
Evaluation of Early Bark Beetle Infestation Localization by Drone-Based Monoterpene Detection

 , and
, and
Abstract
:1. Introduction
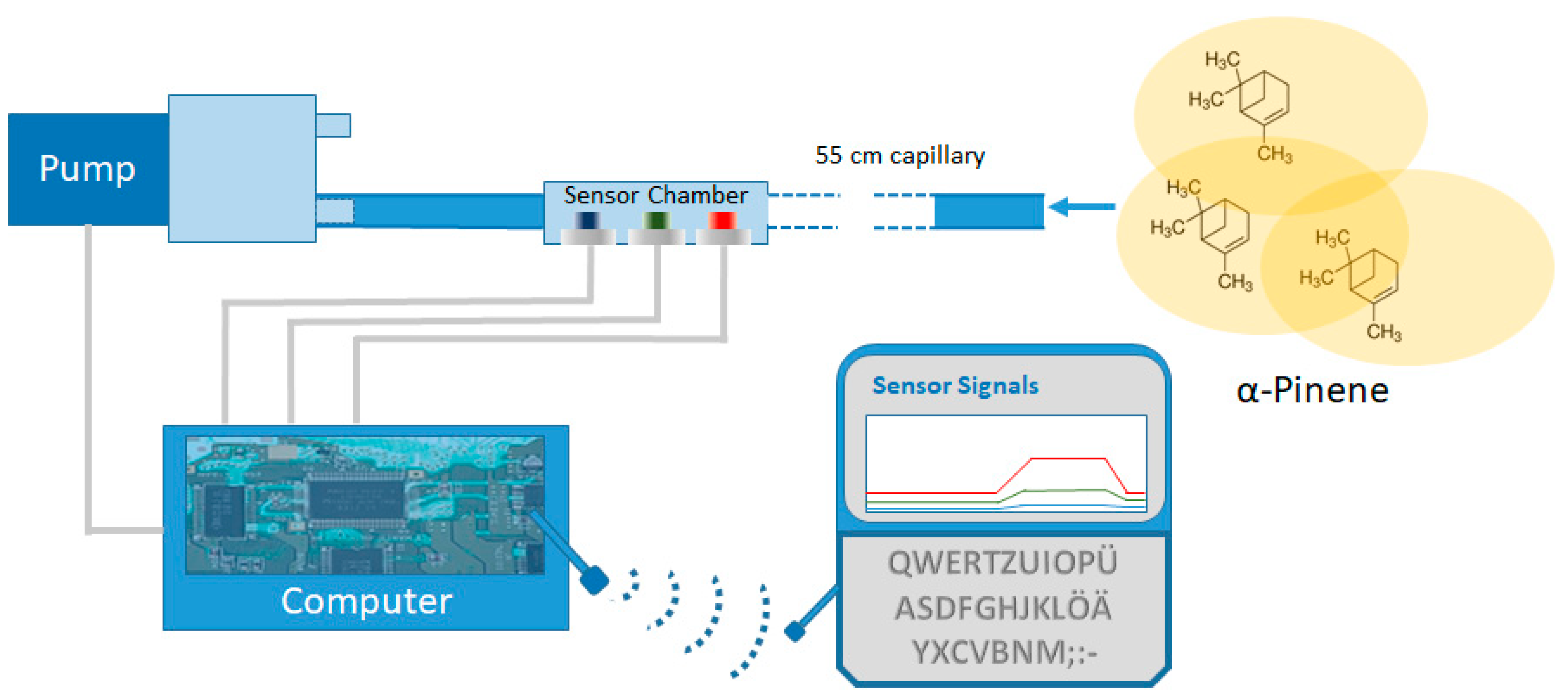
2. Materials and Methods
2.1. Sensor Calibration
Data Analysis of Sensor Calibration
2.2. Sensor Test
Data Analysis of Sensor Test
2.3. Sensor Field Test under Artificial Conditions
Data Analysis of Sensor Field Test under Artificial Conditions
2.4. Sensor Field Test above a Forest Stand
Data Analysis of Sensor Field Test above a Forest Stand
3. Results
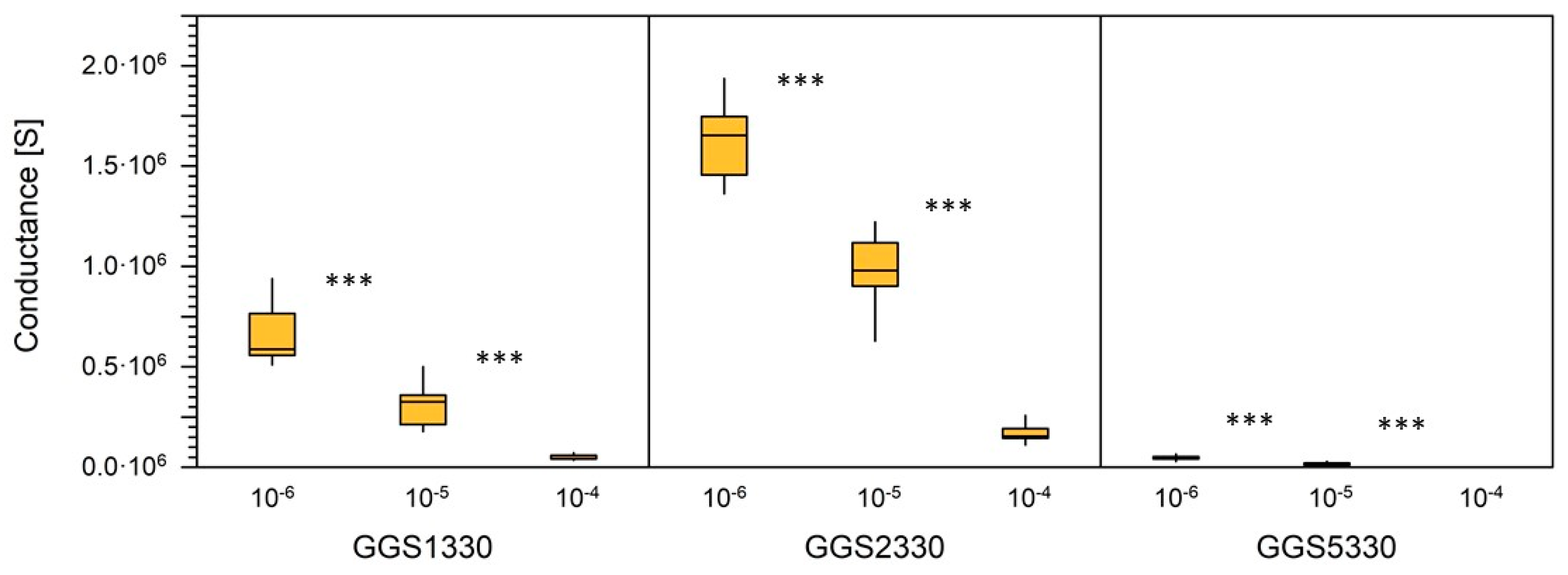
3.1. Sensor Calibration
3.2. Sensor Test under Lab Condition
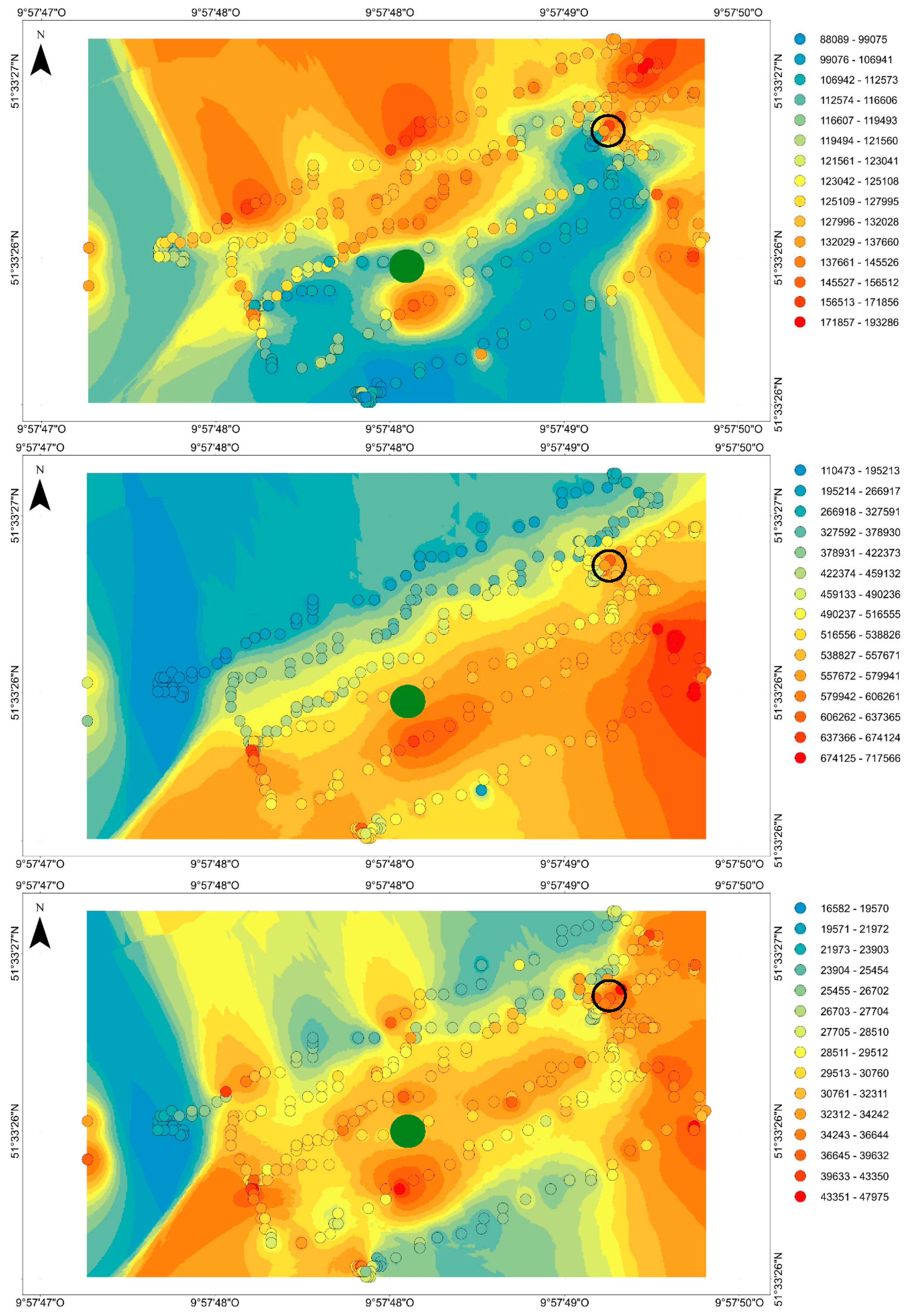
3.3. Sensor Field Test under Artificial Conditions
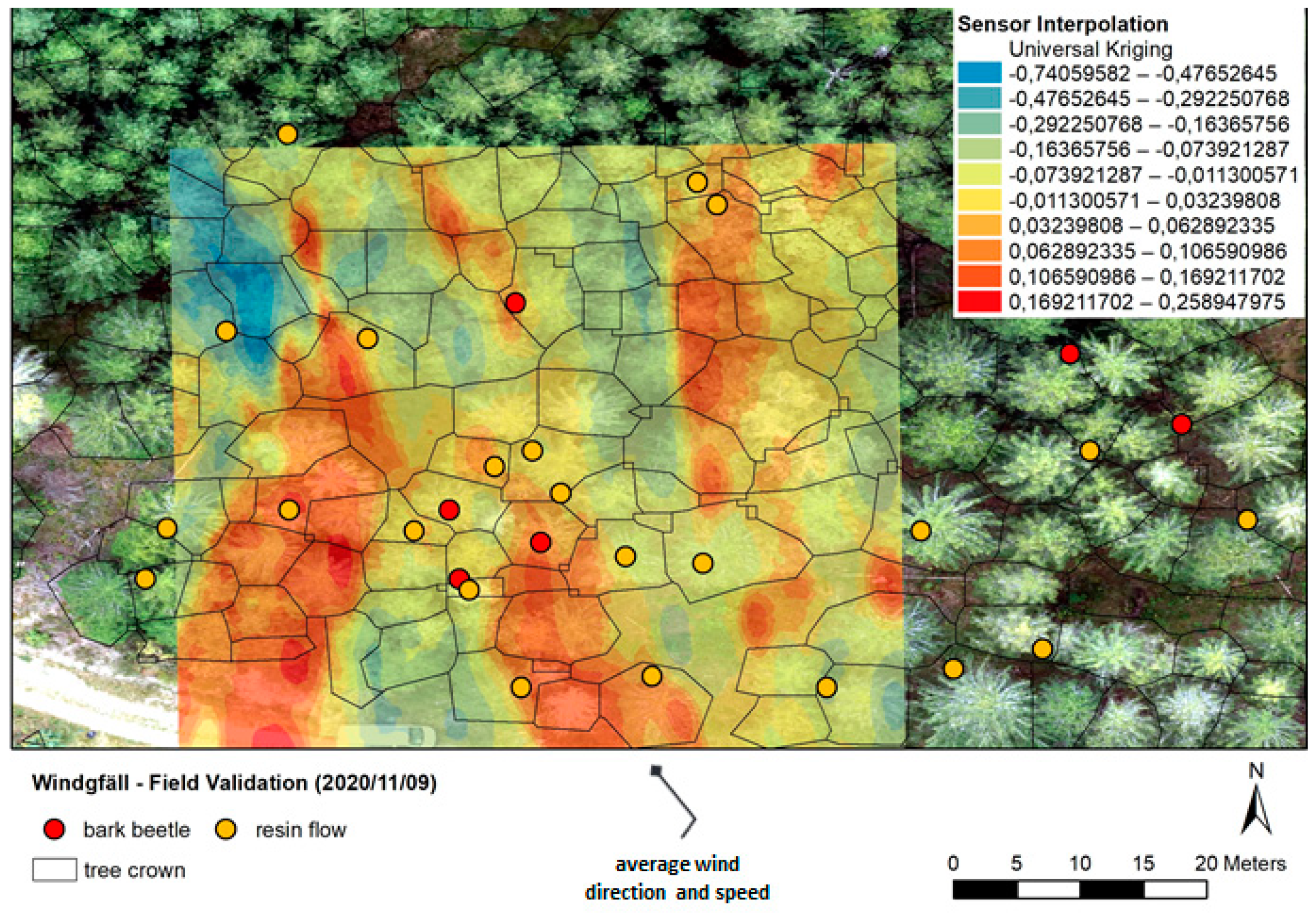
3.4. Sensor Field Test above a Forest Stand
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Grandpré, L.; Pureswaran, D.; Bouchard, M.; Kneeshaw, D. Climate-induced range shifts in boreal forest pests: Ecological, economic, and social consequences. Can. J. For. Res. 2018, 48, v–vi. [Google Scholar] [CrossRef] [Green Version]
- Rouault, G.; Candau, J.-N.; Lieutier, F.; Nageleisen, L.-M.; Martin, J.-C.; Warzée, N. Effects of drought and heat on forest insect populations in relation to the 2003 drought in Western Europe. Ann. For. Sci. 2006, 63, 613–624. [Google Scholar] [CrossRef]
- Corbett, L.J.; Withey, P.; Lantz, V.A.; Ochuodho, T.O. The economic impact of the mountain pine beetle infestation in British Columbia: Provincial estimates from a CGE analysis. Forestry 2016, 89, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.; Joyce, A.L.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Bundesministerium für Ernährung und Landwirtschaft. Waldschäden: Bundesministerium Veröffentlicht Aktuelle Zahlen. Available online: https://www.bmel.de/SharedDocs/Pressemitteilungen/DE/2020/040-waldschaeden.html;jsessionid=E322EBC9C439CDFD426657E41A3E9DC5.internet2851 (accessed on 1 July 2020).
- Bundesministerium für Ernährung und Landwirtschaft. Bundeswaldinventur. Available online: https://bwi.info/inhalt1.3.aspx?Text=3.05%20Altersklasse&prRolle=public&prInv=BWI2012&prKapitel=3.05 (accessed on 10 October 2020).
- McCollum, D.W.; Lundquist, J.E. Bark Beetle Infestation of Western US Forests: A Context for Assessing and Evaluating Impacts. J. Forest. 2019, 117, 171–177. [Google Scholar] [CrossRef]
