biología: Ciudad de México, Mexico, 1998; ISBN 970-90000-02-0. [Google Scholar]
- Villavicencio-Enríquez, L.; Valdez-Hernández, J.I. Análisis de La Estructura Arbórea Del Sistema Agroforestal Rusticano de Café En San Miguel, Veracruz, México. Agrociencia 2003, 37, 413–422. [Google Scholar]
- Odum, E.P. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1953. [Google Scholar]
- Trejo, I.; Martínez-Meyer, E.; Calixto-Pérez, E.; Sánchez-Colón, S.; Vázquez De La Torre, R.; Villers-Ruiz, L. Analysis of the Effects of Climate Change on Plant Communities and Mammals in México. Atmosfera 2011, 24, 1–14. [Google Scholar]
- Estrada-Contreras, I.; Equihua, M.; Castillo-Campos, G.; Rojas-Soto, O. Climate Change and Effects on Vegetation in Veracruz, Mexico: An Approach Using Ecological Niche Modelling. Acta Bot. Mex. 2015, 2015, 73–93. [Google Scholar] [CrossRef] [Green Version]
- Booth, T.H.; Muir, P.R. Climate Change Impacts on Australia’s Eucalypt and Coral Species: Comparing and Sharing Knowledge across Disciplines. Wiley Interdiscip. Rev. Clim. Chang. 2020, 11. [Google Scholar] [CrossRef]
- García, E. Modificaciones Al Sistema de Clasificación Climática de Köppen; Serie Libros, núm. 6; Instituto de Geografía, UNAM: Ciudad de México, Mexico, 2004. [Google Scholar]
- Romahn-Hernández, L.F.; Rodríguez-Trejo, D.A.; Villanueva-Morales, A.; Monterroso-Rivas, A.I.; Pérez-Hernández, M.D.J. Rango Altitudinal: Factor de Vigor Forestal y Determinante En La Regeneración Natural Del Oyamel. Entrecienc. Diálogos Soc. Conoc. 2020, 8. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Reference | Notes |
|---|---|---|
| Annona glabra L. * | [33] | |
| Aspidosperma megalocarpon Müll. Arg. | [1,2] | |
| Brosimum alicastrum Sw. * | [1,2,27,34] | |
| Bursera simaruba (L.) Sarg. * | [1,30] | |
| Calophyllum brasiliense Cambess. * | [2,27] | NOM(T) |
| Cecropia obtusifolia Bertol.* | [2,30] | SSS |
| Dialium guianense (Aubl.) Sandwith * | [2,30] | |
| Enterolobium cyclocarpum (Jacq.) Griseb. * | [1] | SSS |
| Guarea glabra Vahl * | [27,30] | |
| Guatteria anomala R.E. Fr. | [1,27] | NOM(T) |
| Heliocarpus donnell-smithii Rose ex Donn.Sm. * | [2] | SSS |
| Licania platypus (Hemsl.) Fritsch | [1,2,30] | |
| Magnolia mexicana DC. | [1,2] | NOM(T) |
| Manilkara zapota (L.) P. Royen * | [1,27] | CV, NOM(SP) |
| Pouteria campechiana (Kunth) Baehni | [1,27] | CV |
| Schefflera morototoni (Aubl.) Maguire, Steyerm. & Frodin * | [35] | |
| Spondias mombin Jacq. * | [30,34,36] | |
| Swietenia macrophylla King | [2,27] | CV |
| Tabebuia rosea (Bertol.) Bertero ex A.DC. * | [27] | SSS |
| Terminalia amazonia (J.F.Gmel.) Excell * | [2,27] | |
| Vatairea lundellii (Standl.) Killip ex Record * | [2,27,30] | NOM(DE) |
| Vochysia guatemalensis Donn. Sm. * | [2,27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Hernández, A.F.; Monterroso-Rivas, A.I.; Granados-Sánchez, D.; Villanueva-Morales, A.; Santacruz-Carrillo, M. Projections for Mexico’s Tropical Rainforests Considering Ecological Niche and Climate Change. Forests 2021, 12, 119. https://doi.org/10.3390/f12020119
Santos-Hernández AF, Monterroso-Rivas AI, Granados-Sánchez D, Villanueva-Morales A, Santacruz-Carrillo M. Projections for Mexico’s Tropical Rainforests Considering Ecological Niche and Climate Change. Forests. 2021; 12(2):119. https://doi.org/10.3390/f12020119
Chicago/Turabian StyleSantos-Hernández, Antonio Fidel, Alejandro Ismael Monterroso-Rivas, Diódoro Granados-Sánchez, Antonio Villanueva-Morales, and Malinali Santacruz-Carrillo. 2021. "Projections for Mexico’s Tropical Rainforests Considering Ecological Niche and Climate Change" Forests 12, no. 2: 119. https://doi.org/10.3390/f12020119
APA StyleSantos-Hernández, A. F., Monterroso-Rivas, A. I., Granados-Sánchez, D., Villanueva-Morales, A., & Santacruz-Carrillo, M. (2021). Projections for Mexico’s Tropical Rainforests Considering Ecological Niche and Climate Change. Forests, 12(2), 119. https://doi.org/10.3390/f12020119