1. Introduction
Temperature is an important abiotic factor that influences insect life-history processes, such as survival, development, and reproduction. Understanding how insects respond to temperature makes it possible to estimate potential geographic ranges and develop phenology models to predict the timing of stages that are critical to deploying survey and management options. It is particularly important to develop these predictive tools for species like
Anoplophora chinensis (Forster) (Coleoptera: Cerambycidae) that become invasive after being transported, either in live plants or solid wood packaging materials, through international trade routes. The native range of
A. chinensis largely overlaps that of its congeneric,
Anoplophora glabripennis (Motschulsky), but it has been recorded from warmer areas of southern Asia and is not found in some of the more northern parts of China and far east Russia [
1].
Anoplophora chinensis is widely distributed in China, Japan, and Korea, and occasionally found in Vietnam, Malaysia, Indonesia, and the Philippines [
2,
3]. It is a polyphagous xylophage that attacks 73 different deciduous tree genera (more than
A. glabripennis) and also is reported to attack some coniferous trees in the genera
Cryptomena and
Pinus [
4]. The beetle is the most important wood infesting citrus pest in Japan and China, and also causes major damage to
Casuarina equisetifolia L. shelterbelts in China [
5,
6,
7]. In its invaded range,
A. chinensis prefers trees in the genera
Acer,
Betula, and
Corylus but has completed development on 24 genera and partial development on another five genera of trees [
3,
8]. Females lay their eggs in sites they chew in the bark of the lower bole or exposed roots of the tree, and the early instars feed under the bark, while later instars feed in the xylem [
3]. The larvae cause damage to the vascular system of the tree and create structural weaknesses that can lead to the death of the tree. Adults are present from the late spring through the summer and into the early fall, depending on the local climatic conditions.
A recent revision of
Anoplophora considered
Anoplophora malasiaca (Thomson) to be a synonym of
A. chinensis based on shared morphological traits of the reproductive system [
2]. There is, however, still controversy over this synonymy, since they can be differentiated by whether or not two white spots are present on the pronotum and by their mitochondrial COI region haplotypes [
9,
10]. The first invasive population of
A. chinensis was found in the Lombardy region of Italy in 2000, but in the last 20 years, additional infestations have been found in Italy, France, the Netherlands, Croatia, and Turkey [
11]. The infestations in the Netherlands and Croatia have been traced to live
Acer palmatum Thunberg imported from China, while the one in France started with beetles that emerged from bonsais imported from Asia [
11,
12]. A find outside a greenhouse in Tukwila, Washington, United States in 2001 also came from imported bonsais [
13]. Using the mitochondrial DNA haplotypes, the Italian infestations from the Lazio and Lombardy regions were found to be similar to Japanese populations, while those from Tuscany were closer to Chinese populations [
14]. Therefore, the invasive populations may originate from areas where the two original species are found and could be different biologically in their responses to temperature and other traits. Since all invasive infestations or potential ones (Washington State, US) have resulted in the establishment of eradication efforts, it is important to know if there are differences between these original species. For simplicity,
A. malasiaca will be referred to as “
A. chinensis from Japan”.
There is some information on the temperature effects on the life-history of
A. chinensis adults and eggs. At natural temperatures on citrus (twigs for food and bolts for oviposition) adults from a Japanese population of
A. chinensis survived 77.6 ± 20.3 days, maturation fed for 9.8 ± 1.4 days, and females laid 193.8 ± 65.2 eggs (peaked at 30 days post eclosion and 90% of the eggs hatched) [
7]. Another study using Japanese adults found differences in maturation feeding (feeding required to reach sexual maturity), longevity, and fecundity at 24 °C when different hosts were used [
15]. Fecundity ranged from 60–140 eggs when another group of Japanese females was offered
Acer,
Citrus, and
Platanus bolts to lay on in outdoor cages [
10]. When Chinese
A. chinensis females were fed on
Melia azedarach L., on average, they laid 176 eggs and lived 92 days (males 103 days), but they only laid 56 eggs and lived 62 days (males 63 days) when fed
Casuarina sp. [
1]. Finally, it was estimated that a total of 184 degree-days is required for 50% of Japanese
A. chinensis eggs to hatch when using the estimated
Tmin of 6.7 °C [
7].
In this study, we determined the response of A. chinensis adults and eggs to a range of constant temperatures. Then the mathematical relationships between temperature and adult survival, reproduction, and egg hatch were developed for A. chinensis to facilitate their use in developing phenology models and to compare them to what has been previously reported for A. glabripennis. Finally, the implications for management and eradication programs are discussed.
2. Materials and Methods
2.1. Populations and Temperatures
The populations used in this study were established from larvae that were shipped on 26 February 2018, from the USDA Agricultural Research Service (ARS), European Biological Control Laboratory in Montferrier-sur-Lez, France, under permit to the USDA Forest Service quarantine facility in Ansonia, CT. Fifty larvae from the fifteenth ARS laboratory generation of an invasive A. chinensis population from Lombardy, Italy (41.58° N and 87.42° W) were used to start the USDA Forest Service (FS) colony. The Italian colony was initiated by ARS in 2002 and augmented each year with freshly collected adults. The Italian colony was determined to have originated from Japan based on molecular tests. Individuals used in this study were from the sixteenth or seventeenth Italian laboratory generations.
