
Variations in Soil Properties and CO2 Emissions of a Temperate Forest Gully Soil along a Topographical Gradient

 ,
,  ,
,  ,
,  and
and 
Abstract
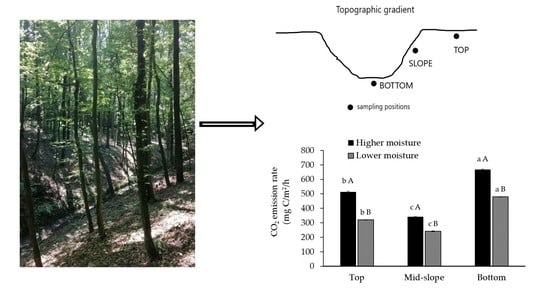
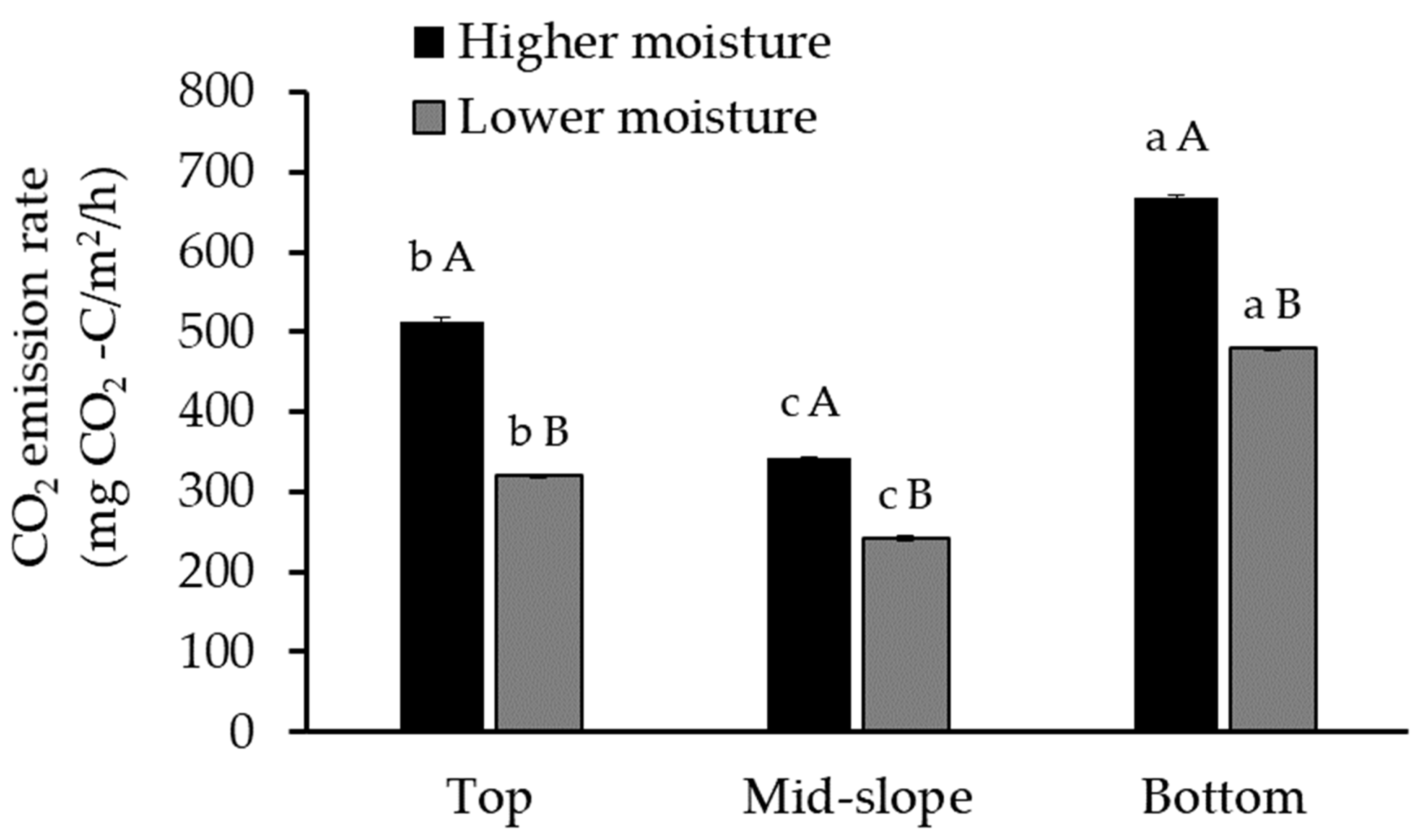
:
1. Introduction
- (1)
- Higher clay fractions in the gully bottom, because of downward runoff;
- (2)
- Higher SOC stocks in the gully bottom due to absorption by clay particles;
- (3)
- Higher total N contents in the gully bottom because of a higher SOC and the absence of vegetation;
- (4)
- A higher microbial biomass in locations with a higher C and N availability;
- (5)
- Generally higher soil water contents which will either stimulate or decrease soil microbial activity, depending on the range of soil moisture values experienced.
