Genetic Diversity and Structure of Apomictic and Sexually Reproducing Lindera Species (Lauraceae) in Japan

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Focal Species
2.2. Field Collections and DNA Extraction
2.3. Multiplexed ISSR Genotyping by Sequencing (MIG-Seq) Analysis
2.4. Data Analyses
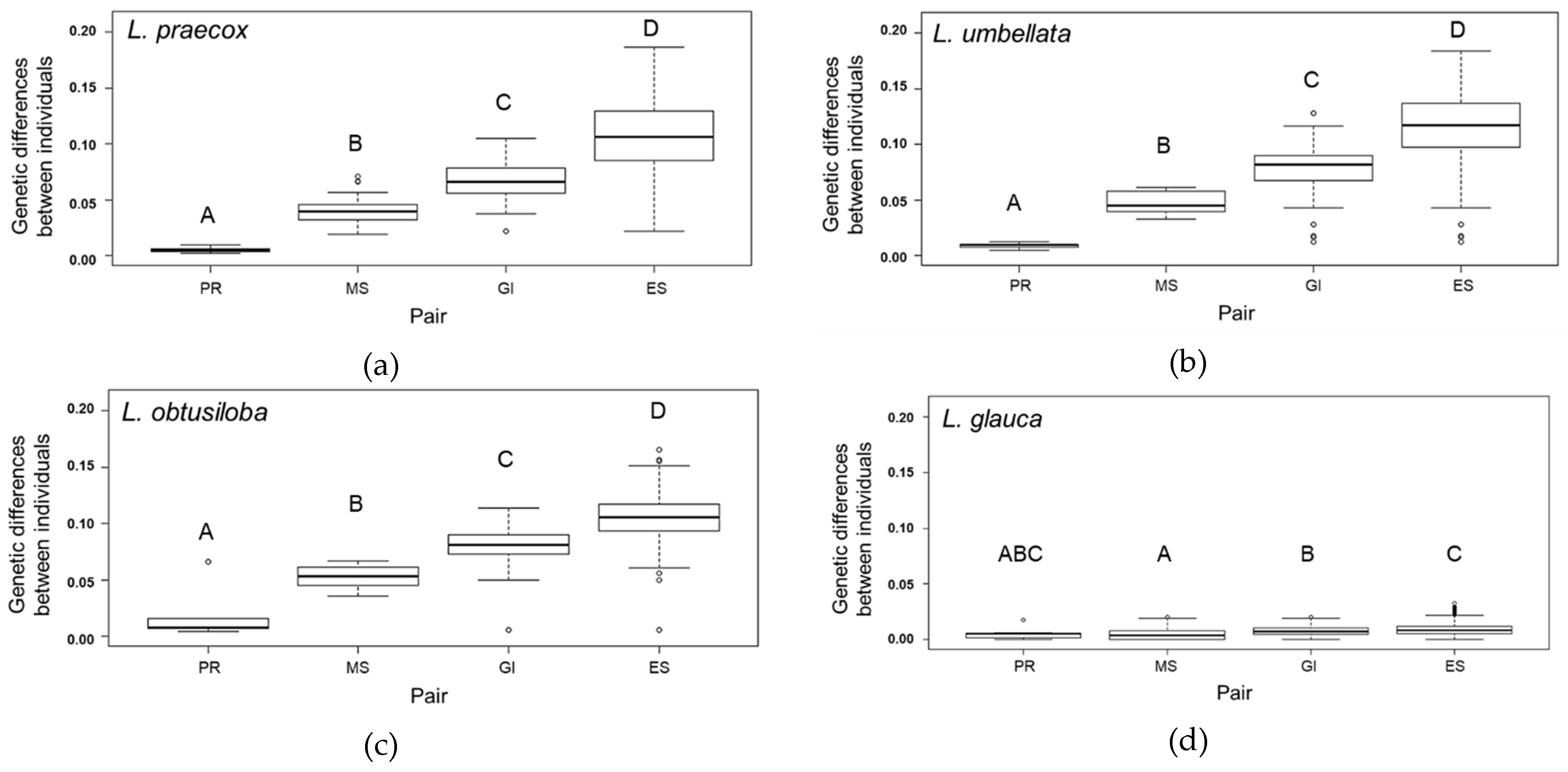
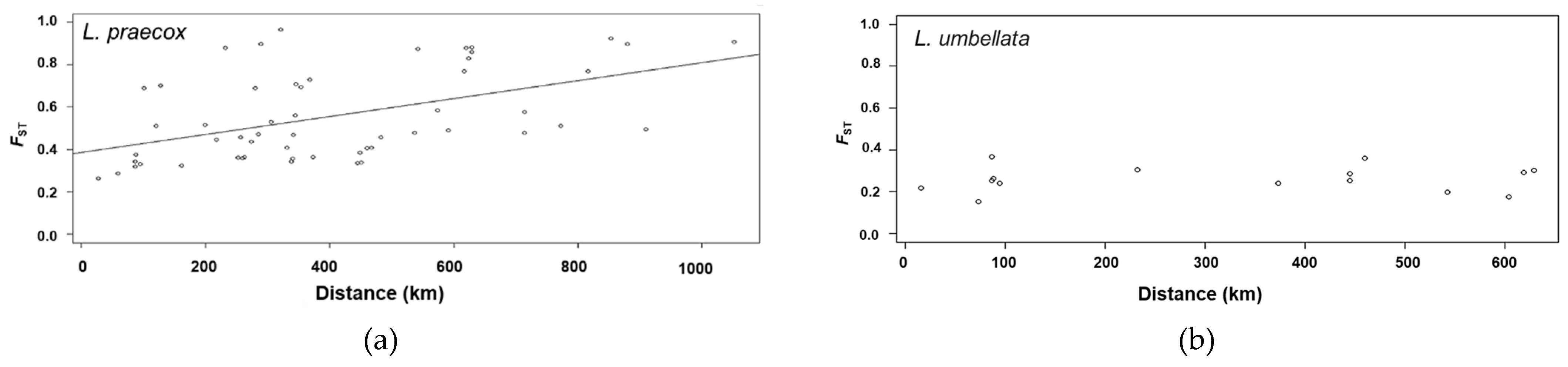
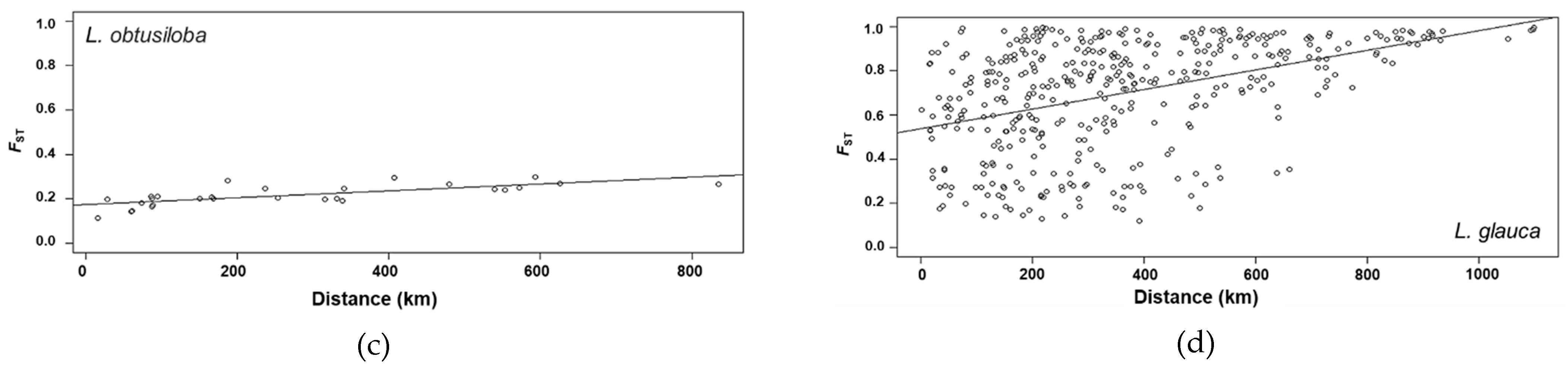
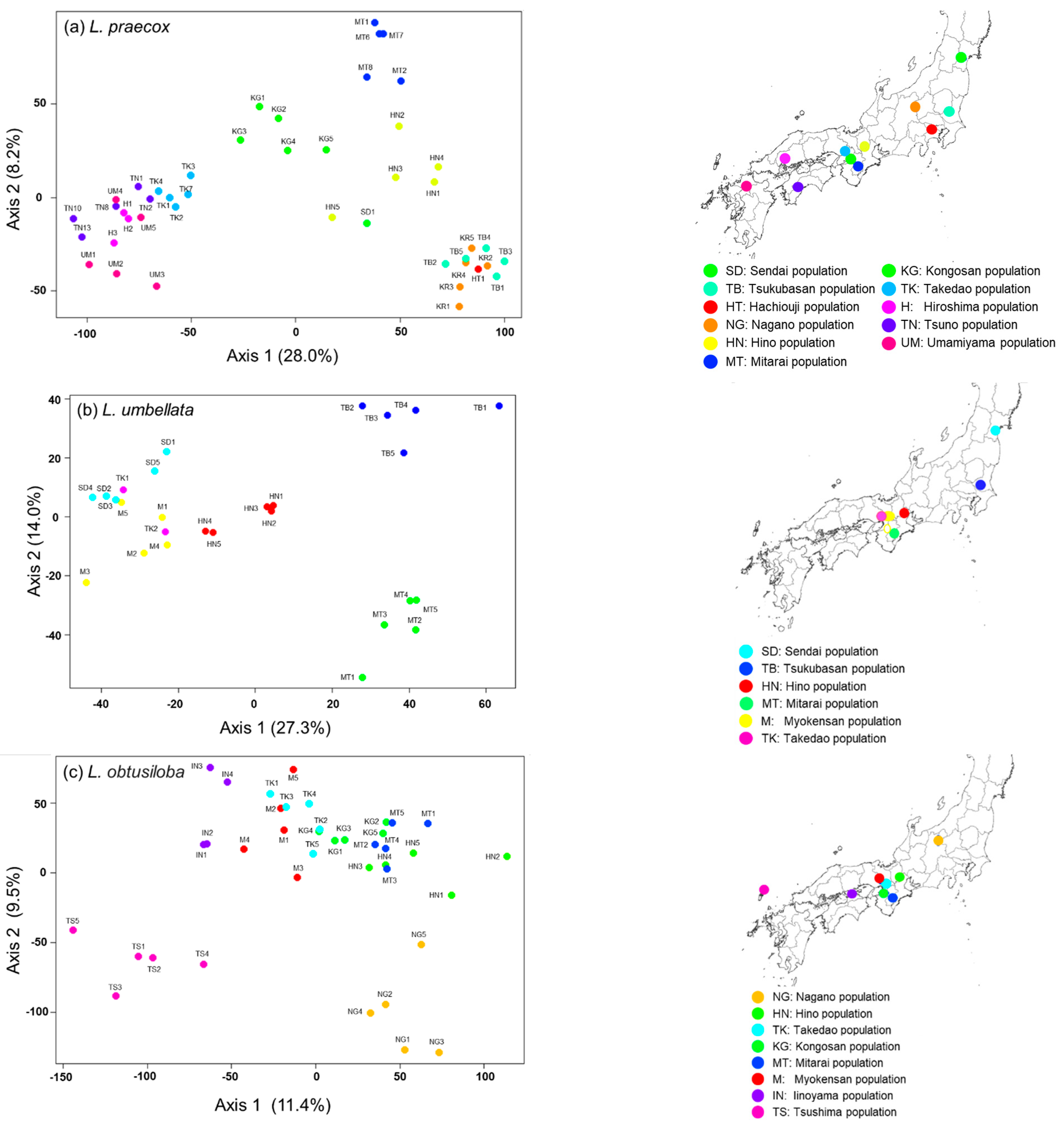
3. Results
3.1. Genetic Diversity Parameters
3.2. Clone Identification and Genotype Diversity
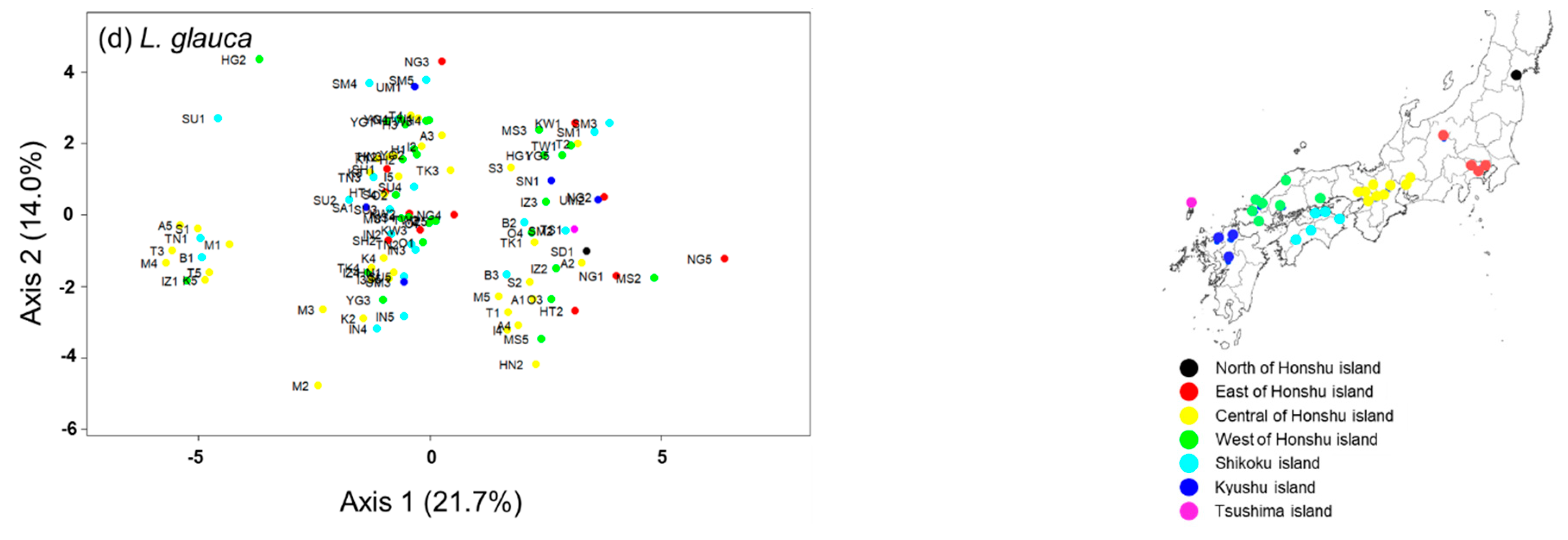
3.3. Genetic Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Accession Codes
References
- Booy, G.; Hendriks, R.J.J.; Smulders, M.J.M.; Van Groenendael, J.M.; Vosman, B. Genetic diversity and the survival of populations. Plant Biol. 2000, 2, 379–395. [Google Scholar] [CrossRef]
- Booth, R.E.; Grime, J.P. Effects of genetic impoverishment on plant community diversity. J. Ecol. 2003, 91, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Hamrick, J.L.; Godt, M.J.W. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar] [CrossRef]
- Muyle, A.; Martin, H.; Zemp, N.; Mollion, M.; Gallina, S.; Tavares, R.; Silva, A.; Bataillon, T.; Widmer, A.; Glémin, S.; et al. Dioecy is associated with high genetic diversity and adaptation rates in the plant genus Silene. Mol. Biol. Evol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nanami, S.; Kawaguchi, H.; Yamakura, T. Dioecy-induced spatial patterns of two codominant tree species, Podocarpus nagi and Neolitsea aciculata. J. Ecol. 1999, 87, 678–687. [Google Scholar] [CrossRef]
- Lo, E.Y.Y.; Stefanović, S.; Dickinson, T.A. Population genetic structure of diploid sexual and polyploid apomictic hawthorns (Crataegus; Rosaceae) in the Pacific Northwest. Mol. Ecol. 2009, 18, 1145–1160. [Google Scholar] [CrossRef]