- Wermelinger, B.; Seifert, M. Temperature-dependent reproduction of the spruce bark beetle Ips typographus, and analysis of the potential population growth. Ecol. Entomol. 1999, 24, 103–110. [Google Scholar] [CrossRef]
- Immitzer, M.; Einzmann, K.; Pinnel, N.; Seitz, R.; Atzberger, C. Vitaliltätserfassung von Fichten mittels Fernerkundung. AFZ Wald 2018, 17, 20–23. [Google Scholar]
- Anderbrant, O. Reemergence and Second Brood in the Bark Beetle Ips typographus. Holartic Ecol. 1989, 12, 494–500. [Google Scholar] [CrossRef]
- Thatcher, C.R. The Southern Pine Beetle; United State Department of Agriculture: Washington, DC, USA, 1981. [Google Scholar]
- Schwerdtfeger, F. Waldkrankheiten; Paul Parey: Hamburg/Berlin, Germany, 1970. [Google Scholar]
- Lausch, A.; Heurich, M.; Gordalla, D.; Dobner, H.-J.; Gwillym-Margianto, S.; Salbach, C. Forecasting potential bark beetle outbreaks based on spruce forest vitality using hyperspectral remote-sensing techniques at different scales. Forest. Ecol. Manag. 2013, 308, 76–89. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Ghosh, A.; Joshi, P.K.; Koch, B. Assessing the potential of hyperspectral imagery to map bark beetle-induced tree mortality. Remote Sens. Environ. 2014, 140, 533–548. [Google Scholar] [CrossRef]
- Hais, M.; Wild, J.; Berec, L.; Brůna, J.; Kennedy, R.; Braaten, J.; Brož, Z. Landsat Imagery Spectral Trajectories—Important Variables for Spatially Predicting the Risks of Bark Beetle Disturbance. Remote Sens. Environ. 2016, 8, 687. [Google Scholar] [CrossRef] [Green Version]
- Guenther, A.; Hewitt, C.N.; Erickson, D.; Fall, R.; Geron, C.; Graedel, T.; Harley, P.; Klinger, L.; Lerdau, M.; Mckay, W.A.; et al. A global model of natural volatile organic compound emissions. J. Geophys. Res. 1995, 100, 8873. [Google Scholar] [CrossRef]
- Isidorov, V.A.; Zenkevich, I.G.; Ioffe, B.V. Volatile organic compounds in the atmosphere of forests. Atmos. Environ. 1985, 19, 1–8. [Google Scholar] [CrossRef]
- Enders, G.; Dlugi, R.; Steinbrecher, R.; Clement, B.; Daiber, R.; Euk, J.; Gäb, S.; Haziza, M.; Helas, G.; Herrmann, U.; et al. Biosphere/Atmosphere interactions: Integrated research in a European coniferous forest ecosystem. Atmos. Environ. 1992, 26, 171–189. [Google Scholar] [CrossRef]
