
Empowering Forest Owners with Simple Volume Equations for Poplar Plantations in the Órbigo River Basin (NW Spain)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Trees and Soil Measurements, and Data Analysis
3. Results
4. Discussion
4.1. Simplified Volume and Height Equations
4.2. Limitations of the Simplification Approach
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rueda Fernández, J.; García Caballero, J.L. Parcela de Experimentación de Clones de Chopos LE-3 Gradefes; Junta de Castilla y León: Valladolid, Spain, 2013. [Google Scholar]
- Freer-Smith, P.; Muys, B.; Bozzano, M.; Drössler, L.; Farrelly, N.; Jactel, H.; Korhonen, J.; Minotta, G.; Nijnik, M.; Orazio, C. Plantation forests in Europe: Challenges and opportunities. Sci. Policy 2019, 9. [Google Scholar] [CrossRef]
- Christersson, L. Silvicultura de rotación corta: Un complemento de la silvicultura “convencional”. Unasylva 2006, 57, 223. [Google Scholar]
- Vadell, E.; De-Miguel, S.; Pemán, J. Large-scale reforestation and afforestation policy in Spain: A historical review of its underlying ecological, socioeconomic and political dynamics. Land Use Policy 2016, 55, 37–48. [Google Scholar] [CrossRef]
- Martín-García, J.; Barbaro, L.; Diez, J.J.; Jactel, H. Contribution of poplar plantations to bird conservation in riparian landscapes. Silva Fennica. 2013, 47, 4. [Google Scholar] [CrossRef] [Green Version]
- Díaz, L.; Romero, C. Caracterización económica de las choperas en Castilla y León: Rentabilidad y turnos óptimos; Libro de actas del I Simposio del Chopo: Zamora, Spain, 2001; pp. 489–500. [Google Scholar]
- Fernández Manso, A.; Hernanz Arroyo, G. El Chopo (Populus sp.): Manual de Gestión Forestal Sostenible; Junta de Castilla y León: Valladolid, Spain, 2004. [Google Scholar]
- Ambrosio Torrijos, Y.; Picos Martín, J.; Valero Gutiérrez del Olmo, E. Small non industrial forest owners cooperation examples in Galicia (NW Spain). In Proceedings of the FAO Workshop on Forest Operations Improvements in Farm Forest, Logarska Dolina, Slovenia, 9–14 September 2003. [Google Scholar]
- Dominguez, G.; Shannon, M. A wish, a fear and a complaint: Understanding the (dis)engagement of forest owners in forest management. Eur. J. For. Res. 2011, 130, 435–450. [Google Scholar] [CrossRef]
- Wiersum, K.F.; Elands, B.H.M.; Hoogstra, M.A. Small-scale forest ownership across Europe: Characteristics and future potential. Small Scale For. Econ. Manag. Policy 2005, 4, 1–9. [Google Scholar] [CrossRef]
- Rodriguez-Vicente, V.; Marey-Pérez, M.F. Characterization of nonindustrial private forest owners and teir influence on forest management aims and practices in Northern Spain. Small Scale For. 2009, 8, 479–513. [Google Scholar] [CrossRef]
- Vainio, A.; Paloniemi, R. Forest owners and power: A Foucauldian study on Finnish forest policy. Forest Policy Econ. 2012, 21, 118–125. [Google Scholar] [CrossRef]
