1. Introduction
Windstorms cause serious damages to forests in most European countries, especially in Western, Central and Northern Europe [
1]. Its impact is evident also in the Western Carpathians (mostly situated in Slovakia), where the area of damaged forests by windstorms accompanied by bark beetle outbreaks has a gradually increasing tendency especially since the beginning of the new millennium [
2]. Generally, increasing frequency of strong winds and their growing impacts on forests in these European regions are mostly interpreted as an inherent phenomenon of climate change [
3] and accumulating amount of aboveground tree biomass [
4], which can together with some other aspects worsen forest stability [
5]. The main consequence of recent windstorm disturbances is the increasing windthrown area, later covered by young forests. This is the principal reason why current scientific attention should focus on post-disturbance areas and forest development to a larger extent than in the past.
Under the conditions of boreal and temperate Europe, Norway spruce (
Picea abies L. Karst) is considered the tree species most prone to wind damage [
6]. At the same time, the species has long been economically extremely important [
7]. Since the regeneration potential of this species is usually high (thanks to seed-producing survivors after disturbance and well-dispersing and germinating seeds), spruce trees often dominate in species composition of young post-disturbance forests [
8]. At the same time, in disturbed forest areas, seeds of different species—especially light-demanding, are granted an opportunity to germinate and cover the open soil surface [
9]. In the case of large-scale disturbances, “pioneer tree species” play an important role in forest regeneration, which are in Slovakia especially silver birch (
Betula pendula Roth.), goat willow (
Salix caprea L.) and common rowan (
Sorbus aucuparia L.; see for instance Konôpka et al. [
10]).
On the Slovak forests, the largest windstorm disaster occurred on 19 November 2004 [
11]. The negative feature of the episode was that the epicentre of wind destruction was located in the Tatra National Park (TANAP), which is the oldest and most famous national park in Slovakia. At that time, wind damaged nearly 130 km
2 of forests dominated by Norway spruce [
12]. Our previous work [
13] indicated that Norway spruce and birches (
Betula sp.), especially silver birch (
B. pendula Roth.), were most frequent and most contributing tree species to forest biomass stock in the post-disturbance area of TANAP. These two species have very contrasting qualities, out of which some are well-known, such as foliage traits (leaves vs. needles, deciduous vs. evergreen), lifespan, ecological demands (e.g., drought and light tolerance). Due to the above-mentioned differences, we assume that the species differ also in biomass production and allocation, which have not been examined in the form of an inter-specific comparative study yet.
We investigated these two tree species under post-disturbance conditions because both prominently contribute to forest regeneration [
13] and thanks to their contrasting properties they may complement each other at the mutual site. For instance, since in initial developmental stages birch grows faster and tolerates extreme climatic conditions better than spruce [
14], birch trees often create a “shelter” and make favorable conditions for spruce growth [
15]. Although unlike in Northern Europe birch species are not important for commercial purposes in Slovakia, their ecological advantages, as well as their production potential, can be exploited in the context of disturbances and global warming environment (e.g., see Reference [
7]). Previous works showed that in early developmental stages birches are usually more productive than most other forest tree species on post-disturbance [
13] or former agricultural lands [
16,
17]. Wood of birches, but also of other pioneer tree species regenerated on disturbed areas, will very probably become a source of supply for production of renewable energy [
18]. This may reduce demands for wood of “traditional” commercial tree species, such as Norway spruce.
Thanks to fast growth, birches can play an important role in carbon sequestration at sites where other tree species hardly survive. Experience from Scandinavian and Baltic countries [
19,
20] suggests that birches can be economically relevant species with a variety of utilization possibilities. According to Tiebel et al. [
21], renewed interest in silver birch has been recently raised with regard to forest management at higher altitudes. The main reason for it is the ability of birch to promptly and extensively re-colonize post-disturbance areas (mostly after windfalls and bark beetle outbreaks), which is an extremely important quality in the context of the increased risk of catastrophic events in European spruce-dominant forests. At the same time, such a development creates conditions for mixed forests stands with higher species richness. Likely, Norway spruce might be then preferred exclusively only on autochthonous sites, i.e., at high altitudes with mostly sufficiency of precipitation. That is besides other ecological aspects important in terms of the ongoing climate change [
22].
The aim of the paper was to compare contribution of tree components to aboveground biomass between silver birch and Norway spruce growing at the identical site, specifically in the post-disturbance area of TANAP. Further, we focused on the inter-specific comparisons from the point of their wood density, diameter increment and aboveground biomass production. The main mission of our study was to widen the knowledge base related to the potential (especially in wood production and carbon sequestration) of silver birch as an alternative or additive species to Norway spruce in young forest growth stages.
2. Materials and Methods
2.1. Research Site and Transects
Our field activities were performed in the High Tatra Mts., which are a part of TANAP, Northern Slovakia. The bedrock of the High Tatra Mts. is predominantly represented by granodiorites sediments. The soils in the mountains are mostly lithic leptosols, cambisoils and podzols. The climate is typically cold with annual mean temperature around 5.0 °C, annual precipitation totals over 1000 mm, and snow cover of nearly 120 days.