- Dias, A.C.C.; Serra, A.C.; Sampaio, D.S.; Borba, E.L.; Bonetti, A.M.; Oliveira, P.E. Unexpectedly high genetic diversity and divergence among populations of the apomictic Neotropical tree Miconia albicans. Plant Biol. 2018, 20, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Tokumoto, Y.; Matsushita, M.; Kishimoto-Yamada, K.; Nikkeshi, A.; Isogimi, T.; Nakagawa, M. Floral visitors and reproductive success in two sequentially flowering Lindera shrubs (Lauraceae) of central Japan. J. For. Res. 2019, 24, 42–51. [Google Scholar] [CrossRef]
- Majeský, Ľ.; Vašut, R.J.; Kitner, M.; Trávníček, B. The pattern of genetic variability in apomictic clones of Taraxacum officinale indicates the alternation of asexual and sexual histories of apomicts. PLoS ONE 2012, 7, 41868. [Google Scholar] [CrossRef] [Green Version]
- Onoe, T. Palaeoenvironmental analysis based on the Pleistocene Shiobata flora in the Shiobata volcanic basin, central Japan. Rep. Geol. Surv. Jpn. 1989, 269, 1–207, (in Japanese with English Summary). [Google Scholar]
- Onoe, T. A Pleistocene flora from Ebino City, Miyazaki Prefecture, Japan. Rep. Geol. Surv. Jpn. 1971, 241, 1–46, (in Japanese with English Summary). [Google Scholar]
- Dupont, Y.L.; Kato, M. Sex ratio variation in dioecious plant species: A comparative ecological study of six species of Lindera (Lauraceae). Nord. J. Bot. 1999, 19, 529–540. [Google Scholar] [CrossRef]
- Watanabe, M. Encyclopedia of Forestry; Japan Forest Technology Association: Maruzen, Tokyo, Japan, 1961; p. 11. (In Japanese) [Google Scholar]
- Yagi, M.; Takabe, W.; Matsumi, S.; Shimode, A.; Maruyama, T.; Yonei, Y. Screening and selection of anti-glycative materials: Kuromoji (Lindera umbellata). Glycative Stress Res. 2017, 4, 317–328. [Google Scholar] [CrossRef]
- Choi, H.G.; Choi, Y.H.; Kim, J.H.; Kim, H.-H.; Kim, S.-H.; Kim, J.A.; Lee, S.M.; Na, M.; Lee, S.H. A new neolignan and lignans from the stems of Lindera obtusiloba Blume and their anti-allergic inflammatory effects. Arch. Pharm. Res. 2014, 37, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Chen, Y.; An, J.; Hou, X.; Cai, J.; Wang, J.; Zhang, Z.; Lin, S. Integrated transcriptome sequencing and dynamic analysis reveal carbon source partitioning between terpenoid and oil accumulation in developing Lindera glauca fruits. Sci. Rep. UK 2017, 5, 15017. [Google Scholar] [CrossRef] [Green Version]
