Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Occurrence Data
2.2. Environmental Variables
2.3. Modelling Procedure
2.4. Centroid Shifts
3. Results
3.1. Model Performance
3.2. Variable Contribution
3.3. Current and Future Potential Distributions
3.4. Centroid Shifts
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leadley, P. Biodiversity Scenarios: Projections of 21st Century Change in Biodiversity, and Associated Ecosystem Services: A Technical Report for the Global Biodiversity Outlook 3; Secretariat of the Convention on Biological Diversity: Montreal, CA, Canada, 2010. [Google Scholar]
- Subba, B.; Sen, S.; Ravikanth, G.; Nobis, M.P. Direct modelling of limited migration improves projected distributions of Himalayan amphibians under climate change. Biol. Conserv. 2018, 227, 352–360. [Google Scholar] [CrossRef]
- Wiens, J.J. Climate-related local extinctions are already widespread among plant and animal species. PLoS Biol. 2016, 14, e2001104. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.B.; Jia, J.S.; Wang, C.L. A general review of the conservation status of Chinese orchids. Biodiv. Sci. 2003, 11, 70–77. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Štípková, Z.; Kindlmann, P. Orchid extinction over the last 150 years in the Czech Republic. Diversity 2021, 13, 78. [Google Scholar] [CrossRef]
- Wagensommer, R.P.; Medagli, P.; Turco, A.; Perrino, E.V. IUCN Red List evaluation of the Orchidaceae endemic to Apulia (Italy) and considerations on the application of the IUCN protocol to rare species. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 90–101. [Google Scholar] [CrossRef]
- Rankou, H. Cypripedium Japonicum. In The IUCN Red List of Threatened Species, 4th ed.; International Union for Conservation of Nature: Gland, Switzerland, 2014. [Google Scholar]
- Cribb, P.; Sandison, M.S. A preliminary assessment of the conservation status of Cypripedium species in the wild. Bot. J. Linn. Soc. 1998, 126, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Cribb, P.; Mathew, B. Plate 324. Cypripedium henryi: Orchidaceae. Curtis’s Bot. Mag. 1997, 14, 148–153. [Google Scholar] [CrossRef]
- Qian, X.; Li, Q.J.; Liu, F.; Gong, M.J.; Wang, C.X.; Tian, M. Conservation genetics of an endangered lady’s slipper orchid: Cypripedium japonicum in China. Int. J. Mol. Sci. 2014, 15, 11578–11596. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.Z.; Han, L.X.; Zhang, J.L.; Li, X.L.; Kawahara, T.; Yukawa, T.; López-Pujol, J.; Kumar, P.; Chung, M.G.; Chung, M.Y. Genetic diversity in the endangered terrestrial orchid Cypripedium japonicum in East Asia: Insights into population history and implications for conservation. Sci. Rep. 2018, 8, 6467. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.W.; Fan, C.S.; Zhu, Z.Y. National Chinese Herbal Medicine Compilation (II), 2nd ed.; People’s Medical Publishing House: Beijing, China, 1978; p. 466. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, S.; Capinha, C.; Weterings, R.; Gao, T. Using species distribution model to predict the impact of climate change on the potential distribution of Japanese whiting Sillago japonica. Ecol. Indic. 2019, 104, 333–340. [Google Scholar] [CrossRef]