- Berg, A.R.; Heald, C.L.; Huff Hartz, K.E.; Hallar, A.G.; Meddens, A.J.H.; Hicke, J.A.; Lamarque, J.-F.; Tilmes, S. The impact of bark beetle infestations on monoterpene emissions and secondary organic aerosol formation in western North America. Atmos. Chem. Phys. 2013, 13, 3149–3161. [Google Scholar] [CrossRef] [Green Version]
- Lerdau, M.; Dilts, S.B.; Westberg, H.; Lamb, B.K.; Allwine, E.J. Monoterpene emission from ponderosa pine. J. Geophys. Res. 1994, 99, 16609. [Google Scholar] [CrossRef]
- Yokouchi, Y.; Ambe, Y. Factors Affecting the Emission of Monoterpenes from Red Pine (Pinus densiflora). Plant. Physiol. 1984, 75, 1009–1012. [Google Scholar] [CrossRef] [Green Version]
- Blanch, J.-S.; Peñuelas, J.; Llusià, J. Sensitivity of terpene emissions to drought and fertilization in terpene-storing Pinus halepensis and non-storing Quercus ilex. Physiol. Plant. 2007, 131, 211–225. [Google Scholar] [CrossRef]
- Holopainen, J.K. Multiple functions of inducible plant volatiles. Trends Plant. Sci. 2004, 9, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Ormeño, E.; Mévy, J.P.; Vila, B.; Bousquet-Mélou, A.; Greff, S.; Bonin, G.; Fernandez, C. Water deficit stress induces different monoterpene and sesquiterpene emission changes in Mediterranean species. Relationship between terpene emissions and plant water potential. Chemosphere 2007, 67, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Page, W.G.; Jenkins, M.J.; Runyon, J.B. Mountain pine beetle attack alters the chemistry and flammability of lodgepole pine foliage. Can. J. For. Res. 2012, 42, 1631–1647. [Google Scholar] [CrossRef] [Green Version]
- Page, W.G.; Jenkins, M.J.; Runyon, J.B. Spruce Beetle-Induced Changes to Engelmann Spruce Foliage Flammability. For. Sci. 2014, 60, 691–702. [Google Scholar] [CrossRef] [Green Version]
- Giunta, A.D.; Runyon, J.B.; Jenkins, M.J.; Teich, M. Volatile and Within-Needle Terpene Changes to Douglas-fir Trees Associated With Douglas-fir Beetle (Coleoptera: Curculionidae) Attack. Environ. Entomol. 2016, 45, 920–929. [Google Scholar] [CrossRef]
- Amin, H.; Atkins, P.T.; Russo, R.S.; Brown, A.W.; Sive, B.; Hallar, A.G.; Huff Hartz, K.E. Effect of bark beetle infestation on secondary organic aerosol precursor emissions. Environ. Sci. Technol. 2012, 46, 5696–5703. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, E.; Gijzen, M.; Croteau, R. Defense Mechanisms of Conifers Differences in Constitutive and Wound-Induced Monoterpene Biosynthesis Among Species. Plant. Physiol. 