- Emil Fraga, E.; Fidalgo, L.; Álvarez Rodríguez, E.; Rodriguez Soalleiro, R.; Oliveira, N.; Sixto, H. Crecimiento a medio turno de plantaciones madereras del clon RASPAJE en suelo ácido en Galicia. In Proceedings of the II Simposio del Chopo; Sociedad Pública de Infraestructuras y Medio Ambiente de Castilla y León: Valladolid, Spain, 2018; pp. 63–72. [Google Scholar]
- Bravo, F.; Grau, J.M.; González Antoñanzas, F. Análisis de modelos de producción para Populus x euroamericana en la cuenca del Duero. For. Syst. 1996, 5, 77–95. [Google Scholar]
- Rodríguez, F.; Molina, C. Análisis de modelos de perfil del fuste y estudio de la cilindricidad para tres clones de chopo (Populus x euramericana) en Navarra. For. Syst. 2003, 12, 73–85. [Google Scholar]
- Barrio-Anta, M.; Sixto Blanco, H.; Cañellas Rey de Viñas, I.; González Antoñanzas, F. Sistema de cubicación con clasificación de productos para plantaciones de Populus x euramericana (Dode) Guinier cv. ‘I-214’ en la meseta norte y centro de España. For. Syst. 2007, 16, 65–75. [Google Scholar]
- Rodríguez, F.; Pemán, J.; Aunços, A. A reduced growth model based on stand basal area. A case for hybrid poplar plantations in northeast Spain. For. Ecol. Manag. 2010, 259, 2093–2102. [Google Scholar] [CrossRef]
- Hjelm, B. Stem taper equations for poplars growing on farmland in Sweden. J. For. Res. 2013, 24, 15–22. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística–INE. Encuesta Sobre la Estructura de las Explotaciones Agrícolas. Available online: https://www.ine.es/dyngs/INEbase/es/operacion.htm?c=Estadistica_C&cid=1254736176854&menu=ultiDatos&idp=1254735727106 (accessed on 3 January 2021).
- Carreras, A.; Tafunell, X. Estadísticas Históricas de España: Siglos XIX-XX, 2nd ed.; Fundación BBVA: Bilbao, Spain, 2005. [Google Scholar]
- Marziliano, P.A.; Russo, D.; Altieri, V.; Macrì, G.; Lombardi, F. Optimizing the sample size to estimate growth in I-214 poplar plantations at definitive tree density for bioenergetic production. Agron. Res. 2018, 16, 821–837. [Google Scholar]
- Köppen, W.; Geiger, R. Das Geographische System der Klimate; Borntraeger: Berlin, Germany, 1936. [Google Scholar]
- Agencia Española de Meteorología–AEMET. 2020. Regionalización AR4-IPCC. Gráficos de evolución. Regionalización estadística análogos. Castilla y León. Accedido el 3 de febrero de 2020. Available online: http://www.aemet.es/es/serviciosclimaticos/cambio_climat/result_graficos?w=0&opc1=cle&opc2=P&opc3=Anual&opc4=0&opc6=1 (accessed on 10 December 2020).
- Gaussen, H.; Bagnouls, F. Saison Seche et Indice Xerotermique; Université de Toulouse, Faculté des Sciences: Toulouse, Francia, 1953. [Google Scholar]
- World Atlas of Desertification; Cherlet, M.; Hutchinson, C.; Reynolds, J.; Hill, J.; Sommer, S.; von Maltitz, G. (Eds.) Publication Office of the European Union: Luxembourg, 2018. [Google Scholar]
- Instituto Tecnológico Agrario de Castilla y León–ITACYL. Visor de Datos de Suelo. Available online: suelos.itacyl.es/visor_datos (accessed on 3 January 2021).