- Lin, C.T.; Chiu, C.A. The relic Trochodendron aralioides Siebold & Zucc. (Trochodendraceae) in Taiwan: Ensemble distribution modeling and climate change impacts. Forests 2018, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Wu, J.; Guan, Q. The potential effects of future climate change on suitable habitat for the Taiwan partridge (Arborophila crudigularis): An ensemble-based forecasting method. Turk. J. Zool. 2017, 41, 513–521. [Google Scholar] [CrossRef]
- Xu, D.; Zhuo, Z.; Wang, R.; Ye, M.; Pu, B. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Glob. Ecol. Conserv. 2019, 19, e00691. [Google Scholar] [CrossRef]
- Uusitalo, R.; Siljander, M.; Culverwell, C.L.; Mutai, N.C.; Forbes, K.M.; Vapalahti, O.; Pellikka, P.K.E. Predictive mapping of mosquito distribution based on environmental and anthropogenic factors in Taita Hills, Kenya. Int. J. Appl. Earth Obs. Geoinf. 2019, 76, 84–92. [Google Scholar] [CrossRef]
- Mathai, J.; Niedballa, J.; Radchuk, V.; Sollmann, R.; Heckmann, I.; Brodie, J.; Struebig, M.; Hearn, A.J.; Ross, J.; Macdonald, D.W.; et al. Identifying refuges for Borneo’s elusive Hose’s civet. Glob. Ecol. Conserv. 2019, 17, e00531. [Google Scholar] [CrossRef]
- Wang, H.H.; Wonkka, C.L.; Treglia, M.L.; Grant, W.E.; Smeins, F.E.; Rogers, W.E. Species distribution modelling for conservation of an endangered endemic orchid. AoB Plants 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Suetsugu, K.; Fukushima, S. Pollination biology of the endangered orchid Cypripedium japonicum in a fragmented forest of Japan. Plant Species Biol. 2014, 29, 294–299. [Google Scholar] [CrossRef]
- Sun, H.Q.; Cheng, J.; Zhang, F.M.; Luo, Y.B.; Ge, S. Reproductive success of non-rewarding Cypripedium japonicum benefits from low spatial dispersion pattern and asynchronous flowering. Ann. Bot. 2009, 103, 1227–1237. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, C.; Tian, M.; Li, C. Variation of phenotypic traits of wild populations of Cypripedium japonicum in Zhejiang Province and its correlation with geographical-soil nutrient factors. J. Plant Resour. Environ. 2012, 21, 45–52. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Buri, A.; Cianfrani, C.; Pinto-Figueroa, E.; Yashiro, E.; Spangenberg, J.E.; Adatte, T.; Verrecchia, E.; Guisan, A.; Pradervand, J.-N. Soil factors improve predictions of plant species distribution in a mountain environment. Prog. Phys. Geogr. 2017, 41, 703–722. [Google Scholar] [CrossRef]
- Watanabe, S.; Hajima, T.; Sudo, K.; Nagashima, T.; Takemura, T.; Okajima, H.; Nozawa, T.; Kawase, H.; Abe, M.; Yokohata, T.; et al. MIROC-ESM: Model description and basic results of CMIP5-20c3m experiments. Geosci. Model Dev. Discuss. 2011, 4, 1063–1128. [Google Scholar] [CrossRef] [Green Version]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Tian, Z.; Jiang, D. Evaluation of the performance of low-to high-resolution CCSM4 over East Asia and China. Chin. J. Atmos. Sci. 2013, 37, 171–186. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- De Marco, P.J.; Nóbrega, C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar] [CrossRef]