1991, 96, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczkowski, S.; Paczkowska, M.; Dippel, S.; Schulze, N.; Schütz, S.; Sauerwald, T.; Weiß, A.; Bauer, M.; Gottschald, J.; Kohl, C.-D. The olfaction of a fire beetle leads to new concepts for early fire warning systems. Sens. Actuat. B Chem. 2013, 183, 273–282. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Figueroa-Teran, R.; Aw, M.; Song, M.; Gorzalski, A.; Abbott, N.L.; Chang, E.; Tittiger, C. Pheromone production in bark beetles. Insect Biochem. Mol. Biol. 2010, 40, 699–712. [Google Scholar] [CrossRef]
- Hedgren, P.O. The bark beetle Pityogenes chalcographus (L.) (Scolytidae) in living trees: Reproductive success, tree mortality and interaction with Ips typographus. J. Appl. Entomol. 2004, 128, 161–166. [Google Scholar] [CrossRef]
- Byers, J.A.; Wood, D.L. Interspecific effects of pheromones on the attraction of the bark beetles, Dendroctonus brevicomis and Ips paraconfusus in the laboratory. J. Chem. Ecol. 1981, 7, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Tittiger, C.; Blomquist, G.J. Pheromone biosynthesis in bark beetles. Curr. Opin. Insect Sci. 2017, 24, 68–74. [Google Scholar] [CrossRef]
- Byers, J.A. Chemical ecology of bark beetles. Experientia 1989, 45, 271–283. [Google Scholar] [CrossRef]
- Gitau, C.W.; Bashford, R.; Carnegie, A.J.; Gurr, G.M. A review of semiochemicals associated with bark beetle (Coleoptera: Curculionidae: Scolytinae) pests of coniferous trees: A focus on beetle interactions with other pests and their associates. For. Ecol. Manag. 2013, 297, 1–14. [Google Scholar] [CrossRef]
- Progar, R.A.; Gillette, N.; Fettig, C.J.; Hrinkevich, K. Applied Chemical Ecology of the Mountain Pine Beetle. For. Sci. 2014, 60, 414–433. [Google Scholar] [CrossRef]
- Erbilgin, N.; Powell, J.S.; Raffa, K.F. Effect of varying monoterpene concentrations on the response of Ips pini (Coleoptera: Scolytidae) to its aggregation pheromone: Implications for pest management and ecology of bark beetles. Agric. For. Ent. 2003, 5, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.N.; Grosse-Wilde, E.; Keeling, C.I.; Bengtsson, J.M.; Yuen, M.M.S.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.S.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (Coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 198. [Google Scholar] [CrossRef] [Green Version]