Currently, the main part of the TANAP territory is covered by post-disturbance forests which have been growing there since the large-scale windstorm episode on 19 November 2004. On that day, the wind destroyed spruce-dominated forests at elevations from 700 to 1400 m a.s.l. The destroyed forests occurred within 3–5 km wide and nearly 35 km long belt [
13]. The forest stands inside the windstorm epicenter were almost completely destroyed (more often uprooted than stem-broken) except for a few forest clusters dominated by European larch (
Larix decidua Mill.) and Scots pine (
Pinus sylvestris L.).
Post-disturbance management was differentiated based on the degree of nature protection: from salvaging all amount of merchantable wood in the parts with the lower degree of nature protection, through partial processing of calamity wood, up to complete exclusion of salvage harvesting with exclusive natural succession in the parts under the higher degree of nature protection. The substantial amount of the calamity wood was logged during the years 2005–2006. Similarly, various approaches were implemented for reforestation respecting the degree of nature protection. While natural regeneration was generally preferred, combined natural and artificial reforestation occurred at some sites, and in a few exceptional cases forests were reforested exclusively by tree planting.
Our research started in 2016 (i.e., twelve years after the wind disturbance), when the area was prevailingly covered by young forests that originated from both natural regeneration and planting. Open areas that occurred within the young forest stands were overgrown by other plant species adapted to forest clearings, particularly by grasses, herbs, and shrubs. To acquire the information on tree characteristics, two research transects—one near the site called “Danielov dom” (hereinafter as DD transect) and another one close to the village of Horný Smokovec (HS transect) were established. The forests at these localities are managed by the State Forests of the Tatra National Park. Both sites belong to the territory with the lowest degree of nature protection. Hence, the post-disturbance management at these sites was performed as full-area processing of calamity wood and combined (tree planting and natural regeneration) reforestation. Actually, the entire forest management, i.e., from forest regeneration to harvest, in this area is very close to “ordinary” forestry approaches in Slovakia promoting both wood production and other ecosystem services. On the other hand, forest management is limited or fully excluded only in the core area of the Tatra National Park (not our case), specifically at high altitudes typical with alpine vegetation, and in deep valleys with old-growth forests.
Both transects are located in the central part of the wind-disturbed belt. Their orientations are from Northwest to Southeast. The altitudes of the DD and HS transects varied from 970 to 1000 m a.s.l and between 920 and 950 m a.s.l., respectively. The transects are 4 m wide and 300 m long. Detailed data from both transects have already been analyzed in our previous works, which thoroughly describe their stand properties [
13,
23].
For our current study we used only data covering tree heights and stem breast diameters (diameter at breast height (DBH)) of Norway spruce and silver birch trees with DBH ≥ 7.0 cm. The minimum DBH of 7 cm was chosen since in Slovak forestry this dimension is a standard threshold used in forestry evidence to calculate stand stock of merchantable wood and other related forest characteristics. Tree heights were measured with a hypsometer TruPulse 360°R with a precision of ±0.1 m, and DBH with a digital caliper Masser BT, precision ± 0.1 mm. The data on tree heights and DBHs of spruce and birch trees measured in the years 2016, 2017, 2018, 2019 and 2020 were utilized for further calculation and modeling. Since additional trees reached the DBH limit of 7 cm in consecutive years, the set of measured trees inter-annually increased (
Table 1). While in 2016, 114 birches and 42 spruces were measured, in 2020 we measured 153 birch and 227 spruce trees. Here we would like to point out that both measured and sampled trees (described in the following chapter) represented the upper tree layer of stands. At the same time, it is necessary to note that trees did not create a closed canopy, but rather sparse forests with mild crown competition.
2.2. Aboveground Tree Biomass Sampling
At the end of 2019 growing season, i.e., nearly 15 years after the wind disturbance, destructive collection of aboveground tree biomass was performed. We selected 20 spruce and 20 birch trees with predominant, dominant or co-dominant social position (Kraft classes [
24]) growing in the surrounding of the DD transect. Each selected tree was cut with a chain saw at the ground level. Tree age was determined as number of annual tree rings visible on the cross section of the stem base. Tree height and DBH were measured with metal measuring roller (precision of ±1 mm) and a digital caliper Masser BT (precision of ±0.01 mm). Then, branches with foliage were cut off from the stem and the stem was divided into 100 cm–long sections. These components were packed separately to labeled paper bags and transported to laboratory.