- Blanco, R. Estimación de Tasas de Crecimiento y Producción de Plantaciones de Chopo (Populus x Euroamericana) en la Ribera Media del Duero. Master’s Thesis, Public University of Navarre, Pamplona, Spain, 2020. [Google Scholar]
- Rueda, J.; García Caballero, J.R. Parcelas de Experimentación de Clones de Chopos LE-4 La Milla del Río; Consejería de Fomento y Medio Ambiente, Junta de Castilla y León: Valladolid, Spain, 2018. [Google Scholar]
- Rueda, J.; García Caballero, J.L.; López Negredo, L.; Gómez Cáceres, C. Parcela de Experimentación de Clones de Chopos LE-1 Valencia de Don Juan; Consejería de Fomento y Medio Ambiente, Junta de Castilla y León: Valladolid, Spain, 2006. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Theil, H. Applied Econometric Forecasting; North-Holland: Amsterdam, The Netherdlands, 1966. [Google Scholar]
- Vanclay, J.; Skovsgaard, J.P. Evaluating forest growth models. Ecol. Model. 1997, 98, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Freese, F. Testing accuracy. For. Sci. 1960, 6, 139–145. [Google Scholar]
- Reynolds, M.R. Estimating the error in model predictions. For. Sci. 1984, 30, 454–469. [Google Scholar]
- Blanco, J.A.; González, E. Exploring the sustainability of current management prescriptions for Pinus caribaea plantations in Cuba: A modelling approach. J. Trop. For. Sci. 2010, 22, 139–154. [Google Scholar]
- Blanco, J.A.; Seely, B.; Welham, C.; Kimmins, J.P.; Seebacher, T.M. Testing the performance of FORECAST, a forest ecosystem model, against 29 years of field data in a Pseudotsuga menziesii plantation. Can. J. For. Res. 2007, 37, 1808–1820. [Google Scholar] [CrossRef]
- Welham, C.; Van Rees, K.C.J.; Seely, B.; Kimmins, J.P. Projected long-term productivity in Saskatchewan hybrid poplar plantations: Weed competition and fertilizer effects. Can. J. For. Res. 2007, 37, 356–370. [Google Scholar] [CrossRef]