- Dupont, Y.L. Evolution of apomixis as a strategy of colonization in the dioecious species Lindera glauca (Lauraceae). Popul. Ecol. 2002, 44, 293–297. [Google Scholar] [CrossRef]
- Zhu, S.S.; Comes, H.P.; Tamaki, I.; Cao, Y.N.; Sakaguchi, S.; Yap, Z.Y.; Ding, Y.Q.; Qiu, Y.X. Patterns of genotype variation and demographic history in Lindera glauca (Lauraceae), an apomict-containing dioecious forest tree. J. Biogeogr. 2020, 47, 2002–2016. [Google Scholar] [CrossRef]
- Xiong, B.; Zhang, L.; Dong, S.; Zhang, Z. Population genetic structure and variability in Lindera glauca (Lauraceae) indicates low levels of genetic diversity and skewed sex ratios in natural populations in mainland China. PeerJ 2020, 8, 8304. [Google Scholar] [CrossRef] [Green Version]
- Suyama, Y.; Matsuki, Y. MIG-seq: An effective PCR-based method for genome-wide single-nucleotide polymorphism genotyping using the next-generation sequencing platform. Sci. Rep. UK 2015, 5, 16963. [Google Scholar] [CrossRef] [Green Version]
- Catchen, J.M.; Amores, A.; Hohenlohe, P.; Cresko, W.; Postlethwait, J.H. Stacks: Building and genotyping loci de novo from short-read sequences. G3-Genes Genom. Genet. 2011, 1, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Smouse, P.E.; Peakall, R. Spatial autocorrelation analysis of individual multiallele and multilocus genetic structure. Heredity 1999, 82, 561–573. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E. pegas: An R package for population genetics with an integrated–modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.F.; Guillaume, B.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: R package version. Community Ecol. Package 2019, 2, 1–259. [Google Scholar]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Richards, Z.T.; Yasuda, N.; Kikuchi, T.; Foster, T.; Mitsuyuki, C.; Stat, M.; Suyama, Y.; Wilson, N.G. Integrated evidence reveals a new species in the ancient blue coral genus Heliopora (Octocorallia). Sci. Rep. Orts. 2018, 8, 15875. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, I.; Yoichi, W.; Matsuki, Y.; Suyama, Y.; Mizuno, M. Inconsistency between morphological traits and ancestry of individuals in the hybrid zone between two Rhododendron japonoheptamerum varieties revealed by a genotyping-by-sequencing approach. Tree Genet. Genomes 2017, 13, 1–10. [Google Scholar] [CrossRef]
- Prasetyo, E.; Widiyatno; Indrioko, S.; Na’iem, M.; Matsui, T.; Matsuo, A.; Suyama, Y.; Tsumura, Y. Genetic diversity and the origin of commercial plantation of Indonesian teak on Java Island. Tree Genet. Genomes 2020, 16, 1–14. [Google Scholar] [CrossRef]
- Ye, J.W.; Bai, W.N.; Bao, L.; Wang, T.M.; Wang, H.-F.; Ge, J.P. Sharp genetic discontinuity in the aridity-sensitive Lindera obtusiloba (Lauraceae): Solid evidence supporting the Tertiary floral subdivision in East Asia. J. Biogeogr. 2017, 44, 2082–2095. [Google Scholar] [CrossRef]
- Xiao, J.-H.; Ding, X.; Li, L.; Ma, H.; Ci, X.-Q.; van der Merwe, M.; Conran, J.G.; Li, J. Miocene diversification of a golden-thread nanmu tree species (Phoebe zhennan, Lauraceae) around the Sichuan Basin shaped by the East Asian monsoon. Ecol. Evol. 2020, 10, 10543–10557. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Zhao, W.; Maier, P.A.; Hu, X.; Jin, Y.; Zhou, S.; Jiao, S.; El-Kassaby, Y.A.; Wang, T.; Wang, X.; et al. Landscape genomics predicts climate change-related genetic offset for the widespread Platycladus orientalis (Cupressaceae). Evol. Appl. 2020, 13, 665–676. [Google Scholar] [CrossRef] [Green Version]
- González-Martínez, S.C.; Krutovsky, K.V.; Neale, D.B. Forest-tree population genomics and adaptive evolution. New Phytol. 2006, 170, 227–238. [Google Scholar] [CrossRef]
- Belletti, P.; Monteleone, I.; Ferrazzini, D. Genetic variability at allozyme markers in sycamore (Acer pseudoplatanus) populations from northwestern Italy. Can. J. For. Res. 2007, 37, 395–403. [Google Scholar] [CrossRef]