- Yuvaraj, J.K.; Roberts, R.E.; Sonntag, Y.; Hou, X.-Q.; Grosse-Wilde, E.; Machara, A.; Zhang, D.-D.; Hansson, B.S.; Johanson, U.; Löfstedt, C.; et al. Putative ligand binding sites of two functionally characterized bark beetle odorant receptors. BMC Biol. 2021, 19, 16. [Google Scholar] [CrossRef]
- Pelosi, P. Odorant-binding proteins. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 199–228. [Google Scholar] [CrossRef]
- Kohl, D.; Heinert, L.; Bock, J.; Hofmann, T.; Schieberle, P. Systematic studies on responses of metal-oxide sensor surfaces to straight chain alkanes, alcohols, aldehydes, ketones, acids and esters using the SOMMSA approach. Sens. Actuat. B Chem. 2000, 70, 43–50. [Google Scholar] [CrossRef]
- Krüll, W.; Tobera, R.; Willms, I.; Essen, H.; von Wahl, N. Early Forest Fire Detection and Verification using Optical Smoke, Gas and Microwave Sensors. Procedia Eng. 2012, 45, 584–594. [Google Scholar] [CrossRef]
- Neuenschwander, U.; Guignard, F.; Hermans, I. Mechanism of the Aerobic Oxidation of α-Pinene. ChemSusChem 2010, 3, 75–84. [Google Scholar] [CrossRef]
- Paczkowski, S.; Pelz, S.; Jaeger, D. Semi-conductor metal oxide gas sensors for online monitoring of oak wood VOC emissions during drying. Dry. Technol. 2019, 37, 1081–1086. [Google Scholar] [CrossRef]
- Schultealbert, C.; Baur, T.; Schütze, A.; Böttcher, S.; Sauerwald, T. A novel approach towards calibrated measurement of trace gases using metal oxide semiconductor sensors. Sens. Actuat. B Chem. 2017, 239, 390–396. [Google Scholar] [CrossRef]
- Schüler, M.; Helwig, N.; Ventura, G.; Schütze, A.; Sauerwald, T. IEEE Sensors, Proceedings of the 12th IEEE Sensors Conference, Baltimore, Maryland, USA, 3–6 November 2013; IEEE: Piscataway, NJ, USA, 2013; ISBN 9781467346405. [Google Scholar]
- Leidinger, M.; Sauerwald, T.; Conrad, T.; Reimringer, W.; Ventura, G.; Schütze, A. Selective Detection of Hazardous Indoor VOCs Using Metal Oxide Gas Sensors. Proc. Engin. 2014, 87, 1449–1452. [Google Scholar] [CrossRef] [Green Version]
- Bohbot, J.D.; Vernick, S. The Emergence of Insect Odorant Receptor-Based Biosensors. Biosensors 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.-H.; Yang, W.-X.; Wang, Y.; Li, F.; Zeng, M. Adapting an ant colony metaphor for multi-robot chemical plume tracing. Sensors 2012, 12, 4737–4763. [Google Scholar] [CrossRef] [PubMed]
- de Croon, G.; O’Connor, L.M.; Nicol, C.; Izzo, D. Evolutionary robotics approach to odor source localization. Neurocomputing 2013, 121, 481–497. [Google Scholar] [CrossRef]