2. Materials and Methods
2.1. Soil Physicochemical Characterization
2.2. Soil Microbial Parameters
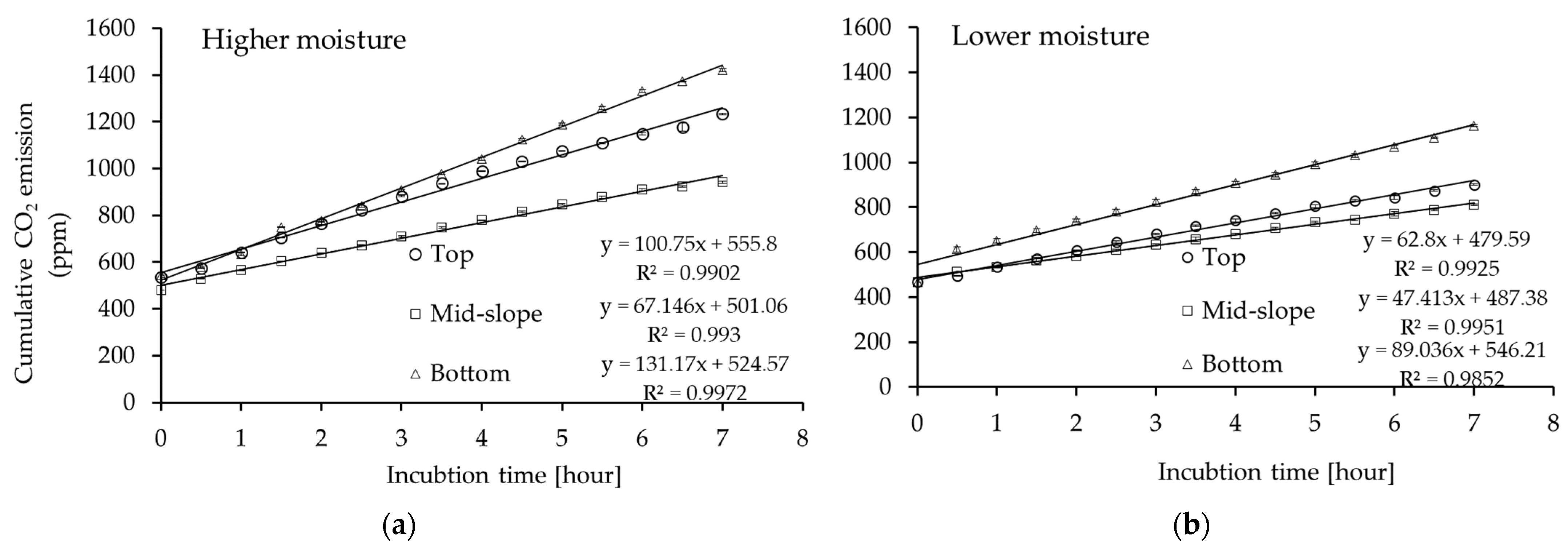
2.3. CO2 Emission Measurements
- kCO2 = gas constant at 273.15 K (0.536 µg C/µL)
- T—air temperature (°K)
- V—chamber headspace volume (dm3)
- A—area of the soil in the chamber (m2)
- S—CO2 concentration change in the chamber based on the slope (dc/dt; ppm/min)
2.4. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of the Forest Gully Soil
3.2. CO2 Emissions from Forest Gully Soil
3.3. Analysis of Interrelationships
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| All Samples | CO2 Emission | DHA | CAT | BR | Cmic | Clay | Silt | Sand | pH | SOC | Ntot. | C:N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CO2 emission | 1 | |||||||||||
| DHA | 0.630 | 1 | ||||||||||
| CAT | 0.877 * | 0.495 | 1 | |||||||||
| BR | 0.792 * | 0.591 | 0.443 | 1 | ||||||||
| Cmic | 0.851 * | 0.317 | 0.830 * | 0.606 | 1 | |||||||
| clay | −0.089 | −0.731 * | 0.169 | −0.322 | 0.336 | 1 | ||||||
| silt | −0.224 | −0.802 * | 0.043 | −0.424 | 0.213 | 0.990 * | 1 | |||||
| sand | 0.205 | 0.792 * | −0.062 | 0.409 | −0.231 | −0.993 * | −0.999 * | 1 | ||||
| pH | 0.991 * | 0.645 | 0.847 * | 0.844 * | 0.852 * | −0.101 | −0.235 | 0.215 | 1 | |||
| SOC | 0.854 * | 0.216 | 0.880 * | 0.544 | 0.796 * | 0.330 | 0.203 | −0.221 | 0.831 * | 1 | ||
| Ntot. | 0.823 * | 0.138 | 0.836 * | 0.560 | 0.822 * | 0.410 | 0.286 | −0.304 | 0.813 * | 0.975 * | 1 | |
| C:N | −0.481 | 0.160 | −0.499 | −0.383 | −0.680 * | −0.596 | −0.518 | 0.530 | −0.514 | −0.643 | −0.792 * | 1 |
References
- Canadell, J.G.; Raupach, M.R. Managing forests for climate change mitigation. Science 2008, 320, 1456–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Peng, Y.; Thomas, S.C.; Tian, D. Forest management and soil respiration: Implications for carbon sequestration. Environ. Rev. 2008, 16, 93–111. [Google Scholar] [CrossRef]
- Meixner, F.X.; Eugster, W. Effects of landscape pattern and topography on emissions and transport. In Integrating Hydrology, Ecosystem Dynamics, and Biogeochemistry in Complex Landscapes; Tenhunen, J.D., Kabat, P., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 1999; Volume 3, pp. 143–175. [Google Scholar]
- Brito, L.D.; Marques, J.; Pereira, G.T.; Souza, Z.M.; La Scala, N. Soil CO2 emission of sugarcane fields as affected by topography. Sci. Agric. 2009, 66, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Qu, B.; Aho, K.S.; Li, C.; Kang, S.; Sillanpää, M.; Yan, F.; Raymond, P.A. Greenhouse gases emissions in rivers of the Tibetan Plateau. Sci Rep. 2017, 7, 1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse gas emissions from soils—A review. Chem. Erde. 2016, 76, 327–352. [Google Scholar] [CrossRef] [Green Version]
- Marty, C.; Piquette, J.; Morin, H.; Bussières, D.; Thiffault, N.; Houle, D.; Bradley, R.L.; Simpson, M.J.; Ouimet, R.; Paré, M.C. Nine years of in situ soil warming and topography impact the temperature sensitivity and basal respiration rate of the forest floor in a Canadian boreal forest. PLoS ONE 2019, 14, e0226909. [Google Scholar] [CrossRef] [Green Version]