- Stoeckel, S.; Grange, J.; Fernández-Manjarres, J.F.; Bilger, I.; Frascaria-Lacoste, N.; Mariette, S. Heterozygote excess in a self-incompatible and partially clonal forest tree species—Prunus avium L. Mol. Ecol. 2006, 15, 2109–2118. [Google Scholar] [CrossRef]
- Stojnić, S.; Avramidou, E.; Fussi, B.; Westergren, M.; Orlović, S.; Matović, B.; Trudić, B.; Kraigher, H.; Aravanopoulos, F.; Konnert, M. Assessment of genetic diversity and population genetic structure of Norway spruce (Picea abies (L.) Karsten) at Its Southern lineage in Europe. Implications for conservation of forest genetic resources. Forests 2019, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Al-Qthanin, R.N.; Alharbi, S.A. Spatial structure and genetic variation of a mangrove species (Avicennia marina (Forssk.) Vierh) in the Farasan Archipelago. Forests 2020, 11, 1287. [Google Scholar] [CrossRef]
- Cai, M.; Wen, Y.; Uchiyama, K.; Onuma, Y.; Tsumura, Y. Population genetic diversity and structure of ancient tree populations of Cryptomeria japonica var. sinensis based on RAD-seq data. Forests 2020, 11, 1192. [Google Scholar] [CrossRef]
- Van Baarlen, P.; van Dijk, P.J.; Hoekstra, R.F.; de Jong, J.H. Meiotic recombination in sexual diploid and apomictic triploid dandelions (Taraxacum officinale). Stadler Gen. 2000, 43, 827–835. [Google Scholar]
- Bricker, E.; Calladine, A.; Virnstein, R.; Waycott, M. Mega clonality in an aquatic plant—a potential survival strategy in a changing environment. Front. Plant Sci. 2018, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, T.; Klahr, A.; Peterson, A.; Levichev, I.G.; Schnittler, M. No sex at all? Extremely low genetic diversity in Gagea spathacea (Liliaceae) across Europe. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 372–378. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Rasmussen, K.K.; Kollmann, J. Low genetic diversity in small peripheral populations of a rare European tree (Sorbus torminalis) dominated by clonal reproduction. Conserv. Genet. 2008, 9, 1533–1539. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Location | N | Na | Ne | He | Ho | FIS |
|---|---|---|---|---|---|---|
| Sendai, Miyagi (SD) | 1 | 0.687 | 0.687 | 0.004 | 0.008 | - |