- Benomar, L.; DesRochers, A.; Larocque, G.R. The effects of spacing on growth, morphology and biomass production and allocation in two hybrid poplar clones growing in the boreal region of Canada. Trees 2012, 26, 939–949. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics: Update Edition; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Rodríguez Pleguezuelo, C.R.; Durán Zuazo, V.H.; Bielders, C.; Jiménez Bocanegra, J.A.; PereaTorres, F.; Francia Martínez, J.R. Bioenergy farming using woody crops: A review. Agron. Sustain. Dev. 2015, 35, 95–119. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Rauscher, H.M.; Isebrands, J.G.; Host, G.E.; Dickson, R.E.; Dickmann, D.I.; Crow, T.R.; Michael, D.A. ECOPHYS: An ecophysiological growth process model for juvenile poplar. Tree Physiol. 1990, 7, 255–281. [Google Scholar] [CrossRef]
- Asadi, F.; Alimohamadi, A. Assessing the performance of Populus caspica and Populus alba cuttings under different irrigation intervals. Agric. For. 2019, 65, 39–51. [Google Scholar] [CrossRef]
- Bagheri, R.; Ghasemi, R.; Calagari, M.; Merrikh, F. Effect of Different Irrigation Interval on Superior Poplar Clones Yield; Research Institute of Forest and Rangeland—RIFR; FAO: Tehran, Iran, 2005. [Google Scholar]
- Lorenc-Plucinska, G.; Walentynowicz, M.; Lwandowski, A. Poplar growth and wood production on a grassland irrigated for decades with potato starch wastewater. Agrofor. Syst. 2017, 91, 307–324. [Google Scholar] [CrossRef] [Green Version]
- Petty, J.A.; Swain, C. Factors influencing stem breakage in high winds. Forestry 1985, 58, 75–84. [Google Scholar] [CrossRef]
- Orzel, S. A comparative analysis of slenderness of the main tree species of the Niepolomice forest. Electron. J. Pol. Agric. Univ. 2007, 10, 13. [Google Scholar]
- Wang, Y.; Titus, S.J.; LeMay, V. Relationships between tree slenderness coefficients and tree or stand characteristics for major species in boreal mixedwood forests. Can. J. For. Res. 1998, 28, 1171–1183. [Google Scholar] [CrossRef]
- Navratil, S. Silvicultural systems for managing deciduous and mixedwood stands with white spruce understory. In Silvicultural of Temperate and Boreal Broadleaf-Conifer Mixture; Comeau, P.G., Thomas, K.D., Eds.; B.C. Ministry of Forests: Victoria, BC, Canada, 1996; pp. 35–46. [Google Scholar]