- Arias-Navarro, C.; Díaz-Pinés, E.; Klatt, S.; Brandt, P.; Rufino, M.C.; Butterbach-Bahl, K.; Verchot, L.V. Spatial variability of soil N2O and CO2 fluxes in different topographic positions in a tropical montane forest in Kenya. J. Geophys. Res. Biogeosci. 2017, 122, 514–527. [Google Scholar] [CrossRef]
- Dagar, J.C.; Singh, A.K. Ravine Lands: Greening for Livelihood an Environmental Security; Springer Nature Singapore Pte Ltd.: Singapore, 2018; p. 4. [Google Scholar]
- Gawrysiak, L.; Harasimiuk, M. Spatial diversity of gully density of the Lublin Upland and Roztocze Hills (SE Poland). Ann. UMCS 2012, 67, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Zgłobicki, W.; Poesen, J.; Cohen, M.; Del Monte, M.; García-Ruiz, J.M.; Ionita, I.; Niacsu, L.; Machová, Z.; Martín-Duque, J.F.; Nadal-Romero, E.; et al. The Potential of Permanent Gullies in Europe as Geomorphosites. Geoheritage 2019, 11, 217–239. [Google Scholar] [CrossRef]
- Vanwalleghem, T.; Van Den Eeckhaut, M.; Poesen, J.; Deckers, J.; Nachtergaele, J.; Van Oost, K.; Slenters, C. Characteristics and controlling factors of old gullies under forest in a temperate humid climate: A case study from the Meerdaal Forest (Central Belgium). Geomorphology 2003, 56, 15–29. [Google Scholar] [CrossRef]
- Baran-Zgłobicka, B.; Zgłobicki, W. Mosaic landscapes of SE Poland: Should we preserve them? Agrofor. Syst. 2012, 85, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Langohr, R.; Sanders, J. The Belgium Loessbelt in the last 20,000 years: Evolution of soils andrelief in the Zonien Forest. In Soils and Quaternary Landscape Evolution; Boardman, J., Ed.; Wiley: Chichester, UK, 1985; pp. 359–371. [Google Scholar]
- Tsui, C.C.; Chen, Z.S.; Hsieh, C.F. Relationships between soil properties and slope position in a lowland rain forest of southern Taiwan. Geoderma 2004, 123, 131–142. [Google Scholar] [CrossRef]
- Hao, Y.; Chang, Q.; Li, L.; Wei, X. Impacts of landform, land use and soil type on soil chemical properties and enzymatic activities in a Loessial Gully watershed. Soil Res. 2014, 52, 453–462. [Google Scholar] [CrossRef]
- Shi, W.-Y.; Du, S.; Morina, J.C.; Guan, J.-H.; Wang, K.-B.; Ma, M.-G.; Yamanaka, N.; Tateno, R. Physical and biogeochemical controls on soil respiration along a topographical gradient in a semiarid forest. Agric. For. Meteorol. 2017, 247, 1–11. [Google Scholar] [CrossRef]
- Konda, R.; Ohta, S.; Ishizuka, S.; Heriyanto, J.; Wicaksono, A. Seasonal changes in the spatial structures of N2O, CO2, and CH4 fluxes from Acacia mangium plantation soils in Indonesia. Soil Biol. Biochem. 2010, 42, 1512–1522. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, J.; Liu, G.B.; Yamanaka, N. Soil properties in natural grassland, Caragana korshinskii planted shrubland, and Robinia pseudoacacia planted forest in gullies on the hilly Loess Plateau, China. Catena 2014, 119, 116–124. [Google Scholar] [CrossRef]