- Mansfield, E.R.; Helms, B.P. Detecting Multicollinearity. Am. Stat. 1982, 36, 158–160. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Barbosa, A.M.; O’Hara, R.B. FuzzySim: Applying fuzzy logic to binary similarity indices in ecology. Methods Ecol. Evol. 2015, 6, 853–858. [Google Scholar] [CrossRef]
- Watling, J.I.; Brandt, L.A.; Bucklin, D.N.; Fujisaki, I.; Mazzotti, F.J.; Romañach, S.S.; Speroterra, C. Performance metrics and variance partitioning reveal sources of uncertainty in species distribution models. Ecol. Modell. 2015, 309–310, 48–59. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Zhang, J.E.; Jiang, Y.P.; Wang, R.L.; Wu, R.S. Predicting the potential distribution of Pseudomonas syringae pv. actinidiae in China using ensemble models. Plant Pathol. 2020, 69, 120–131. [Google Scholar] [CrossRef]
- Brown, J.L.; Anderson, B. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Cassini, M.H. Ranking threats using species distribution models in the IUCN Red List assessment process. Biodivers. Conserv. 2011, 20, 3689–3692. [Google Scholar] [CrossRef]
- Hamid, M.; Khuroo, A.A.; Charles, B.; Ahmad, R.; Singh, C.P.; Aravind, N.A. Impact of climate change on the distribution range and niche dynamics of Himalayan birch, a typical treeline species in Himalayas. Biodivers. Conserv. 2018, 28, 2345–2370. [Google Scholar] [CrossRef]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting future expansion of invasive species: Comparing and improving methodologies for species distribution modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef]
- Shrestha, U.B.; Sharma, K.P.; Devkota, A.; Siwakoti, M.; Shrestha, B.B. Potential impact of climate change on the distribution of six invasive alien plants in Nepal. Ecol. Indic. 2018, 95, 99–107. [Google Scholar] [CrossRef]
- Lang, K.Y.; Chen, X.Q.; Luo, Y.B.; Zhu, G.H. Flora of China; Science Press: Beijing, China, 1999; Volume 17, p. 41. [Google Scholar]
- Arslan, E.S.; Akyol, A.; Örücü, Ö.K.; Sarıkaya, A.G. Distribution of rose hip (Rosa canina L.) under current and future climate conditions. Reg. Environ. Chang. 2020, 20, 107. [Google Scholar] [CrossRef]
- Li, X. Genetic Diversity and Phylogeography of a Rare Endemic Orchid: Cypripedium japonicum Thunb in East Asia. Master’s Thesis, East China Normal University, Shanghai, China, 2014. [Google Scholar]