Fifty larvae from the fifth ARS generation of a native A. chinensis population from Yancun, Guangdong Province, China colony (40.45° N and 73.45° W) were used to start the FS colony. The Chinese population was started by ARS in 2012 with 180 larvae from China. Individuals used in this study were from the sixth or seventh laboratory generations. Voucher specimens of both populations were deposited at the Entomology Division, Yale Peabody Museum of Natural History, New Haven, CT.
Eight constant temperatures were used for these studies: 5, 10, 15, 20, 25, 30, 35, and 40 °C. Temperature fluctuations in the environmental chambers rarely exceeded 1 degree above or below the set value, and a photoperiod of 16:8 (L:D) h was used. Humidity was passively maintained by placing open reservoirs of water (32 × 28 × 14 cm3) in the bottom of the chambers or by humidifiers that were part of the chamber (15, 20 and 25 °C). The humidity in the chambers averaged 80 ± 10%, 85 ± 15%, 70 ± 10%, 80 ± 5%, 60 ± 5%, 45 ± 5%, 68 ± 4%, and 55 ± 10% RH at 5, 10, 15, 20, 25, 30, 35, and 40 °C, respectively. Actual humidity within the rearing containers was likely higher than in the chamber, due to the presence of fresh host material.
2.2. Mating Adults and Obtaining Eggs
Adults used in these studies came from larvae reared on an artificial diet, either the
A. glabripennis diet designated “AG2” [
16] or a drier
A. chinensis diet that was a modification of a host-free diet developed for
A. glabripennis [
17,
18]. Pre-pupae were placed in 50 mL plastic centrifuge tubes (with two pinholes in the lid and a piece of damp paper towel) and held in the dark at 25 °C until pupae formed, and the adults emerged. Callow adults were held for four more days under the same conditions as the pre-pupae/pupae to allow the cuticle to fully sclerotize before they were sexed, weighed, and assigned to a temperature regime. Chinese adults (15 females and 16–18 males) were initially assigned evenly to the 15, 20, 25, and 30 °C temperature regimes, and then ten of each sex were assigned evenly to the 10, 35, and 40 °C temperature regimes. Virgin females in the 15–35 °C temperature regimes were held individually in 3.8-L glass jars with
Acer rubrum L. (red maple) twigs (3–7 mm diameter with leaves removed) as a food source and
A. rubrum bolts (3–7 cm diameter and 22 cm long) with both ends waxed as an oviposition substrate. A virgin male was added to the female’s jar 0–11 days later (i.e., as soon as a female became available at the same temperature). Males that were waiting for a mate were held at their assigned temperatures in 950 mL glass jars with fresh twigs as a food source. All adults held at 10 and 40 °C were held individually in 950 mL glass jars with fresh twigs as a food source. After twelve weeks at 10 °C (presumably enough time for the females to maturation feed), the three males that were still alive were placed with females in the larger glass jars to see if the females would chew pits or lay eggs. Twigs and bolts were cut twice a month and were stored at 4 °C and ≥80% RH until used in the weekly food and bolt changes. Folded paper towels were placed in the bottoms of the jars to collect frass and excess moisture. Two holes (1–2 mm diameter) in the plastic jar lids allowed airflow.
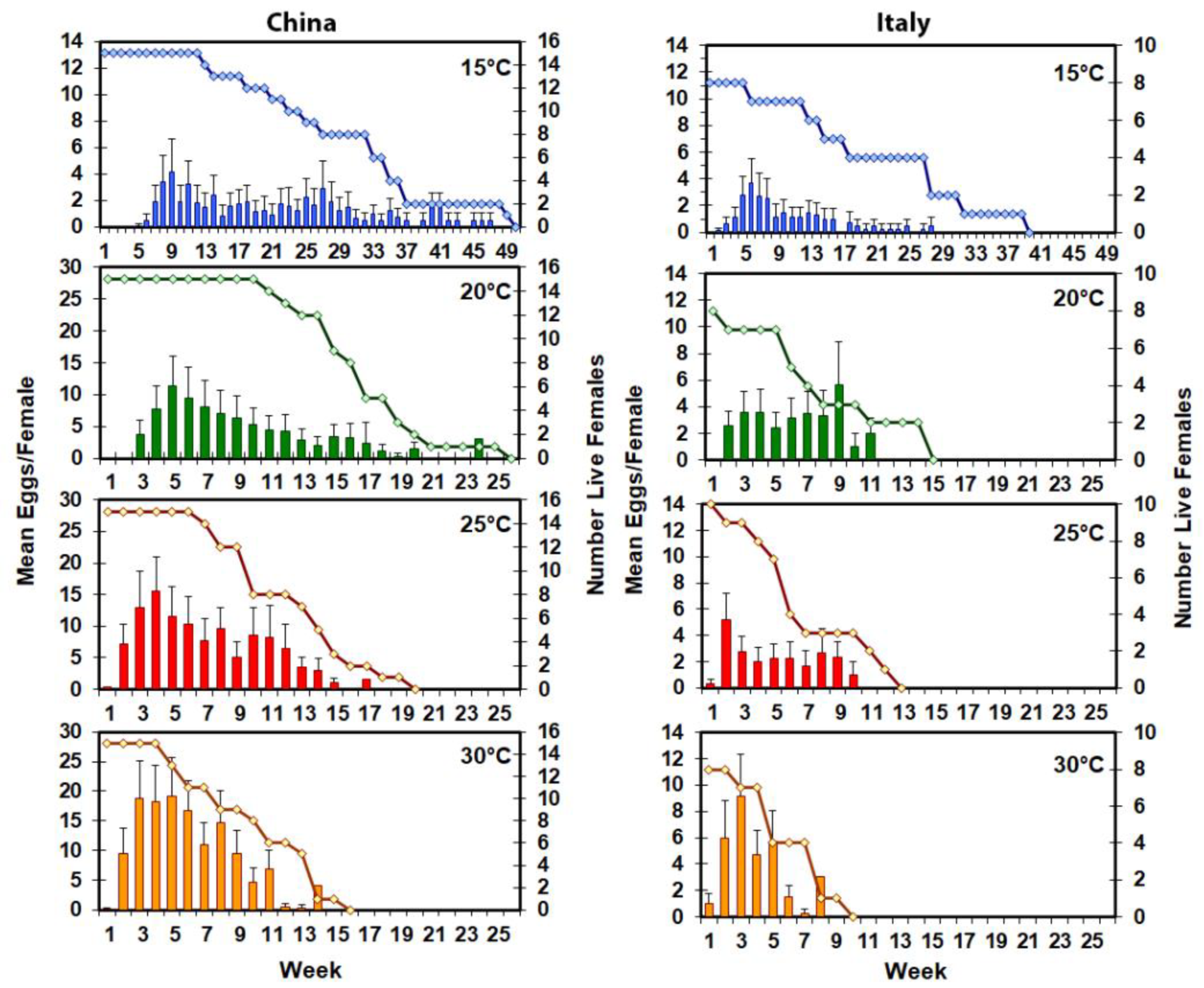
The oviposition bolts were replaced twice a week (30–35 °C), weekly (15–25 °C), or monthly (10 °C) until a female died. Oviposition bolts held at 15–35 °C were checked daily for oviposition sites. The date of each new oviposition pit chewed in the bark was recorded, and the pit marked to distinguish the day it appeared. When the oviposition bolts were removed from the jars, all pits chewed in the bark, including both pits that obviously contained eggs and those that did not, were counted, and the bolt diameter was measured.