- Wolf, K.; Flessa, H.; Veldkamp, E. Atmospheric methane uptake by tropical montane forest soils and the contribution of organic layers. Biogeochemistry 2012, 111, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.A.; Ball, T.; Conen, F.; Dobbie, K.E.; Massheder, J.; Rey, A. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2003, 54, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.; Hernandez, T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal. 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Garcia, C.; Hernandez, T. Biological and biochemical indicators in derelict soils subject to erosion. Soil Biol. Biochem. 1997, 29, 171–177. [Google Scholar] [CrossRef]
- Błońska, E.; Lasota, J.; Zwydak, M. The relationship between soil properties, enzyme activity and land use. Forest Res. Pap. 2017, 78, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Wolińska, A.; Stępniewska, Z. Dehydogenase activity in soil ecosystem. In Dehydrogenases; Canuto, R.A., Ed.; Intech: Rijeka, Croatia, 2012; pp. 183–210. [Google Scholar]
- Moeskops, B.; Sukristiyonubowo; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil microbial communities and activities under intensive organic and conventional vegetable farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, P.; Kong, C. Urease, invertase, dehydrogenase and polyphenoloxidase activities in paddy soils influenced by allelophatic rice variety. Eur. J. Soil Biol. 2009, 45, 436–444. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Camiña, F.; Leirós, M.; Gil-Sotres, F. An improved method to measure catalase activity in soils. Soil Boil. Biochem. 1999, 31, 483–485. [Google Scholar] [CrossRef]
- Garcıa-Gil, J.C.; Kobza, J.; Soler-Rovira, P.; Javoreková, S.; García-Gil, J.C. Soil microbial and enzyme activities response to pollution near an aluminium smelter. CLEAN Soil Air Water 2013, 41, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.-H. Microbial eco-physiological indicators to asses soil quality. Agric. Ecosyst. Environ. 2003, 98, 285–293. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, J.; Zhang, L.; Ren, C.; Han, X.; Yang, G.; Doughty, R.; Deng, J. Understory plants regulate soil respiration through changes in soil enzyme activity and microbial C, N, and P stoichiometry following afforestation. Forests 2018, 9, 436. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, Y.; Li, Y. Soil type and a labile C addition regime control the temperature sensitivity of soil C and N mineralization more than N addition in wetland soils in China. Atmosphere 2020, 11, 1043. [Google Scholar] [CrossRef]
- Krzyżewska, A.; Wereski, S.; Nowosad, M. Thermal variability in the Lublin Region during the frost wave in January 2017. Ann. UMCS Sec. B 2019, 74, 217–229. [Google Scholar] [CrossRef] [Green Version]
- Bhat, S.A.; Dar, M.U.D.; Meena, R.S. Soil erosion and management strategies. In Sustainable Management of Soil and Environment, 1st ed.; Meena, R.S., Kumar, S., Bohra, J.S., Jat, M.L., Eds.; Springer Nature Pte Ltd.: Singapore, 2019; p. 91. [Google Scholar]
- Bieganowski, A.; Chojecki, T.; Ryżak, M.; Sochan, A.; Lamorski, K. Methodological Aspects of Fractal Dimension Estimation on the Basis of Particle Size Distribution. Vadose Zone J. 2013, 12, 12. [Google Scholar] [CrossRef]