- Guo, Y.; Bi, S.B.; Zhang, Y.Q.; Huang, J.L. Estimation and analysis for geographic and orographic influences on precipitation distribution on the yangtze plain, middle and lower. Sci. Tech. Eng. 2013, 13, 8555–8560. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Zhou, C.; Meng, J.; Sun, J.; Zhou, T.; Tao, J. Impact of climate factors on future distributions of Paeonia ostii across China estimated by MaxEnt. Ecol. Inform. 2019, 50, 62–67. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Woodward, F.I.; Lomas, M.R.; Kelly, C.K. Global climate and the distribution of plant biomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 1465–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.M.; Heglund, P.; Morrison, M.L. Predicting Species Occurrences: Issues of Accuracy and Scale; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Duputié, A.; Zimmermann, N.E.; Chuine, I. Where are the wild things? Why we need better data on species distribution. Glob. Ecol. Biogeogr. 2014, 23, 457–467. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total. Environ. 2020, 698, 134141. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.H.; Ren, G.Y.; Shi, G.Y.; Gong, P.; Zheng, X.H.; Zhai, P.M.; Zhang, D.E.; Zhao, Z.C.; Wang, S.W.; Wang, H.J. National assessment report of climate change (I): Climate change in China and its future trend. Adv. Clim. Chang. Res. 2006, 2, 3–8. [Google Scholar]
- Robbirt, K.M.; Roberts, D.L.; Hutchings, M.J.; Davy, A.J. Potential disruption of pollination in a sexually deceptive orchid by climatic change. Curr. Biol. 2014, 24, 2845–2849. [Google Scholar] [CrossRef] [Green Version]
- Bates, B.; Kundzewicz, Z.W.; Wu, S.; Palutikof, J. Climate Change and Water; Intergovernmental Panel on Climate Change Secretariat: Geneva, Switzerland, 2008. [Google Scholar]
- Jeon, C.W.; Kim, D.R.; Gang, G.H.; Kim, B.B.; Kim, N.H.; Nam, S.Y.; Kwak, Y.S. First report of gray mold disease on endangered species Cypripedium japonicum. MycoBiol. 2020, 48, 423–426. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Smithson, A.; Macnair, M.R. Negative frequency-dependent selection by pollinators on artificial flowers without rewards. Evolution 1997, 51, 715–723. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Code | Description | Unit |
|---|---|---|---|
| Bioclimatic factors | Bio01 | Annual mean temperature | °C |
| Bio02 | Mean diurnal range (Mean of monthly (max temp–min temp)) | °C | |
| Bio03 * | Isothermality (Bio02/Bio07) (* 100) | - | |
| Bio04 | Temperature seasonality (standard deviation * 100) | °C | |
| Bio05 | Max temperature of warmest month | °C | |
| Bio06 | Min temperature of coldest month | °C | |
| Bio07 | Temperature annual range (Bio05-Bio06) | °C | |
| Bio08 * | Mean temperature of wettest quarter | °C | |
| Bio09 * | Mean temperature of driest quarter | °C | |
| Bio10 | Mean temperature of warmest quarter | °C | |
| Bio11 | Mean temperature of coldest quarter | °C | |
| Bio12 | Annual precipitation | mm | |
| Bio13 | Precipitation of wettest month | mm | |
| Bio14 | Precipitation of driest month | mm | |
| Bio15 * | Precipitation seasonality (Coefficient of Variation) | 1 | |
| Bio16 | Precipitation of wettest quarter | mm | |
| Bio17 | Precipitation of driest quarter | mm | |
| Bio18 * | Precipitation of warmest quarter | mm | |
| Bio19 * | Precipitation of coldest quarter | mm | |
| Topographic factors | Alt | Altitude | m |
| Aspect * | Aspect | % | |
| Slope * | Slope | ° | |