| Hachioji, Tokyo (HT) | 2 | 0.796 | 0.792 | 0.012 | 0.021 | −0.760 |
| Sagamihara, Kanagawa (SH) | 2 | 0.936 | 0.927 | 0.012 | 0.019 | −0.538 |
| Kawasaki, Kanagawa (KW) | 3 | 0.909 | 0.896 | 0.01 | 0.014 | −0.401 |
| Nagano, Nagano (NG) | 5 | 1.023 | 1.005 | 0.014 | 0.021 | −0.504 |
| Kasugai, Aichi (I) | 5 | 1.023 | 1.009 | 0.012 | 0.018 | −0.461 |
| Aioiyama, Aichi (A) | 5 | 1.034 | 1.021 | 0.014 | 0.02 | −0.381 |
| Hino, Shiga (HN) | 3 | 0.002 | 0.003 | 0.01 | 0.018 | −0.749 |
| Takaragaike, Kyoto (T) | 5 | 1.023 | 1.017 | 0.013 | 0.018 | −0.328 |
| Kasugayama, Nara (K) | 5 | 1.019 | 1.008 | 0.011 | 0.018 | −0.598 |
| Myokensan, Osaka (M) | 5 | 1.038 | 1.029 | 0.018 | 0.027 | −0.499 |
| Shigisan, Osaka (S) | 5 | 1.026 | 1.020 | 0.011 | 0.016 | −0.434 |
| Takedao, Hyogo (TK) | 4 | 0.996 | 0.981 | 0.014 | 0.02 | −0.496 |
| Okayamashi, Okayama (O) | 5 | 1.000 | 0.995 | 0.008 | 0.012 | −0.578 |
| Hiroshimashi, Hiroshima (H) | 4 | 0.909 | 0.900 | 0.01 | 0.016 | −0.665 |
| Masuda, Shimane (MS) | 5 | 1.038 | 1.022 | 0.016 | 0.019 | −0.222 |
| Tsuwano, Shimane (TW) | 2 | 0.977 | 0.972 | 0.006 | 0.009 | −0.527 |
| Izumo, Shimane (IZ) | 5 | 1.038 | 1.023 | 0.013 | 0.017 | −0.263 |
| Yamaguchishi, Yamaguchi (YG) | 5 | 1.023 | 1.012 | 0.007 | 0.01 | −0.360 |
| Hagi, Yamaguchi (HG) | 2 | 0.962 | 0.961 | 0.003 | 0.002 | 0.429 |
| Bizan, Tokushima (B) | 3 | 1.019 | 1.015 | 0.012 | 0.019 | −0.565 |
| Iinoyama, Kagawa (IN) | 5 | 0.985 | 0.961 | 0.014 | 0.019 | −0.391 |
| Shiunzan, Kagawa (SU) | 5 | 1.049 | 1.021 | 0.018 | 0.024 | −0.306 |
| Tsuno, Kochi (TN) | 3 | 0.887 | 0.881 | 0.009 | 0.015 | −0.618 |
| Sameura, Kochi (SM) | 5 | 1.038 | 1.018 | 0.011 | 0.016 | −0.377 |
| Tsushima, Nagasaki (TS) | 1 | 0.596 | 0.596 | 0.008 | 0.015 | - |
| Umamiyama, Fukuoka (UM) | 3 | 0.898 | 0.886 | 0.011 | 0.016 | −0.435 |
| Sasga, Saga (SA) | 1 | 0.811 | 0.811 | 0.008 | 0.015 | - |
| Sannotake, Kumamoto (SN) | 1 | 0.457 | 0.457 | 0 | 0 | - |
| Regional mean | 0.903 | 0.894 | 0.011 | 0.016 | −0.441 |
| Sampling Location | N | Na | Ne | He | Ho | FIS |