The bark was stripped off many of the daily marked bolts the day they were removed from the jar to obtain the eggs for the egg hatch part of the study. After the bark was stripped off, the eggs were carefully removed from the bolt by lifting a thin strip of wood with the egg on it to prevent breaking the eggshell. Eggs attached to the small pieces of wood were placed individually in wells of a 24-well tissue culture plate that was held in a water box at the temperature at which the egg was laid or to which the egg was moved. Ten eggs maximum, or half of the eggs at minimum, that were laid by females at 15 °C were kept at that temperature. For females held at other temperatures, a minimum of 10 eggs was held at the oviposition temperature, and the others were moved as needed.
To minimize temperature shifts for eggs that had already begun developing, one-day-old eggs were only moved to temperatures that were ±5 °C of the laying temperature, but eggs laid the same day were moved to other temperatures as needed. Eggs laid more than one day before the bolt was removed from the mating jar were held at the oviposition temperature. Eggs laid by Chinese females held at 25 or 30 °C were moved to any temperature, while eggs laid at 15 °C were only moved to 20–30 °C and eggs laid at 20 °C were only moved to 10–35 °C. This was because there were fewer eggs laid at the cooler temperatures. Two to ten eggs from a Chinese female were held at each temperature 15–30 °C, and 1–3 eggs were held at each temperature 5–10 °C and 35–40 °C.
Once enough eggs were obtained from each mating pair for the egg hatch part of the study, the rest of the bolts from each female were held at 25 or 30 °C for 7–10 days to allow the eggs to hatch before stripping the bark off the bolt. This provided a better measure of the percentage of the eggs that were fertile, since removing the eggs could damage or kill them. Eggs were checked daily for 4, 4, 4, 6, 9, 12, 30, and 30 weeks at 40, 35, 30, 25, 20, 15, 10, and 5 °C, respectively. These time periods were chosen to ensure that all eggs that could hatch at a given temperature did. Eggs that had not hatched after the given time period at 5–20 °C were moved to 25 °C and observed for an additional four weeks.
Adults from the invasive Italian population were more limited in numbers, so only 8–10 females and 11–14 males were assigned on a rotating basis to the 15, 20, 25, and 30 °C temperature regimes. All the same feeding and holding methods were used as detailed above for the Chinese population adults and eggs. Since Italian females also laid fewer eggs, their eggs were only held at 15–30 °C. Eggs laid at these temperatures were moved using the same scheme as detailed for the Chinese eggs.
2.3. Adult Survival and Reproduction
Mating pairs and single adults were checked daily for mortality, with the only exception that single adults at 10 °C were checked three times a week. When the female in the pair died, the male was moved to a 950 mL glass jar and maintained until death. When the male in the pair died, the female was maintained in the mating jar until death. For each temperature at which females laid eggs, the mean number of eggs laid per week per live female during that week was calculated. When the female died, she was dissected to determine if she had any fully formed eggs in her ovarioles that she had not oviposited. The following dependent variables were analyzed in PROC GLIMMIX [
19]: Female and male weight at four days, female and male longevity, days to the first egg, fecundity, maximum weekly oviposition, a week at which maximum oviposition occurred, mean weekly oviposition pits per female, mature unlaid eggs at female death, and percentage hatch on bolts held at 25–30 °C. A gamma distribution with a log link was used for all the variables except the percentage hatch for which a beta distribution with a logit link was used and the unlaid eggs in the female at death for which a Poisson distribution with a log link was used. The two populations were analyzed separately for each variable. The model used temperature, population (Italy or China), and the interaction between the two as fixed effects. Differences among means were determined using the Tukey–Kramer test with α = 0.05 [
19].