- Bieganowski, A.; Ryżak, M.; Sochan, A.; Barna, G.; Hernádi, H.; Beczek, M.; Polakowski, C.; Makó, A. Laser diffractometry in the measurements of soil and sediment particle size distribution. Adv. Agron. 2018, 151, 215–279. [Google Scholar] [CrossRef]
- You, G.; Zhang, Z.; Zhang, R. Temperature adaptability of soil respiration in short-term incubation experiments. J. Soil. Sediment. 2019, 19, 557–565. [Google Scholar] [CrossRef]
- Walkiewicz, A.; Brzezińska, M.; Bieganowski, A.; Sas-Paszt, L.; Frąc, M. Early response of soil microbial biomass and activity to biofertilizer application in degraded Brunic Arenosol and Abruptic Luvisol of contrasting textures. Agronomy 2020, 10, 1347. [Google Scholar] [CrossRef]
- Trevors, J. Rapid gas chromatographic method to measure H2O2 oxidoreductase (catalase) activity in soil. Soil Boil. Biochem. 1984, 16, 525–526. [Google Scholar] [CrossRef]
- Walkiewicz, A.; Brzezińska, M. Interactive effects of nitrate and oxygen on methane oxidation in three different soils. Soil Boil. Biochem. 2019, 133, 116–118. [Google Scholar] [CrossRef]
- Casida, L.E.; Klein, D.A.; Santoro, T. Soil Dehydrogenase Activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Anderson, J.; Domsch, K. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Boil. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Šimek, M.; Kalčík, J. Carbon and nitrate utilization in soils: The effect of long-term fertilization on potential denitrification. Geoderma 1998, 83, 269–280. [Google Scholar] [CrossRef]
- Chojnicki, B.H.; Michalak, M.; Acosta, M.; Juszczak, R.; Augustin, J.; Drösler, M.; Olejnik, J. Measurements of carbon dioxide fluxes by chamber method at the Rzecin wetland ecosystem, Poland. Pol. J. Environ. Stud. 2010, 19, 283–291. [Google Scholar]
- Flessa, H.; Wild, U.; Klemisch, M.; Pfadenhauer, J. Nitrous oxide and methane fluxes from organic soils under agriculture. Eur. J. Soil Sci. 1998, 49, 327. [Google Scholar]
- Patton, N.R.; Lohse, K.A.; Seyfried, M.S.; Godsey, S.E.; Parsons, S.B. Topographic controls of soil organic carbon on soil-mantled landscapes. Sci. Rep. 2019, 9, 6390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, K.; Anusontpornperm, S.; Kheoruenromne, I.; Thanachit, S. Carbon sequestration in relation to topographic aspects and land use in northeast of Thailand. Int. J. Environ. Clim. Chang. 2018, 8, 118–137. [Google Scholar] [CrossRef]
- Hou, G.; Delang, C.O.; Lu, X. Afforestation changes soil organic carbon stocks on sloping land: The role of previous land cover and tree type. Ecol. Eng. 2020, 152, 105860. [Google Scholar] [CrossRef]
- Zhang, X.; Adamowski, J.F.; Liu, C.; Zhou, J.; Zhu, G.; Dong, X.; Cao, J.; Feng, Q. Which slope aspect and gradient provides the best afforestation-driven soil carbon sequestration on the China’s Loess Plateau? Ecol. Eng. 2020, 147, 105782. [Google Scholar] [CrossRef]
- Feller, C.; Beare, M.H. Physical control of soil organic matter dynamics in the Tropics. Geoderma 1997, 79, 69–116. [Google Scholar] [CrossRef]
- Siqueira Neto, M.; Scopel, E.; Corbeels, M.; Nunes Cardoso, A.; Douzet, J.M.; Feller, C.; Piccolo, M.C.; Cerri, C.C.; Bernoux, M. Soil carbon stocks under no-tillage mulch-based cropping systems in the Brazilian Cerrado: An on-farm synchronic assessment. Soil Till. Res. 2010, 110, 187–195. [Google Scholar] [CrossRef]