| Edaphic factors | T_CaCO3 * | Topsoil calcium carbonate | % weight |
| T_Clay * | Topsoil clay fraction | % wt. | |
| T_Gravel * | Topsoil gravel content | % vol. | |
| T_OC * | Topsoil organic carbon | % weight | |
| T_Sand * | Topsoil sand fraction | % wt. | |
| T_PH | Topsoil pH (H2O) | −log(H+) |
| Model | TSS | Kappa | ROC |
|---|---|---|---|
| Random forest (RF) | 0.915 | 0.847 | 0.983 |
| Generalized boosting model (GBM) | 0.906 | 0.824 | 0.981 |
| Generalized linear model (GLM) | 0.903 | 0.806 | 0.973 |
| Multiple adaptive regression splines (MARS) | 0.903 | 0.818 | 0.970 |
| Maximum entropy (Maxent) | 0.888 | 0.797 | 0.966 |
| Flexible discriminant analysis (FDA) | 0.866 | 0.784 | 0.965 |
| Artificial neural network (ANN) | 0.823 | 0.692 | 0.933 |
| Classification tree analysis (CTA) | 0.848 | 0.765 | 0.923 |
| Generalized additive model (GAM) | 0.777 | 0.744 | 0.893 |
| Surface range envelop (SRE) | 0.586 | 0.656 | 0.793 |
| Ensemble model | 0.968 | 0.906 | 0.995 |
| Code | Mean Variable Importance | Percentage |
|---|---|---|
| Bio18 | 0.7347 | 24.00% |
| Bio09 | 0.7161 | 23.39% |
| Bio15 | 0.5841 | 19.08% |
| Slope | 0.3669 | 11.98% |
| Bio03 | 0.3297 | 10.77% |
| Bio19 | 0.1579 | 5.16% |
| Bio08 | 0.0847 | 2.77% |
| T_Clay | 0.0267 | 0.87% |
| Aspect | 0.0156 | 0.51% |
| T_CaCO3 | 0.0153 | 0.50% |
| T_Gravel | 0.0144 | 0.47% |
| T_Sand | 0.0123 | 0.40% |
| T_OC | 0.0033 | 0.11% |
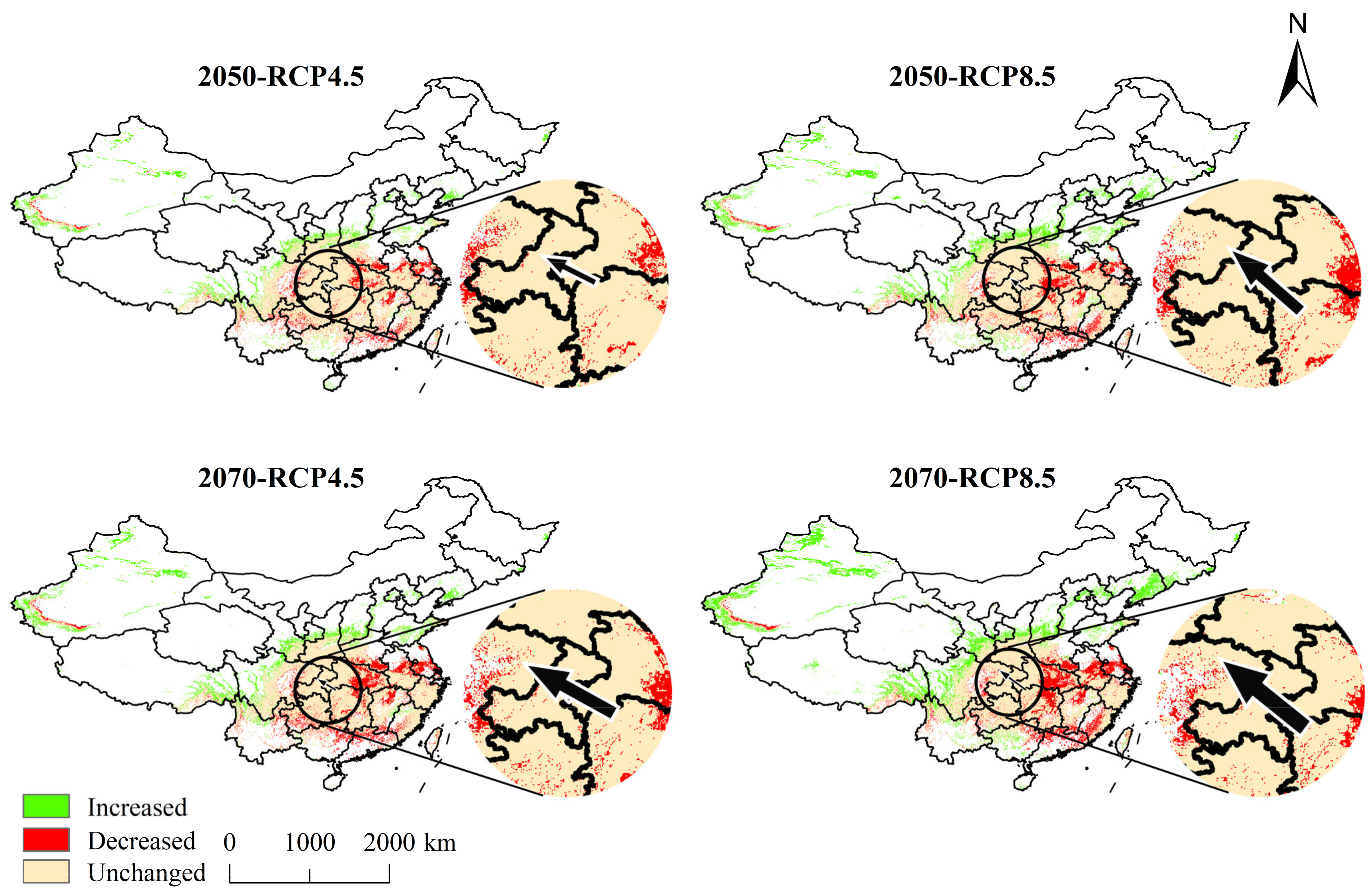
| Area (×105 km2) and Change (%) | Lowly Suitable | Moderately Suitable | Highly Suitable | Total | |||||
|---|---|---|---|---|---|---|---|---|---|
| Current | 9.66 | 2.70 | 4.53 | 16.89 | |||||
| RCP4.5 | 2050 | 10.84 | ↑12.19% | 4.02 | ↑49.32% | 2.49 | ↓44.95% | 17.36 | ↑2.79% |
| 2070 | 11.19 | ↑15.84% | 4.23 | ↑56.99% | 1.92 | ↓57.69% | 17.34 | ↑2.68% | |
| RCP8.5 | 2050 | 11.66 | ↑20.65% | 4.45 | ↑65.28% | 2.27 | ↓49.98% | 18.38 | ↑8.83% |
| 2070 | 13.63 | ↑41.03% | 4.49 | ↑66.58% | 1.26 | ↓72.24% | 19.37 | ↑14.72% | |
| Area (×105 km2) | Current-RCP4.5 | Current-RCP8.5 | ||
|---|---|---|---|---|
| 2050 | 2070 | 2050 | 2070 | |
| Increased | 2.41 | 3.24 | 3.74 | 5.80 |
| Decreased | 2.36 | 3.17 | 2.78 | 3.84 |
| Unchanged | 14.53 | 13.72 | 14.11 | 13.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Huang, Y.; Zhao, H.; Yang, M.; Zhuang, Y.; Ye, X. Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China. Forests 2021, 12, 429. https://doi.org/10.3390/f12040429
Xu Y, Huang Y, Zhao H, Yang M, Zhuang Y, Ye X. Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China. Forests. 2021; 12(4):429. https://doi.org/10.3390/f12040429
Chicago/Turabian StyleXu, Yadong, Yi Huang, Huiru Zhao, Meiling Yang, Yuqi Zhuang, and Xinping Ye. 2021. "Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China" Forests 12, no. 4: 429. https://doi.org/10.3390/f12040429
APA StyleXu, Y., Huang, Y., Zhao, H., Yang, M., Zhuang, Y., & Ye, X. (2021). Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China. Forests, 12(4), 429. https://doi.org/10.3390/f12040429