- Rueda, J.; García Caballero, J.L.; Cuevas, Y.; García-Jiménez, C.; Villar, C. Cultivo de chopos en Castilla y León; Consejería de Fomento y Medio Ambiente, Junta de Castilla y León: Valladolid, Spain, 2019. [Google Scholar]
- DeBell, D.S.; Harrington, C.A. Productivity of Populus in monoclonal and polyclonal blocks at three spacings. Can. J. For. Res. 1997, 27, 978–985. [Google Scholar] [CrossRef]
- Nguyen, T.H. Effects of Thinning on Growth and Development of Second Poplar Generations. Master’s Thesis, Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences, Alnarp, Sweden, 2018. no. 303. [Google Scholar]
- Feng, J.; Huang, P.; Wan, X. Interactive effects of wind and light on growth and architecture of poplar saplings. Ecol. Res. 2019, 34, 94–105. [Google Scholar] [CrossRef]
- United States Department of Agriculture. USDA. Hybrid Poplar: An Alternative Crop for the Intermountain West; USDA Technical note on Plant Materials no. 37; USDA: Aberdeen, WA, USA, 2001. [Google Scholar]
- Truax, B.; Gagnon, D.; Fortier, J.; Lambert, F. Biomass and volume yield in mature hybrid poplar plantations on temperate abandoned farmland. Forests 2014, 5, 3107–3130. [Google Scholar] [CrossRef] [Green Version]
- Montoya Oliver, J.M. Chopos y Choperas; Mundi Prensa: Madrid, Spain, 1988. [Google Scholar]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Wang, F.; Mladenoff, D.J.; Forrester, J.A.; Blanco, J.A.; Schelle, R.M.; Peckham, S.D.; Keough, C.; Lucash, M.S.; Gower, S.T. Multimodel simulations of forest harvesting effects on long-term productivity and CN cycling in aspen forests. Ecol. Appl. 2014, 24, 1374–1389. [Google Scholar] [CrossRef] [Green Version]
- Lo, Y.H.; Blanco, J.A.; González de Andrés, E.; Imbert, J.B.; Castillo, F.J. CO2 fertilization plays a minor role in long-term carbon accumulation patterns in temperate pine forests in the Pyrenees. Ecol. Model. 2019, 407, 108737. [Google Scholar] [CrossRef]
- Wu, S.; Harris, T.J.; McAuley, K.B. The use of simplified or misspecified models: The linear case. Can. J. Chem. Eng. 2007, 85, 386–398. [Google Scholar] [CrossRef]