- Sattler, D.; Murray, L.T.; Kirchner, A.; Lindner, A. Influence of soil and topography on aboveground biomass accumulation and carbon stocks of afforested pastures in South East Brazil. Ecol. Eng. 2014, 73, 126–131. [Google Scholar] [CrossRef]
- Li, T.; Zhang, H.; Wang, X.; Cheng, S.; Fang, H.; Liu, G.; Yuan, W. Soil erosion affects variations of soil organic carbon and soil respiration along a slope in Northeast China. Ecol. Process 2019, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Churchill, R.R. Aspect-related differences in badlands slope morphology. Ann. Assoc. Am. Geogr. 1981, 71, 374–388. [Google Scholar]
- Parsons, A.J. Hillslope Form; Routledge: London, UK, 1988. [Google Scholar]
- Regmi, N.R.; McDonald, E.V.; Rasmussen, C. Hillslope response under variable microclimate. Earth Surf. Proc. Land 2019, 44, 2615–2627. [Google Scholar] [CrossRef]
- Zhang, Y.; Mu, C.C.; Zheng, T.; Li, N.N. Ecosystem carbon storage of natural secondary birch forests in Xiaoxing’an Mountains of China. J. Beijing For. Univ. 2015, 37, 38–47. [Google Scholar]
- Wang, W.; Wang, Y.; Hoch, G.; Wang, Z.; Gu, J. Linkage of root morphology to anatomy with increasing nitrogen availability in six temperate tree species. Plant Soil 2018, 425, 189–200. [Google Scholar] [CrossRef]
- Gressel, N.; Inbar, Y.; Singer, A.; Chen, Y. Chemical and spectroscopic properties of leaf litter and decomposed organic matter in the Carmel Range, Israel. Soil Biol. Biochem. 1995, 27, 23–31. [Google Scholar] [CrossRef]
- Tsai, S.-H.; Selvam, A.; Yang, S.-S. Microbial diversity of topographical gradient profiles in Fushan forest soils of Taiwan. Ecol. Res. 2007, 22, 814–824. [Google Scholar] [CrossRef]
- Schlaghamerský, J.; Devetter, M.; Hánel, L.; Tajkovský, K.; Starý, J.; Tuf, I.H.; Pizl, V. Soil fauna across Central European sandstone ravines with temperature inversion: From cool and shady to dry and hot places. App. Soil Ecol. 2014, 83, 30–38. [Google Scholar] [CrossRef]
- Blättermann, M.; Frechen, M.; Gass, A.; Hoelzmann, P.; Parzinger, H.; Schütt, B. Late Holocene landscape reconstruction in the Land of Seven Rivers, Kazakhstan. Quat. Int. 2012, 251, 42–51. [Google Scholar] [CrossRef]
- Whitmyer, R.W.; Blake, G.R. Influence of silt and clay on the physical performance of sand-soil mixtures. Agron. J. 1989, 81, 5–12. [Google Scholar] [CrossRef]
- Kosheleva, N.E.; Kasimov, N.S.; Vlasow, D.V. Factors of the accumulation of heavy metals and metalloids at geochemical barriers in urban soils. Eurasian Soil Sci. 2015, 48, 476–492. [Google Scholar] [CrossRef]
- Quilchano, C.; Marañón, T. Dehydrogenase activity in Mediterranean forest soils. Biol. Fertil. Soils 2002, 35, 102–107. [Google Scholar] [CrossRef]
- Moreno-de las Heras, M. Development of soil physical structure and biological functionality in mining spoils affected by soil erosion in a Mediterranean-Continental environment. Geoderma 2009, 149, 249–256. [Google Scholar] [CrossRef]
- Alef, K.; Nannipieri, P. Catalase activity. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 362–363. [Google Scholar]