|---|---|---|---|---|---|---|
| Sendai, Miyagi (SD) | 1 | 0.402 | 0.402 | 0.008 | 0.016 | - |
| Tsukubasan, Ibaraki (TB) | 5 | 1.136 | 1.064 | 0.063 | 0.056 | 0.108 |
| Hachioji, Tokyo (HT) | 1 | 0.786 | 0.786 | 0.022 | 0.043 | - |
| Nagano, Nagano (NG) | 5 | 1.125 | 1.059 | 0.059 | 0.054 | 0.094 |
| Hino, Shiga (HN) | 5 | 1.146 | 1.073 | 0.070 | 0.070 | 0.010 |
| Mitarai, Nara (MT) | 5 | 1.164 | 1.083 | 0.061 | 0.057 | 0.072 |
| Kongosan, Osaka (KG) | 5 | 1.195 | 1.114 | 0.075 | 0.068 | 0.094 |
| Takedao, Hyogo (TK) | 5 | 1.160 | 1.072 | 0.062 | 0.052 | 0.151 |
| Hiroshimashi, Hiroshima (H) | 3 | 0.996 | 0.960 | 0.043 | 0.052 | −0.223 |
| Tsuno, Kochi (TN) | 5 | 1.094 | 1.031 | 0.055 | 0.047 | 0.140 |
| Umamiyama, Fukuoka (UM) | 5 | 1.070 | 1.009 | 0.054 | 0.050 | 0.071 |
| Regional mean | 1.025 | 0.968 | 0.052 | 0.051 | 0.057 |
| Sampling Location | N | Na | Ne | He | Ho | FIS |
|---|---|---|---|---|---|---|
| Sendai, Miyagi (SD) | 5 | 1.120 | 1.102 | 0.071 | 0.089 | −0.252 |
| Tsukubasan, Ibaraki (TB) | 5 | 1.234 | 1.142 | 0.096 | 0.114 | −0.183 |
| Hino, Shiga (HN) | 5 | 1.268 | 1.133 | 0.090 | 0.088 | 0.020 |
| Mitarai, Nara (MT) | 5 | 1.204 | 1.109 | 0.080 | 0.087 | −0.089 |
| Myokensan, Osaka (M) | 5 | 1.257 | 1.138 | 0.084 | 0.084 | 0.006 |
| Takedao, Hyogo (TK) | 2 | 1.102 | 1.069 | 0.056 | 0.075 | −0.356 |
| Regional mean | 1.210 | 1.115 | 0.079 | 0.089 | −0.142 |
| Sampling Location | N | Na | Ne | He | Ho | FIS |
|---|---|---|---|---|---|---|
| Nagano, Nagano (NG) | 5 | 1.268 | 1.14 | 0.097 | 0.083 | 0.149 |
| Hino, Shiga (HN) | 5 | 1.233 | 1.133 | 0.085 | 0.083 | 0.027 |
| Mitarai, Nara (MT) | 5 | 1.203 | 1.108 | 0.078 | 0.084 | −0.078 |
| Myokensan, Osaka (M) | 5 | 1.248 | 1.134 | 0.084 | 0.073 | 0.133 |
| Kongosan, Osaka (KG) | 5 | 1.264 | 1.145 | 0.093 | 0.084 | 0.095 |
| Takedao, Hyogo (TK) | 5 | 1.249 | 1.138 | 0.085 | 0.084 | 0.008 |
| Iinoyama, Kagawa (IN) | 4 | 1.177 | 1.103 | 0.072 | 0.079 | −0.087 |
| Tsushima, Nagasaki (TS) | 5 | 1.172 | 1.095 | 0.071 | 0.076 | −0.070 |
| Regional mean | 1.227 | 1.124 | 0.083 | 0.081 | 0.022 |