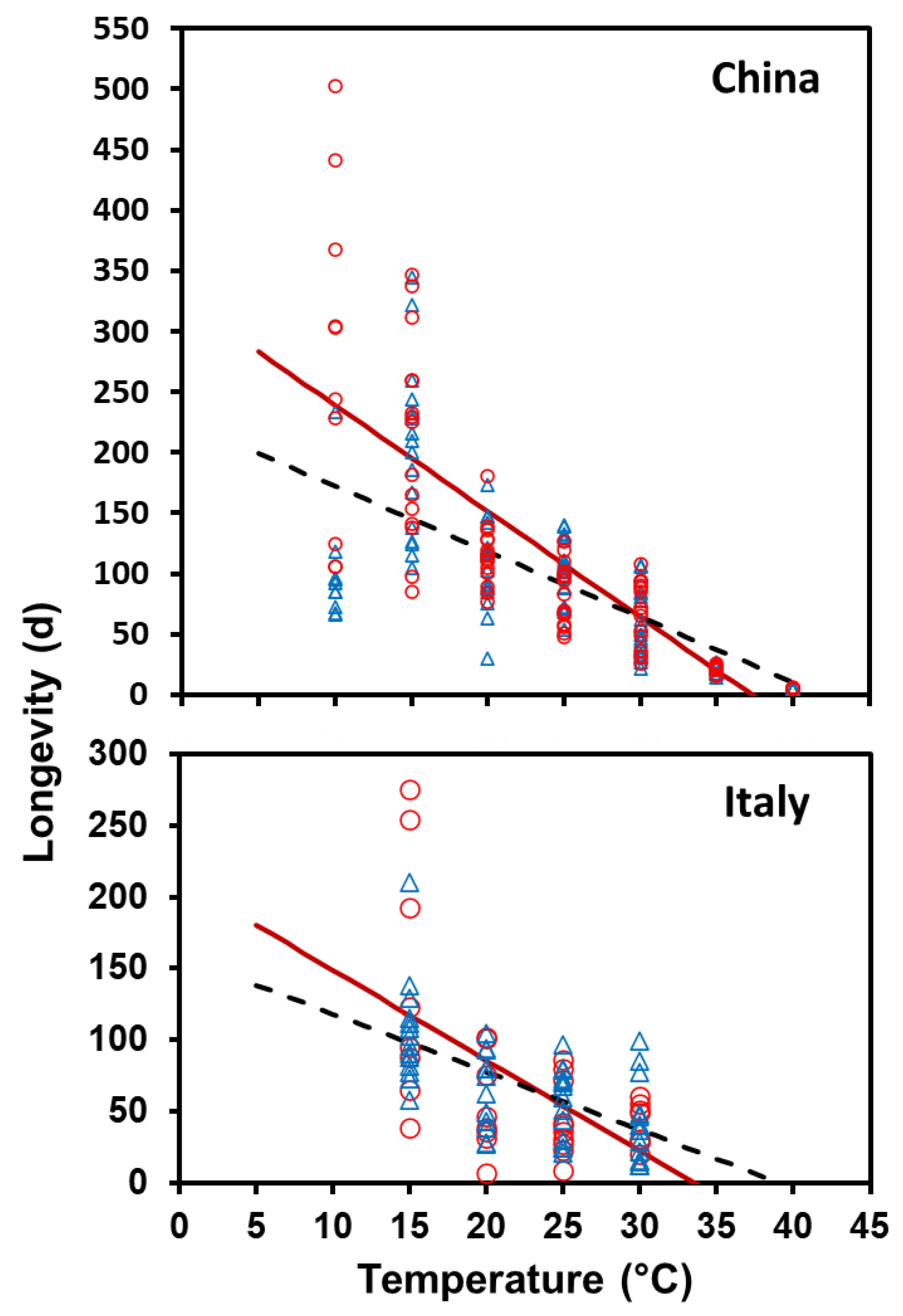
The relationship between longevity and temperature was separately fit for each sex and population combination to a linear model (
y =
bx +
a), and the upper threshold (
Tmax) was calculated to be the point at which the line crossed the
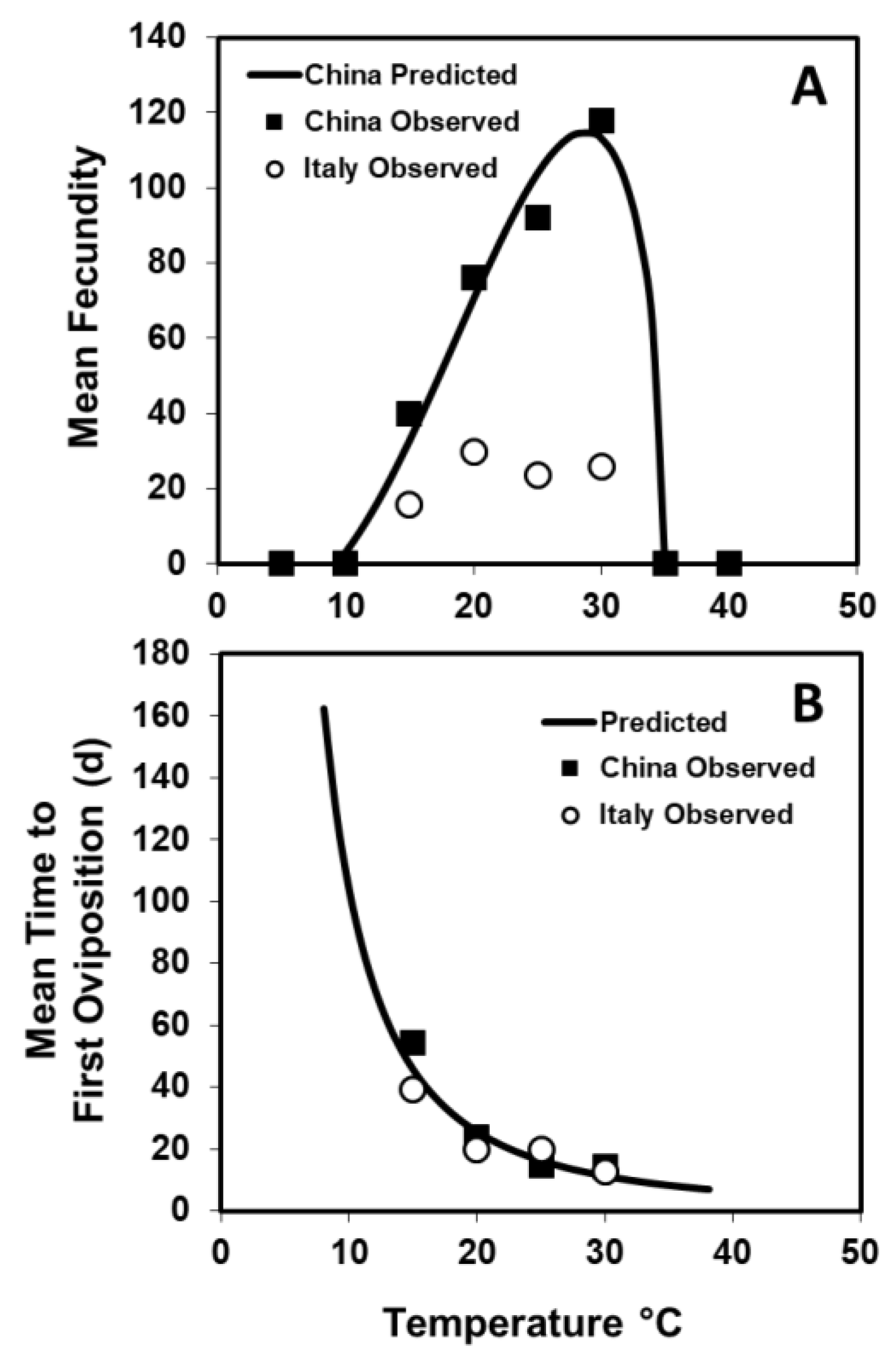
x-axis. The relationship between fecundity and temperature for the Chinese population was fit using the following:
where
y = fecundity,
t = temperature °C are the known values, and the others are estimated. The a is an empirical constant, b modulates the slopes, and
Tmax and
Tmin are the upper and lower temperature thresholds, respectively [
20]. Nonlinear convergence for the model was based on the Marquardt method (PROC NLIN [
19]). The optimum temperature for Equation (1) was calculated by equating the first derivative to zero and solving for t.
The relationship between temperature (
t) and
a number of days from female emergence to first oviposition was fit using the following:
where
a and
b are empirical constants, and nonlinear convergence was based on the Marquardt method (PROC NLIN [
19]). The percentage of females that oviposited and produced progeny was also calculated for each temperature and population combination.
2.4. Egg Hatch
Two dependent variables were analyzed using PROC GLIMMIX [
19]: Days to hatch and percent hatch on bolts held at 25–30 °C. The model treated temperature, population, and temperature by population as fixed effects, whereas maternal family was treated as a random effect. Differences among means were determined using the Tukey–Kramer test with α = 0.05 [
19]. A gamma distribution with a log link was used for days to hatch, and a beta distribution with a logit link was used for the percentage hatch on bolts. The percentage hatch of eggs laid at or moved to various temperatures after being removed from under the bark was also calculated.