- Kimmins, J.P.; Blanco, J.A.; Seely, B.; Welham, C.; Scoullar, K. Complexity in Modeling Forest Ecosystems; How Much is Enough? For. Ecol. Manag. 2008, 256, 1646–1658. [Google Scholar] [CrossRef]
- Blanco, J.A. Modelos ecológicos: Descripción, explicación y predicción. Ecosistemas 2013, 22, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, R.; Anand, M. Trees do not always act their age: Size-deterministic tree ring standardization for long-term trend estimation of shade-tolerant trees. Biogeosciences 2019, 16, 4815–4827. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.A.; Lo, Y.H.; Welham, C.; Larson, B. Productivity of forest ecosystems. In Sustainable Forest Management: From Principles to Practice; Innes, J.I., Tikina, A., Eds.; Earthscan: London, UK, 2017; pp. 72–100. [Google Scholar]
- Barrio-Anta, M.; Sixto-Blanco, H.; Cañellas-Rey de Viñas, I.; Castedo-Dorado, F. Dynamic growth model for I-214 poplar plantations in the northern and central plateaux in Spain. For. Ecol. Manag. 2008, 255, 1167–1178. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | Plot 1 | Plot 2 | Plot 3 | All Plots |
|---|---|---|---|---|
| Number of trees | 51 | 79 | 274 | 404 |
| DBH (cm) | 24.64 ± 7.05a | 25.70 ± 5.49a | 24.55 ± 3.48a | 24.78 ± 4.52 |
| Height (m) | 25.20 ± 5.48a | 22.68 ± 3.30b | 20.72 ± 2.13c | 21.67 ± 3.36 |
| Slenderness index (m/m) | 105.2 ± 16.6a | 89.9 ± 10.8b | 85.4 ± 10.3c | 88.8 ± 13.1 |
| Tree volume (m3) | 0.492 ± 0.386a | 0.457 ± 0.271a | 0.356 ± 0.149b | 0.393 ± 0.226 |
| Tree basal area (m2) | 0.051 ± 0.029a | 0.054 ± 0.023a | 0.048 ± 0.014a | 0.050 ± 0.018 |
| Variable | Plot 1 | Plot 2 | Plot 3 | All Plots |
|---|---|---|---|---|
| Annual DBH increment (cm year−1) | 2.33 ± 0.86 (9) | 2.16 ± 0.71 (7) | 2.90 ± 0.83 (4) | 2.39 ± 0.60 (8) |
| Annual height increment (m year−1) | 2.22 ± 0.93 (9) | 2.10 ± 0.84 (9) | 1.80 ± 0.50 (7) | 1.82 ± 0.60 (9) |
| Annual volume increment (m3 year−1) | 0.077 ± 0.064 (8) | 0.067 ± 0.034 (9) | 0.092 ± 0.050 (13) | 0.081 ± 0.040 (13) |
| Variable | Plot 1 | Plot 2 | Plot 3 |
|---|---|---|---|
| Soil pH | 7.346 ± 0.286ab | 7.210 ± 0.164b | 7.515 ± 0.212a |
| Conductivity (µs cm−1) | 66.400 ± 10.511b | 97.900 ± 31.3804a | 94.775 ± 14.076a |