| Source of Variation | df | Sum of Squares | Variance Components | % | |
|---|---|---|---|---|---|
| (A) | L. praecox | ||||
| Among population | 10 | 427,635.500 | 8034.000 | 44 | |
| Within population | 34 | 352,842.900 | 10,378.000 | 56 | |
| Total | 44 | 780,478.400 | 18,412.000 | ||
| (B) | L. umbellata | ||||
| Among population | 5 | 57,977.820 | 2084.700 | 47 | |
| Within population | 21 | 48,930.350 | 2330.000 | 53 | |
| Total | 26 | 106,908.170 | 4414.700 | ||
| (C) | L. obtusiloba | ||||
| Among population | 7 | 477,950.100 | 9319.100 | 76 | |
| Within population | 31 | 709,215.400 | 2877.900 | 24 | |
| Total | 38 | 1,187,165.500 | 12,197.000 | ||
| (D) | L. glauca | ||||
| Among population | 28 | 1005.172 | 2.706 | 9 | |
| Within population | 76 | 2007.043 | 26.066 | 91 | |
| Total | 104 | 3012.215 | 28.772 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, M.; Nanami, S.; Okuno, S.; Hirota, S.K.; Matsuo, A.; Suyama, Y.; Tokumoto, H.; Yoshihara, S.; Itoh, A. Genetic Diversity and Structure of Apomictic and Sexually Reproducing Lindera Species (Lauraceae) in Japan. Forests 2021, 12, 227. https://doi.org/10.3390/f12020227
Nakamura M, Nanami S, Okuno S, Hirota SK, Matsuo A, Suyama Y, Tokumoto H, Yoshihara S, Itoh A. Genetic Diversity and Structure of Apomictic and Sexually Reproducing Lindera Species (Lauraceae) in Japan. Forests. 2021; 12(2):227. https://doi.org/10.3390/f12020227
Chicago/Turabian StyleNakamura, Mizuho, Satoshi Nanami, Seiya Okuno, Shun K. Hirota, Ayumi Matsuo, Yoshihisa Suyama, Hayato Tokumoto, Shizue Yoshihara, and Akira Itoh. 2021. "Genetic Diversity and Structure of Apomictic and Sexually Reproducing Lindera Species (Lauraceae) in Japan" Forests 12, no. 2: 227. https://doi.org/10.3390/f12020227
APA StyleNakamura, M., Nanami, S., Okuno, S., Hirota, S. K., Matsuo, A., Suyama, Y., Tokumoto, H., Yoshihara, S., & Itoh, A. (2021). Genetic Diversity and Structure of Apomictic and Sexually Reproducing Lindera Species (Lauraceae) in Japan. Forests, 12(2), 227. https://doi.org/10.3390/f12020227