- Wang, B.; Wang, Y.; Wang, L. The effects of erosional topography on soil properties in a Pinus massoniana forest in southern China. J. Soil Water Conserv. 2017, 72, 36–44. [Google Scholar] [CrossRef]
- Clay, G.D.; Dixon, S.; Evans, M.G.; Rowson, J.G.; Worrall, F. Carbon dioxide fluxes and DOC concentrations of eroding blanket peat gullies. Earth Surf. Process. Land. 2012, 37, 562–571. [Google Scholar] [CrossRef]
- Soosaar, K.; Mander, Ü.; Maddison, M.; Kanal, A.; Kull, A.; Lőhmus, K.; Truu, J.; Augustin, J. Dynamics of gaseous nitrogen and carbon fluxes in riparian alder forests. Ecol. Eng. 2011, 37, 40–53. [Google Scholar] [CrossRef]
- Kang, S.; Doh, S.; Lee, D.; Lee, D.; Jin, V.L.; Kimball, J.S. Topographic and climatic controls on soil respiration in six temperate mixed-hardwood forest slopes, Korea. Glob. Change Biol. 2003, 9, 1427–1437. [Google Scholar] [CrossRef] [Green Version]
- Oh, N.-H.; Kim, H.-S.; Richter, D.D. What regulates soil CO2 concentrations? A modeling approach to CO2 diffusion in deep soil profiles. Environ. Eng. Sci. 2005, 22, 38–45. [Google Scholar] [CrossRef]
- Gliński, J.; Stępniewski, W. Soil Aeration and Its Role for Plants; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]
- Schaufler, G.; Kitzler, B.; Schindlbacher, A.; Skiba, U.; Sutton, M.A.; Zechmeister-Boltenstern, S. Greenhouse gas emissions from European soils under different land use: Effects of soil moisture and temperature. Eur. J. Soil Sci. 2010, 61, 683–696. [Google Scholar] [CrossRef]
- Brzezińska, M.; Stępniewska, Z.; Stępniewski, W. Soil oxygen status and dehydrogenase activity. Soil Biol. Biochem. 1998, 30, 1783–1790. [Google Scholar] [CrossRef]
- Cools, N.; Vesterdal, L.; De Vos, B.; Vanguelova, E.; Hansen, K. Tree species is the major factor explaining C:N ratios in European forest soils. Forest Ecol. Manag. 2014, 311, 3–16. [Google Scholar] [CrossRef]
- Zhang, J.; Quine, T.A.; Ni, S.; Ge, F. Stocks and dynamics of SOC in relation to soil redistribution by water and tillage erosion. Glob. Change Biol. 2006, 12, 1834–1841. [Google Scholar] [CrossRef]
- Li, Z.W.; Liu, C.; Dong, Y.T.; Chang, X.F.; Nie, X.D.; Lin, L.; Xiao, H.B.; Lu, Y.M.; Zeng, G.M. Response of soil organic carbon and nitrogen stocks to soil erosion and land use types in the Loess hilly-gully region of China. Soil Tillage Res. 2017, 166, 1–9. [Google Scholar] [CrossRef]
- Li, Z.; Xiao, H.; Tang, Z.; Huang, J.; Nie, X.; Huang, B.; Ma, W.; Lu, Y.; Zeng, G. Microbial responses to erosion-induced soil physico-chemical property changes in the hilly red soil region of southern China. Eur. J. Soil Biol. 2015, 71, 37–44. [Google Scholar] [CrossRef]
- Seaton, F.M.; George, P.B.L.; Lebron, I.; Jones, D.L.; Creer, S.; Robinson, D.A. Soil textural heterogeneity impacts bacterial but not fungal diversity. Soil Biol. Biochem. 2020, 144, 107766. [Google Scholar] [CrossRef]