| Organic Matter (%) | 6.880 ± 0.850a | 7.180 ± 0.537a | 3.130 ± 0.450b |
| Total C (%) | 4.270 ± 0.425a | 4.530 ± 0.453a | 1.970 ± 0.242b |
| Organic C (%) | 3.990 ± 0.492a | 4.160 ± 0.311a | 1.820 ± 0.277b |
| CaCO3 (%) | 2.290 ± 2.549a | 3.050 ± 3.253a | 1.270 ± 1.663a |
| N (%) | 0.400 ± 0.045a | 0.430 ± 0.057a | 0.170 ± 0.035b |
| Ca (%) | 0.514 ± 0.095a | 0.415 ± 0.126ab | 0.287 ± 0.109b |
| K (%) | 0.640 ± 0.066a | 0.561 ± 0.114ab | 0.447 ± 0.088b |
| Mg (%) | 0.214 ± 0.030a | 0.181 ± 0.041a | 0.131 ± 0.027b |
| Na (%) | 0.038 ± 0.006a | 0.036 ± 0.007a | 0.028 ± 0.005b |
| P (%) | 0.056 ± 0.009a | 0.046 ± 0.012a | 0.027 ± 0.007b |
| S (%) | 0.052 ± 0.011a | 0.044 ± 0.011a | 0.025 ± 0.005b |
| C/N | 10.656 ± 0.242a | 10.561 ± 0.486a | 11.367 ± 0.694b |
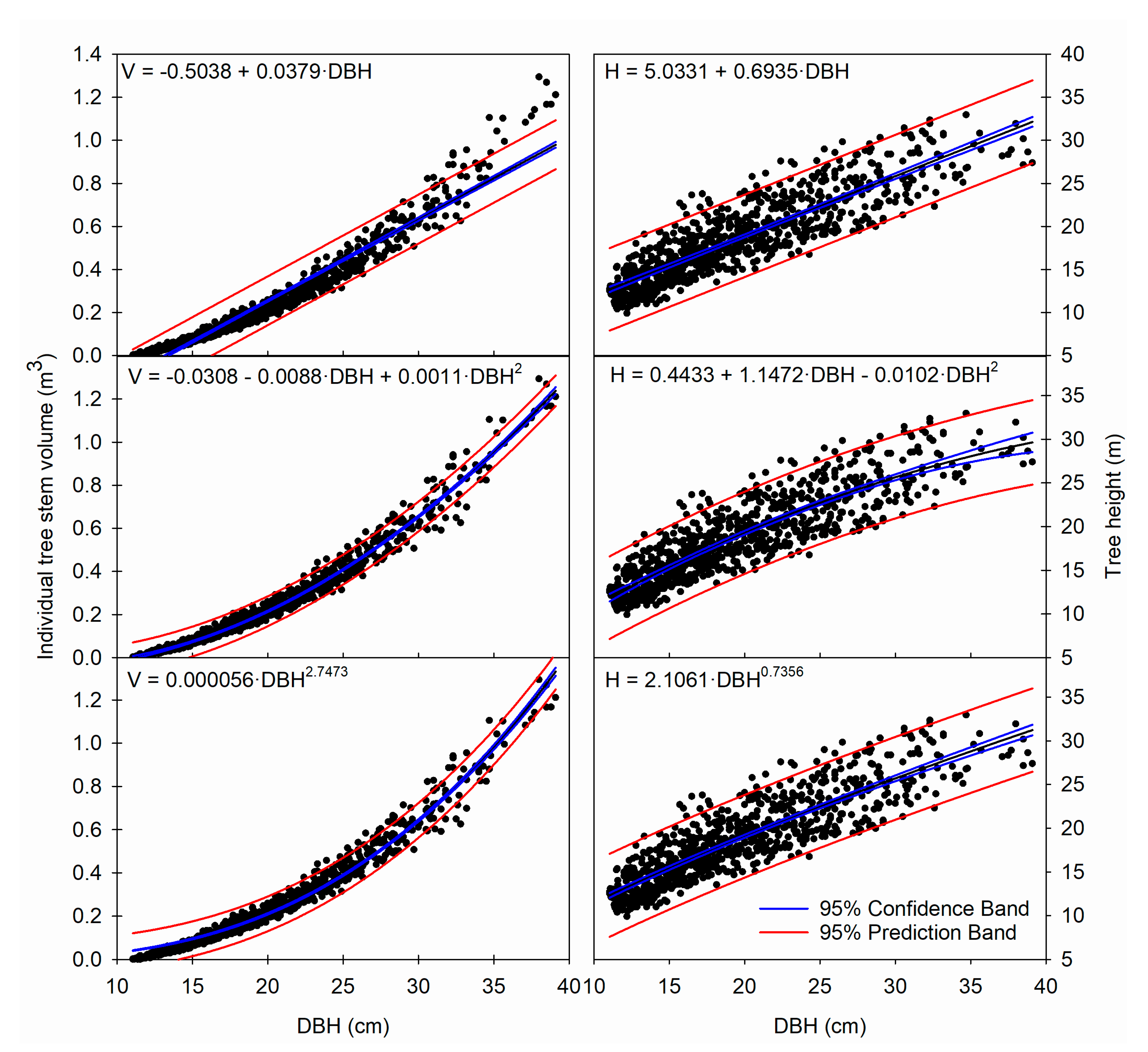
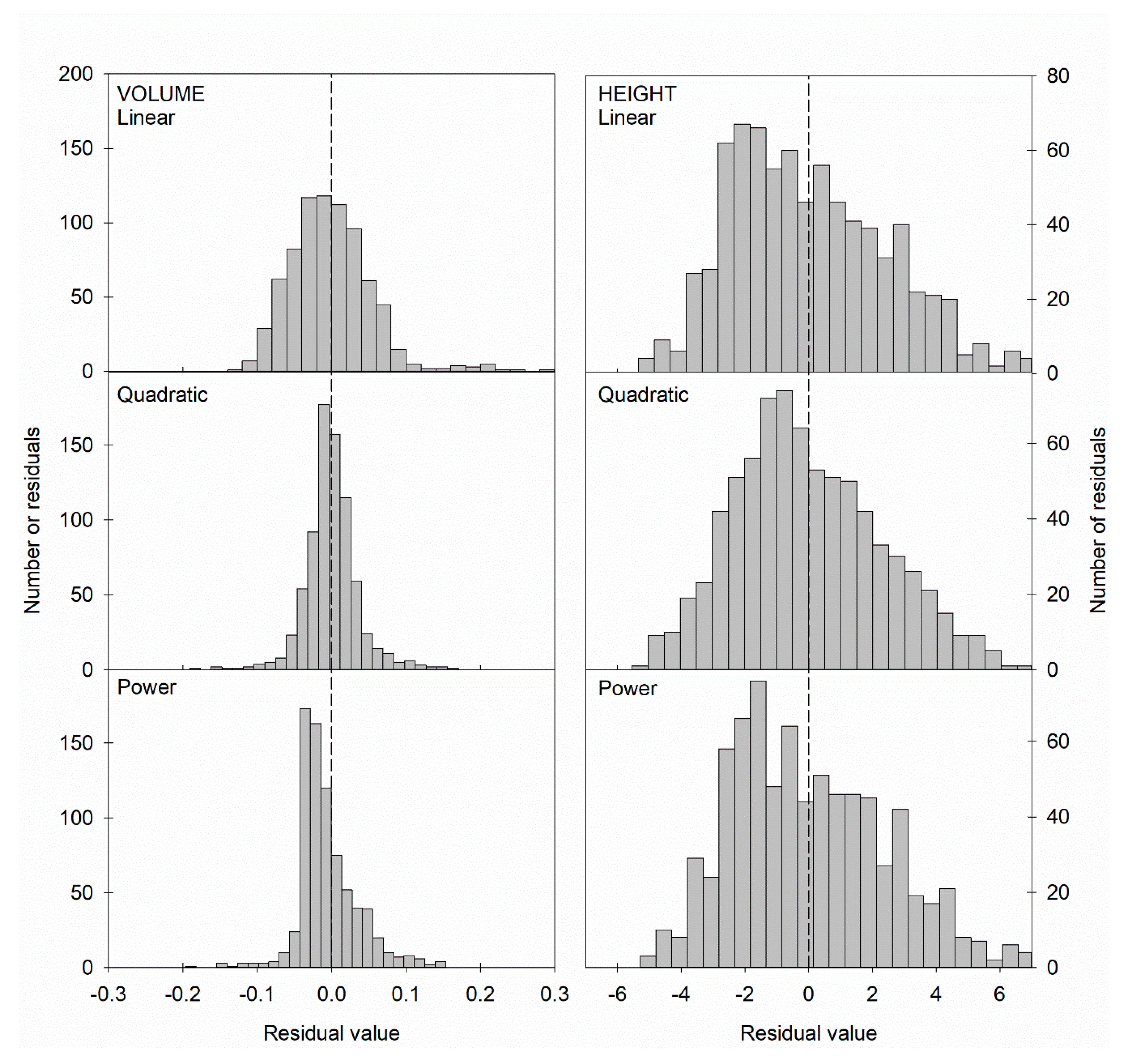
| Equation | Y0 | A | B | R2adj | SEestimate |
|---|---|---|---|---|---|
| Volume | |||||
| Linear | −0.5038 ** ± 0.0071 | 0.0379 ** ± 0.0003 | - | 0.9418 | 0.0577 |
| Quadratic | −0.0308 * ± 0.0013 | −0.0088 ** ± 0.0013 | 0.0011 ** ± 2.9·10−5 | 0.9782 | 0.0353 |
| Power | - | 0.000056 ** ± 3.7·10−6 | 2.7473 ** ± 0.0196 | 0.9709 | 0.0408 |
| Height | |||||
| Linear | 5.0331 ** ± 0.2994 | 0.6935 ** ± 0.0143 | - | 0.7521 | 2.4357 |
| Quadratic | 0.4433 ± 0.9425 | 1.1472 ** ± 0.0896 | −0.0102 ** ± 0.0020 | 0.7600 | 2.3966 |
| Power | - | 2.1061 ** ± 0.0982 | 0.7356** ± 0.0151 | 0.7575 | 2.4092 |
| Variable | Plot 1 | Plot 2 | Plot 3 | All Plots |
|---|---|---|---|---|
| Number of trees | 10 | 10 | 11 | 31 |
| DBH (cm) | 24.93 ± 7.543a | 24.30 ± 3.41a | 24.37 ± 3.61a | 24.83 ± 4.98 |
| Height (m) | 25.25 ± 5.54a | 21.58 ± 4.02b | 20.26 ± 2.16c | 22.01 ± 4.29 |
| Slenderness index (m/m) | 104.1 ± 16.9a | 92.3 ± 7.5b | 87.6 ± 8.2c | 93.4 ± 13.3 |
| Tree volume (m3) | 0.519 ± 0.405a | 0.454 ± 0.145a | 0.337 ± 0.139b | 0.395 ± 0.257 |
| Tree basal area (m2) | 0.053 ± 0.031a | 0.053 ± 0.011a | 0.044 ± 0.013a | 0.046 ± 0.020 |
| Equation | Average Bias | MAD | RMSE | U | ME | e* α = 0.05 | e* α = 0.20 |
|---|---|---|---|---|---|---|---|