- Monroy, J.; Hernandez-Bennets, V.; Fan, H.; Lilienthal, A.; Gonzalez-Jimenez, J. GADEN: A 3D Gas Dispersion Simulator for Mobile Robot Olfaction in Realistic Environments. Sensors 2017, 17, 1479. [Google Scholar] [CrossRef] [Green Version]
- Ishida, H.; Wada, Y.; Matsukura, H. Chemical Sensing in Robotic Applications: A Review. IEEE Sens. J. 2012, 12, 3163–3173. [Google Scholar] [CrossRef]
- Balkovsky, E.; Shraiman, B.I. Olfactory search at high Reynolds number. Proc. Natl. Acad. Sci. USA 2002, 99, 12589–12593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turski, M.; Beker, C.; Kazmierczak, K.; Najgrakowski, T. Allometric equations for estimating the mass and volume of fresh assimilational apparatus of standing scots pine (Pinus sylvestris L.) trees. For. Ecol. Manag. 2008, 255, 2678–2687. [Google Scholar] [CrossRef]
- Li, Y.; Ma, H.; Wan, Y.; Li, T.; Liu, X.; Sun, Z.; Li, Z. Volatile Organic Compounds Emissions from Luculia pinceana Flower and Its Changes at Different Stages of Flower Development. Molecules 2016, 21, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, U.; Rottenberger, S.; Biesenthal, T.; Wolf, A.; Schebeske, G.; Ciccioli, P.; Kesselmeier, J. Strong correlation between isoprene emission and gross photosynthetic capacity during leaf phenology of the tropical tree species Hymenaea courbaril with fundamental changes in volatile organic compounds emission composition during early leaf development. Plant. Cell Environ. 2004, 27, 1469–1485. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species (Specification) | Compound (Unit) | Emission (Not Infested) | Emission (Infested) | Infestation Stage | Emission Ratio a | Source |
| Pinus contorta (canopy) | α-pinene (ng × L−1) | 1.1 ± 0.56 | 1.5 ± 0.6 | unknown | 1.4 | [28] |
| 2.6 ± 0.9 | 2.7 ± 1.1 | 1.0 | ||||
| Pseudotsuga menziesii (lower branches) | α-pinene (ng × h−1 × gFM−1) | 322 ± 166 | 813 ± 482 | Green | 2.5 | [27] |
| Pseudotsuga menziesii (lower branches) | Total terpenes (ng × h−1 × gFM−1) | 870 ± 417 | 4472 ± 2759 | Green | 5.1 | [27] |
| 1515 ± 737 | 1472 ± 834 | 1.0 | ||||
| 1695 ± 797 | 5881 ± 3685 | 3.4 | ||||
| 571 ± 226 | 2480 ± 1094 | 4.3 | ||||
| 284 ± 59 | 462 ± 189 | 1.6 | ||||
| 557 ± 251 | 229 ± 68 | 0.4 | ||||
| 842 ± 268 | 724 ± 243 | 0.9 | ||||
| 948 ± 684 | 2124 ± 1782 | 2.2 | ||||