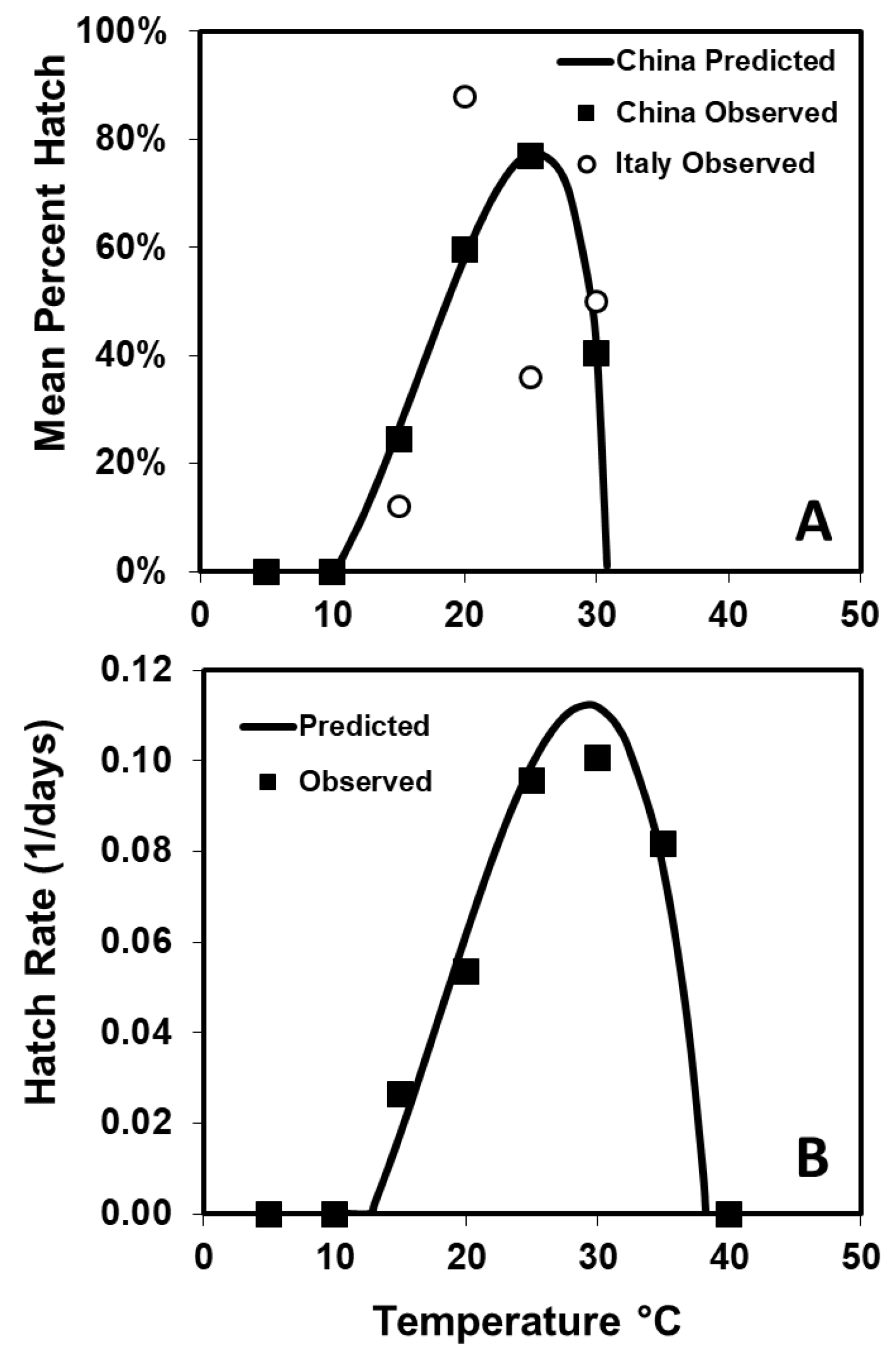
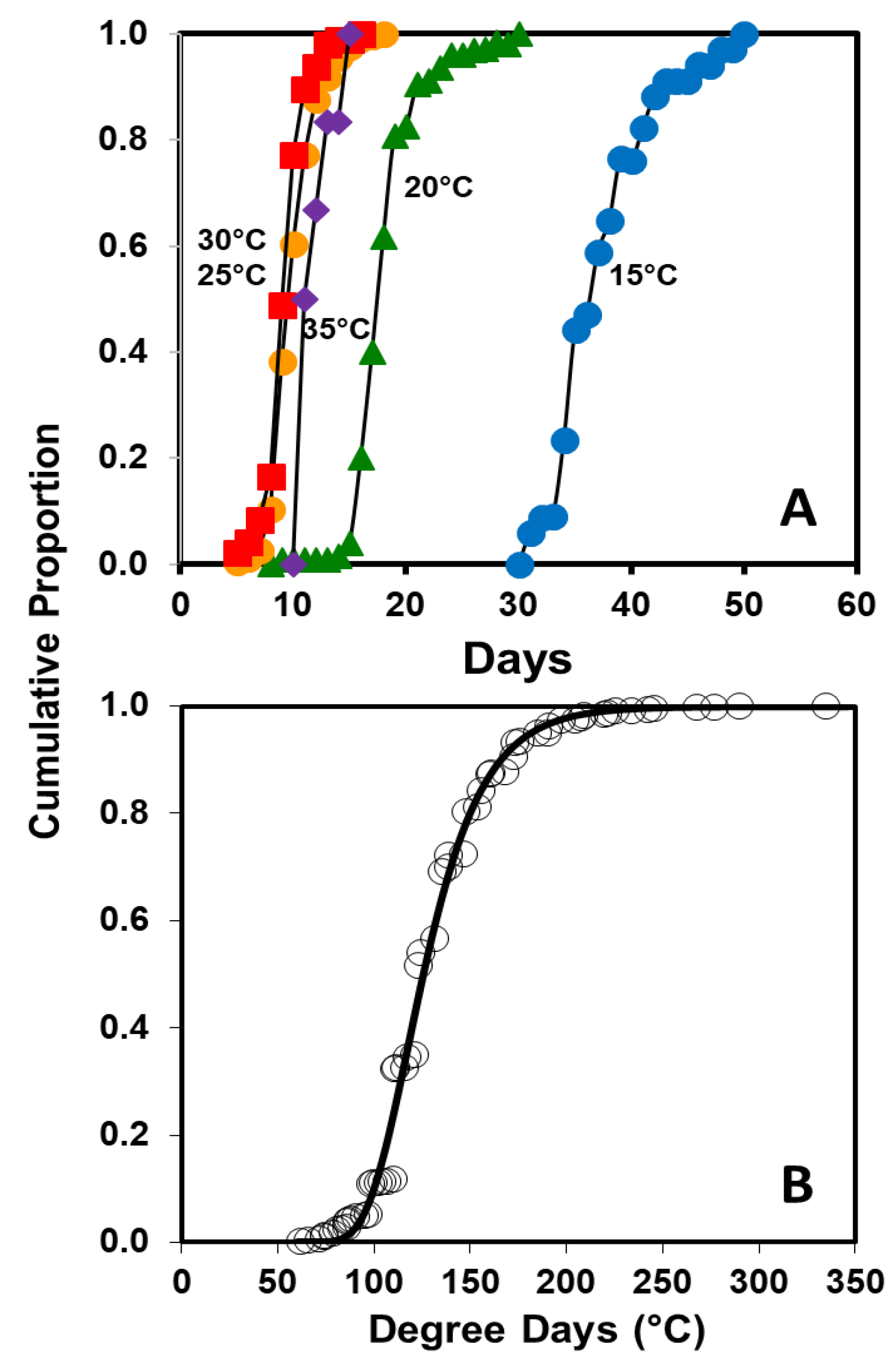
To describe the relationship between temperature and rate of hatch for eggs from the combined populations, Equation (1) was altered by changing
y = 1/days to hatch, and the curve was fit using the Marquardt convergence method (PROC NLIN [
19]). The cumulative distributions of time to egg hatch at 15, 20, 25, and 30 °C and degree days to egg hatch for all eggs were calculated using the estimated
Tmin from Equation (1). The cumulative proportion of eggs hatching (
P) over accumulated degree-days (
DD) was described using a Gompertz function:
in which
a and
b are the lag and the rate of increase, respectively ([
21], PROC NLIN and Marquardt convergence method, [
19]).