| Volume | |||||||
| Linear | 0.032 | 0.041 | 0.056 | 0.287 | 0.918 | 0.101 | 0.069 |
| Quadratic | 0.067 | 0.026 | 0.034 | 0.176 | 0.969 | 0.062 | 0.042 |
| Power | 0.011 | 0.029 | 0.037 | 0.188 | 0.965 | 0.066 | 0.045 |
| Height | |||||||
| Linear | −0.003 | 2.058 | 2.547 | 0.555 | 0.691 | 4.610 | 3.134 |
| Quadratic | 0.002 | 2.046 | 2.531 | 0.552 | 0.695 | 4.582 | 3.115 |
| Power | 0.000 | 2.053 | 2.534 | 0.553 | 0.694 | 4.588 | 3.119 |
| DBH (cm) | Volume (m3 stem−1) | Height (m) | ||||
|---|---|---|---|---|---|---|
| Linear | Quadratic | Power | Linear | Quadratic | Power | |
| 10 | 0.000–0.009 | 0.000–0.062 | 0.000–0.113 | 7.10–16.84 | 6.09–15.68 | 6.64–16.27 |
| 15 | 0.000–0.180 | 0.014–0.155 | 0.014–0.177 | 10.56–20.31 | 10.55–20.13 | 10.62–20.26 |
| 20 | 0.139–0.370 | 0.163–0.304 | 0.129–0.293 | 14.03–23.77 | 14.49–24.09 | 14.26–23.90 |
| 25 | 0.328–0.559 | 0.366–0.507 | 0.308–0.471 | 17.50–27.24 | 17.93–27.52 | 17.66–27.30 |
| 30 | 0.518–0.749 | 0.625–0.766 | 0.561–0.725 | 20.97–30.71 | 20.86–30.44 | 20.89–30.53 |
| 35 | 0.707–0.938 | 0.938–1.079 | 0.900–1.064 | 24.43–34.18 | 23.27–32.86 | 23–98-33.61 |
| 40 | 0.897–1.128 | 1.307–1.448 | 1.335–1.499 | 27.90–37.64 | 25.18–34.76 | 26.95–36.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, R.; Blanco, J.A. Empowering Forest Owners with Simple Volume Equations for Poplar Plantations in the Órbigo River Basin (NW Spain). Forests 2021, 12, 124. https://doi.org/10.3390/f12020124
Blanco R, Blanco JA. Empowering Forest Owners with Simple Volume Equations for Poplar Plantations in the Órbigo River Basin (NW Spain). Forests. 2021; 12(2):124. https://doi.org/10.3390/f12020124
Chicago/Turabian StyleBlanco, Roberto, and Juan A. Blanco. 2021. "Empowering Forest Owners with Simple Volume Equations for Poplar Plantations in the Órbigo River Basin (NW Spain)" Forests 12, no. 2: 124. https://doi.org/10.3390/f12020124
APA StyleBlanco, R., & Blanco, J. A. (2021). Empowering Forest Owners with Simple Volume Equations for Poplar Plantations in the Órbigo River Basin (NW Spain). Forests, 12(2), 124. https://doi.org/10.3390/f12020124