| Pseudotsuga menziesii (lower branches) | α-pinene (ng × h−1 × gFM−1) | 77 | 190 | Yellow b | 2.5 | [26] |
| 100 | 190 | 1.9 | ||||
| 60 | 134 | 2.2 | ||||
| 70 | 184 | 2.6 | ||||
| Tree Species | Compound | Emission (No Draught) | Emission (Draught) | Draught Intensity | Emission Ratio a | Source |
| Pinus halepensis (seedlings) | α-pinene (µg × h−1 × gDM−1) | 23.9 ± 5.8 | 17.9 ± 1.3 | 2 weeks | 0.7 | [22] |
| 5.6 ± 0.7 | 16.4 ± 5.3 | 4 weeks | 2.9 | [22] | ||
| 8.8 ± 3.2 | 4.0 ± 1.3 | 6 weeks | 0.5 | [22] | ||
| 30.7 ± 4.5 | 20.6 ± 4.8 | 8 weeks | 0.7 | [22] | ||
| 10.9 ± 4.5 | 21.2 ± 5.8 | 10 weeks | 1.9 | [22] | ||
| 34.2 ± 7.5 | 21.4 ± 4.8 | 12 weeks | 0.6 | [22] | ||
| Pinus halepensis (seedlings) | α-pinene (%total VOC) | 30.8 ± 10.6 | 60.0 ±14.3 | 1 week | 1.9 | [24] |
| 10−6 | 10−5 | 10−4 | |
|---|---|---|---|
| GGS1330, N = 13 | |||
| Mean (S) | 669,022 | 308,851 | 51,870 |
| Standard deviation (S) | 147,007 | 114,605 | 11,260 |
| Variation coefficient (%) | 22 | 37 | 22 |
| R2 of mean | y = 51,595x2 – 514,955x + 1,132,382 R2 = 1.00 | ||
| R2 of all values (linear) | y = 1,171,683x – 302,126 R2 = 0.83 | ||
| GGS2330, N = 13 | |||
| Mean (S) | 1,622,037 | 990,640 | 171,820 |
| Standard deviation (S) | 193,071 | 15,4917 | 40,674 |
| Variation coefficient (%) | 12 | 16 | 24 |
| Linear R2 of mean | y = −93,711x2 − 350,263x + 2,066,011 R2 = 1.00 | ||
| R2 of all values (linear) | y = 2,748,138x − 736,822 R2 = 0.94 | ||
| GGS5330, N = 13 | |||
| Mean (S) | 47,119 | 16,323 | 1551 |
| Standard deviation (S) | 10,525 | 6814 | 538 |
| Variation coefficient (%) | 22 | 42 | 35 |
| Linear R2 of mean | y = 8011x2 − 54,830x + 93,938 R2 = 1.00 | ||
| R2 of all values (linear) | y = −87,914x + 21,782 R2 = 0.83 | ||
| GGS1330 (S), N = 5 | GGS2330 (S), N = 5 | GGS5330 (S), N = 5 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| °C | Amount α-pinene (mL) | 1.5 m; 1 m/s | 1.5 m; 2 m/s | 3 m; 1 m/s | 3 m; 2 m/s | 1.5m; 1 m/s | 1.5m; 2 m/s | 3 m; 1 m/s | 3 m; 2 m/s | 1.5 m; 1 m/s | 1.5 m; 2 m/s | 3 m; 1 m/s | 3 m; 2 m/s |
| 15 | 0.1 | 78.383 | 89.711 | 79.732 | 91.260 | 73.774 | 71.256 | 77.092 | 86.777 | 9.107 | 10.700 | 9.567 | 13.508 |
| 3.648 | 1.663 | 1.669 | 1.449 | 4.160 | 2.965 | 1.121 | 3.056 | 711 | 345 | 321 | 689 | ||
| 5% | 2% | 2% | 2% | 6% | 4% | 1% | 4% | 8% | 3% | 3% | 5% | ||