- Chau, J.F.; Bagtzoglou, A.C.; Willig, M.R. The effect of soil texture on richness and diversity of bacterial communities. Environ. Forensics 2011, 12, 333–341. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Dohrmann, A.B.; Christensen, B.T.; Tebbe, C.C. Bacterial preferences for specific soil particle size fractions revealed by community analyses. Front. Microbiol. 2018, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Sessitsch, A.; Weilharter, A.; Gerzabek, M.H.; Kirchmann, H.; Kandeler, E. Microbial population structures in soil particle size fractions of a long-term fertilizer field experiment. Appl. Environ. Microbiol. 2001, 67, 4215–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Tested Positions | Top | Mid-Slope | Bottom |
|---|---|---|---|
| SOC (%) | 1.28 ± 0.046 (a) | 1.27 ± 0.036 (a) | 1.70 ± 0.128 (b) |
| Ntot (%) | 0.09 ± 0.003 (a) | 0.10 ± 0.002 (a) | 0.13 ± 0.012 (b) |
| C:N Ratio | 13.99 ± 0.30 (b) | 13.40 ± 0.45 (ab) | 12.70 ± 0.64 (a) |
| pH | 5.44 ± 0.06 (b) | 4.89 ± 0.15 (a) | 6.17 ± 0.08 (c) |
| Texture | loamy sand | sandy loam | sandy loam |
| Clay (%) | 1.70 ± 0.11 (a) | 3.51 ± 0.09 (b) | 3.51 ± 0.41 (b) |
| Silt (%) | 19.93 ± 1.14 (a) | 38.30 ± 1.06 (c) | 34.07 ± 0.97 (b) |
| Sand (%) | 78.37 ± 1.25 (c) | 58.26 ± 1.08 (a) | 62.42 ± 1.37 (b) |
| Silt:Clay Ratio | 11.72 ± 0.13 (c) | 10.93 ± 0.10 (b) | 9.80 ± 0.83 (a) |
| Tested Positions | Top | Mid-Slope | Bottom |
|---|---|---|---|
| CAT (µmol O2/g/min) | 10.77 ± 2.09 (a) | 10.64 ± 0.98 (a) | 11.26 ± 0.88 (b) |
| DHA (µg TPF/g/20 h) | 2.74 ± 0.35 (b) | 1.00 ± 0.25 (a) | 2.32 ± 0.43 (b) |
| BR (µg CO2-C/g/h) | 6.91 ± 0.82 (ab) | 5.41 ± 0.79 (a) | 7.51 ± 0.60 (b) |
| Cmic (mg C/g) | 0.97 ± 0.02 (a) | 0.97 ± 0.01 (a) | 1.15 ± 0.06 (b) |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Ratio | p-Value |
|---|---|---|---|---|---|
| Factor | |||||
| Position | 239,717 | 2 | 119,859 | 15,472.1 | 0.00 |
| Moisture | 116,850 | 1 | 116,850 | 15,083.8 | 0.00 |
| Position × Moisture | 8259 | 2 | 4129 | 533.1 | 0.00 |
| Error | 93 | 12 | 8 |
| DHA | CAT | BR | Cmic | Clay | Silt | Sand | pH | SOC | Ntot | C:N | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CO2 Emission | 0.63 | 0.877 * | 0.792 * | 0.851 * | −0.089 | −0.224 | 0.205 | 0.991 * | 0.854 * | 0.823 * | −0.481 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walkiewicz, A.; Bulak, P.; Brzezińska, M.; Khalil, M.I.; Osborne, B. Variations in Soil Properties and CO2 Emissions of a Temperate Forest Gully Soil along a Topographical Gradient. Forests 2021, 12, 226. https://doi.org/10.3390/f12020226
Walkiewicz A, Bulak P, Brzezińska M, Khalil MI, Osborne B. Variations in Soil Properties and CO2 Emissions of a Temperate Forest Gully Soil along a Topographical Gradient. Forests. 2021; 12(2):226. https://doi.org/10.3390/f12020226
Chicago/Turabian StyleWalkiewicz, Anna, Piotr Bulak, Małgorzata Brzezińska, Mohammad I. Khalil, and Bruce Osborne. 2021. "Variations in Soil Properties and CO2 Emissions of a Temperate Forest Gully Soil along a Topographical Gradient" Forests 12, no. 2: 226. https://doi.org/10.3390/f12020226
APA StyleWalkiewicz, A., Bulak, P., Brzezińska, M., Khalil, M. I., & Osborne, B. (2021). Variations in Soil Properties and CO2 Emissions of a Temperate Forest Gully Soil along a Topographical Gradient. Forests, 12(2), 226. https://doi.org/10.3390/f12020226