4. Discussion
The observed adult activity over the range of temperatures assessed was comparable to what was seen for
A. glabripennis [
22,
23]. Adults held at 15–25 °C exhibited the full range of normal behaviors (resting, feeding, mating, and ovipositing), just more slowly at cooler temperatures. At 10 °C Chinese adults moved very slowly, fed little, and did not chew oviposition sites. At 30–35 °C adult behavior increased, and they would attempt to walk or fly away when their twigs and bolts were changed. At 35 °C, Chinese females did not oviposit even though they chewed some oviposition pits. Chinese adults held at 40 °C were more lethargic, fed little, and did not live long. The attempts to move away from the higher temperatures fit with the observations that
A. glabripennis seeks shaded areas to rest when the temperature is >29 °C in China [
24].
Adult longevity declined with the increase in activity at higher temperatures. This is likely due to increased metabolism at higher temperatures which has been found to reduce the life span in insects [
25]. The longevity at 20–25 °C on
A. rubrum for both sexes from the Chinese population is comparable to that recorded on
Melia azedarach (91.8 days female and 103.0 days male) and higher than recorded on
Casuarina sp. (61.6 days female and 63.0 days male) at room temperature in China [
1]. Conversely, both sexes of the Italian population on
A. rubrum at 25 °C lived for a shorter time than did individuals from a Japanese population at 24 °C on either
Citrus unshiu (Yu.Tanaka ex Swingle) or
Salix schwerinii E. Wolf [
15]. This highlights the effects of host plants on longevity and suggests that caution should be used when applying predictions solely based on a single host in the laboratory. Female
A. chinensis from both populations lived longer than
A. glabripennis females at temperatures ≤15 °C, but comparable lengths of time at higher temperatures [
23]. However, the male longevity of individuals from the Chinese population was comparable to that of
A. glabripennis individuals over the 10–35 °C range [
23]. The longer survival of female
A. chinensis than
A. glabripennis at cooler temperatures is surprising, since the native range of
A. glabripennis extends further north than that of
A. chinensis, but fits with the fact that
A. glabripennis has a higher metabolic rate than
A. chinensis at cold temperatures [
17].
The timing of first oviposition (13–17 days to first egg at 24 °C depending on the host) and the shape of the weekly oviposition curve (bell-shaped with a long right tail) are comparable to the findings of this study for
A. chinensis [
1,
7]. The mean fecundity (92 for Chinese and 24 for Italian at 25 °C), however, was lower than that of either Chinese females on
M.
azedarach (176 eggs) or field-reared Japanese females on citrus (193.8 eggs) [
1,
7]. This is most likely due to the host used in the studies, since the host has been shown to affect fecundity in
Anoplophora spp. [
15,
26]. The increasing number of oviposition sites chewed per week, maximum oviposition per week, and fecundity for females from the Chinese population with increasing temperatures 15–30 °C was to be expected because of increased efficiencies of assimilation and conversion of ingested food at higher temperatures [
27]. What was not expected was a lack of temperature response for reproductive output for the Italian population, and the causes for this are unknown. Compared to
A. glabripennis, the
A. chinensis Chinese females laid more eggs across the entire temperature range of 15–30 °C, and fecundity peaked at a higher temperature [
23]. The higher fecundity could have been because
A. chinensis were, on average, heavier than
A. glabripennis females, and fecundity has been correlated with size in both species [
7,
28]. The ability of
A. chinensis to handle higher temperatures better than
A. glabripennis is also suggested by its native range extending further south, even though much of their ranges overlap [
2]. At the more extreme temperatures, the pattern of reproductive arrest seen for
A. chinensis is comparable to that found for
Hylotrupes bajulus L. [
29] and
A. glabripennis [
28]. At 10 °C and ≥35 °C, females laid no eggs, and there were no fully developed eggs in their ovarioles at death. This was either due to a lack of egg maturation or eggs being reabsorbed within these females to use as an energy source.