| 0.5 | 100.692 | 96.687 | 92.099 | 89.899 | 85.104 | 61.596 | 80.888 | 77.253 | 9.535 | 6.773 | 8.116 | 8.857 | |
| 1.742 | 2.850 | 1.954 | 2.556 | 3.467 | 3.668 | 2.518 | 2.263 | 759 | 249 | 374 | 520 | ||
| 2% | 3% | 2% | 3% | 4% | 6% | 3% | 3% | 8% | 4% | 5% | 6% | ||
| 20 | 0.1 | 81.612 | 80.852 | 74.108 | 88.336 | 82.872 | 76.040 | 77.313 | 93.394 | 14.005 | 11.515 | 10.917 | 16.927 |
| 2.430 | 2.567 | 2.184 | 1.241 | 3.098 | 6.387 | 2.465 | 2.877 | 901 | 482 | 365 | 538 | ||
| 3% | 3% | 3% | 1% | 4% | 8% | 3% | 3% | 6% | 4% | 3% | 3% | ||
| 0.5 | 64.598 | 73.860 | 77.323 | 74.619 | 69.516 | 59.824 | 78.921 | 75.072 | 7.755 | 8.280 | 9.721 | 10.165 | |
| 2.165 | 2.577 | 3.070 | 2.350 | 3.243 | 1.711 | 2.986 | 1.373 | 323 | 577 | 278 | 275 | ||
| 3% | 3% | 4% | 3% | 5% | 3% | 4% | 2% | 4% | 7% | 3% | 3% | ||
| 25 | 0.1 | 57.741 | 70.578 | 77.092 | 73.391 | 69.281 | 75.375 | 92.740 | 87.419 | 10.042 | 13.602 | 18.530 | 16.703 |
| 787 | 1.754 | 2.472 | 1.328 | 3.098 | 3.199 | 6.931 | 3.379 | 1.030 | 499 | 1.654 | 398 | ||
| 1% | 2% | 3% | 2% | 4% | 4% | 7% | 4% | 10% | 4% | 9% | 2% | ||
| 0.5 | 54.504 | 63.156 | 68.440 | 66.394 | 63.844 | 62.421 | 82.840 | 79.421 | 7.813 | 8.494 | 12.276 | 11.734 | |
| 4.158 | 3.247 | 2.804 | 2.112 | 4.094 | 3.408 | 1.203 | 987 | 588 | 514 | 919 | 737 | ||
| 8% | 5% | 4% | 3% | 6% | 5% | 1% | 1% | 8% | 6% | 7% | 6% | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paczkowski, S.; Datta, P.; Irion, H.; Paczkowska, M.; Habert, T.; Pelz, S.; Jaeger, D. Evaluation of Early Bark Beetle Infestation Localization by Drone-Based Monoterpene Detection. Forests 2021, 12, 228. https://doi.org/10.3390/f12020228
Paczkowski S, Datta P, Irion H, Paczkowska M, Habert T, Pelz S, Jaeger D. Evaluation of Early Bark Beetle Infestation Localization by Drone-Based Monoterpene Detection. Forests. 2021; 12(2):228. https://doi.org/10.3390/f12020228
Chicago/Turabian StylePaczkowski, Sebastian, Pawan Datta, Heidrun Irion, Marta Paczkowska, Thilo Habert, Stefan Pelz, and Dirk Jaeger. 2021. "Evaluation of Early Bark Beetle Infestation Localization by Drone-Based Monoterpene Detection" Forests 12, no. 2: 228. https://doi.org/10.3390/f12020228
APA StylePaczkowski, S., Datta, P., Irion, H., Paczkowska, M., Habert, T., Pelz, S., & Jaeger, D. (2021). Evaluation of Early Bark Beetle Infestation Localization by Drone-Based Monoterpene Detection. Forests, 12(2), 228. https://doi.org/10.3390/f12020228