Eggs of most insect species will hatch over a 15 °C range of constant temperatures, and under varying temperatures like occur in nature, egg hatch may occur 3–4 °C below that range [
30]. As temperatures approach (within 2–3 °C), the
Tmax and
Tmin, percentage hatch declines dramatically [
30].
Anoplophora chinensis eggs from the Chinese population appear to be an exception, as eggs hatched over a broader range of constant temperatures (15–35 °C), although the predicted
Tmin and
Tmax were only 3 °C below the temperatures at which hatch percentage declined sharply. However, the cold tolerance of
A. chinensis eggs seems to be less than that of
A. glabripennis, since none of the
A. chinensis eggs that were held at 5–10 °C for 200 days and then were moved to 25 °C were able to hatch, while eggs of
A. glabripennis did hatch under similar conditions. This would suggest that
A. chinensis eggs would not be able to overwinter, unless the more sheltered tree bole and roots location they use provides more protection from colder temperatures than is afforded
A. glabripennis eggs laid under the bark up higher on the tree. However, care should be taken in making this assumption, since removing the eggs from the bark can damage
A. chinensis eggs, and we did not confirm this by holding eggs under the bark at the low temperatures.
The percentage of Italian eggs laid at 20 °C and hatched at 25 °C was similar to that for Japanese
A. chinensis eggs laid and hatched at 20 °C in
Citrus sp. Bolts (90%) [
7]. The lower hatch percentages seen for eggs removed from bolts was probably due to damage sustained when removing them, since there were reductions for all temperatures, and the eggshells of
A. chinensis broke more easily than those of
A. glabripennis. The percentage hatch of the Chinese population eggs kept on bolts was comparable to
A. glabripennis egg hatch when removed from bolts at 15–30 °C. However, unlike
A. glabripennis, some
A. chinensis eggs that were removed from bolts were able to hatch at 35 °C, possibly another indication that
A. chinensis may be better adapted for warm climates than
A. glabripennis.The predicted number of degree-days to 50% hatch of
A. chinensis eggs was 166
DD which is comparable to previous estimates for Japanese
A. chinensis (184
DD) [
7], but considerably less than the 239
DD required by
A. glabripennis [
23]. The
Tmin of 12.7 °C is higher than the 6.7 °C estimated for Japanese
A. chinensis and slightly higher than the 9.7 °C estimated for
A. glabripennis [
7,
23]. In addition, the
Tmax is quite a bit higher for
A. chinensis than
A. glabripennis. The lack of hatch for
A. chinensis eggs held at 10 °C substantiates the
Tmin estimated in this study and suggests that either the previous estimate was low or that some Japanese populations of
A. chinensis may have a lower threshold.
Diurnal and seasonal temperature variations in the lower bole and exposed roots of trees where
A. chinensis eggs are laid could be substantially different from ambient or from tree trunks and branches where
A. glabripennis eggs are laid. Previous studies have shown that phloem temperatures in the upper bole and branches can be much higher than ambient (averaging 2–10 °C higher and maximum 21 °C higher), especially when the tree section is exposed to direct sunlight [
31,
32]. However, other factors can also alter within-tree temperatures, such as reflected and thermal radiation, wind chills, and soil temperature around the roots. In fact, the deeper in the trunk or under soil, the more insulated the wood is, and the slower the temperatures may respond to ambient changes [
32]. When using the estimated
DD and
Tmin for predicting when
A. chinensis eggs will hatch, ambient air temperatures will likely not provide accurate estimates. Further work to determine exactly how air temperature differs and temperature changes diurnally and seasonally in the root zone of trees is needed to accurately predict egg hatch timing and potential effects on percentage hatch for
A. chinensis.The impact of temperatures on the adults can be seen if the temperatures from the most northern and southern invasive
A. chinensis populations in Europe are compared. Monthly mean temperatures in Rome, Italy are 17, 21, 23, 24, 21, and 17 °C during May, June, July, August, September, and October, respectively, while the corresponding temperatures in Rotterdam, Netherlands were 12, 15, 17, 17, 14, and 11 °C, respectively (
https://weatherbase.com, accessed 2 April 2021). For Rome, Italy, during the same time period, the maximum temperatures averaged 22, 26, 29, 29, 26, and 22 °C, the maximum recorded temperature was 37 °C for July-September, and there are, on average, four days a year that the temperature exceeds 32 °C. Thus, Rome experiences temperatures at which the beetles can survive and reproduce well from June to August, and summer temperatures rarely reach >32 °C, so it should not reduce adult survival or suspend oviposition. Average temperatures in Rotterdam, however, barely reach levels at which beetles will oviposit June–August, and even high averages only reach ≥20 °C in July and August—so although the beetles may survive a long time, they will lay few eggs. This may partially explain the total of 25 exit holes seen in the Westland, Netherlands, infestation and the relative ease of eradicating that infestation, while the infestation in Rome is larger and more difficult to eradicate [
11]. Average summer temperatures in major citrus-growing areas in California like Visalia would be near-optimal, but average highs and maximum recorded temperatures could suspend oviposition and reduce adult longevity part of the time. Citrus-growing areas in Florida (e.g., Bartow) would be near-optimum for beetles, and even the maximum recorded temperatures would rarely reach the
Tmax for adult survival or result in the suspension of oviposition. These data on temperature effects on reproduction and survival of adults and eggs of
A. chinensis, if combined with temperature response data for larvae, could be used to develop maps of the potential geographical range for this species and stage-based phenological models to predict the timing of life stages. Such information is critical for informing management and eradication programs, both in the native and introduced ranges.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}