In laboratory, diameters of stem sections were measured with a digital caliper (precision of ±0.01 mm) at both ends in two perpendicular directions. Bark was peeled off from stems with a knife, and diameters of stem wood were measured in the same way as before debarking. Stem wood and stem bark of individual trees were packed in labeled paper bags and dried in a large-capacity drying oven, Komeg KOV-1500L, to reach the constant weight (under a temperature of 95 °C for 120 h). Dry material of stem wood and bark was weighed by using a digital scale, Radwag, WLT 3/6/X (precision of ±0.1 g). Branches with foliage were left in a well-ventilated room for a couple of weeks. Then, foliage was manually separated from branches (nearly all spruce needles and a part of birch leaves shed from branches themselves). Foliage and branches were dried in the drying machine, to reach the constant weight (under a temperature of 95 °C for 96 and 72 h for foliage and branches, respectively) and weighed.
2.3. Calculations, Modeling and Statistical Approach
Stem volumes under and over bark (
VSUB,
VSOB) were calculated for each sampled tree as a sum of volumes of all 100 cm–long stem sections. Volume of one stem section was calculated by using Newton’s formula:
where
Vij is volume of stem section
i of tree
j (cm
3),
L is a length of stem section (cm),
Ab is a cross-sectional area at the bottom end of the section (cm
2),
Am is a cross-sectional area in the middle of the section (cm
2) and
As is a cross-sectional area at the top end of the section (cm
2).
Bark volume was calculated as a difference of stem volume over bark and stem volume under bark.
Stem wood density and stem bark density were calculated as ratios between the respective biomass weight (of wood or bark) and the corresponding volume. Under the term “wood density” we understand here basic wood density,
ρ, calculated as a ratio of oven-dry wood mass,
m0, to its green volume,
Vmax:
where
ρ is basic wood or bark density (kg m
−3),
m0 is oven-dry wood or bark mass (g),
Vmax is green wood or bark volume with wood moisture above the hygroscopicity threshold (m
3).
Further, we constructed allometric relations for stem volume and biomass of separate tree components (foliage, branches, stem bark and stem wood), as well as aboveground tree biomass using DBH and a combination of DBH and tree height as predictors. We applied a basic (
Y =
a Xb), as well as an expanded version of an allometric equation as follows:
where
Y is the dependent variable, i.e., either stem volume under or over bark (
VSUB, VSOB in m
3), or biomass (
B in kg) of a particular tree component as defined above;
X1 and
X2 are independent variables, i.e., breast height diameter (DBH in cm) and tree height (h in m);
b0,
b1, and
b2 are model regression coefficients; and
θ is a multiplicative error term.
The relationship between tree height
h and
DBH was described with the following equation:
where
h is tree height (m);
DBH is tree diameter at breast height, i.e., at a height of 1.3 m (cm);
c0,
c1 and
c2 are regression coefficients.
Tree annual production of aboveground biomass, of stem biomass over bark, and of stem volume over bark was calculated as an inter-annual change of the respective variable, using the formula:
where Δ
Y is the tree annual production of a particular biomass, i.e., its annual increment (kg);
DBH1 is the actual diameter at breast height in the first year (cm);
DBH2 is the diameter at breast height in the second year (cm) estimated as the actual diameter at breast height increased by species-specific mean annual diameter increment, i.e.,
DBH2 =
DBH1 + Δ
DBH;
b0 and
b1 are regression coefficients of a particular equation.
Mean annual diameter increment ΔDBH was calculated separately for each tree species from repeated measurements of trees monitored at transects. By accounting for all years of measurements, we obtained mean annual increment of spruce and birch equal to 1.0 and 1.4 cm, respectively.
Species-specific annual production of foliage was derived by using biomass equations as follows:
The formula for the calculation of spruce annual production of foliage is based on the previous experience and knowledge obtained from a similar experiment [
25] that annual foliage production of spruce is approximately one third of the total foliage biomass at the end of the growing season.
To test significant differences between the tree species we used two-way ANOVA followed by Fisher’s Least Significant Difference (LSD) test and t-test (with p < 0.05).
3. Results
Based on the number of annual tree rings at a stem base of the sampled trees we found that in the year 2019 trees were 14 (±1) years old. Hence, they were born in the growing season of 2005 or 2006. Our set of sampled trees indicated that birch trees had greater dimensions than spruce trees (
Table 2). Specifically, in the sample of trees taken for biomass analysis, birches were thicker and higher than spruce trees (mean DBH = 14.98 cm versus 8.9 m, and mean height = 9.80 versus 6.4 m), and rather interestingly the difference in mean aboveground tree biomass between the species was threefold (biomass of a birch and a spruce tree was 72.86 and 24.08 kg, respectively).
Derived allometric relationships for aboveground tree components of birch (
Table 3) and spruce (
Table 4) showed that DBH was a good predictor of their biomass since
R2 values of derived relationships fluctuated between 0.923 and 0.972 for birch and between 0.914 and 0.980 for spruce. Adding tree height to the model improved the model explanatory power only slightly, specifically
R2 values of these relationships were between 0.940 and 0.977 for birch and between 0.917 and 0.992 for spruce depending on the tree component (
Table 3 and
Table 4).
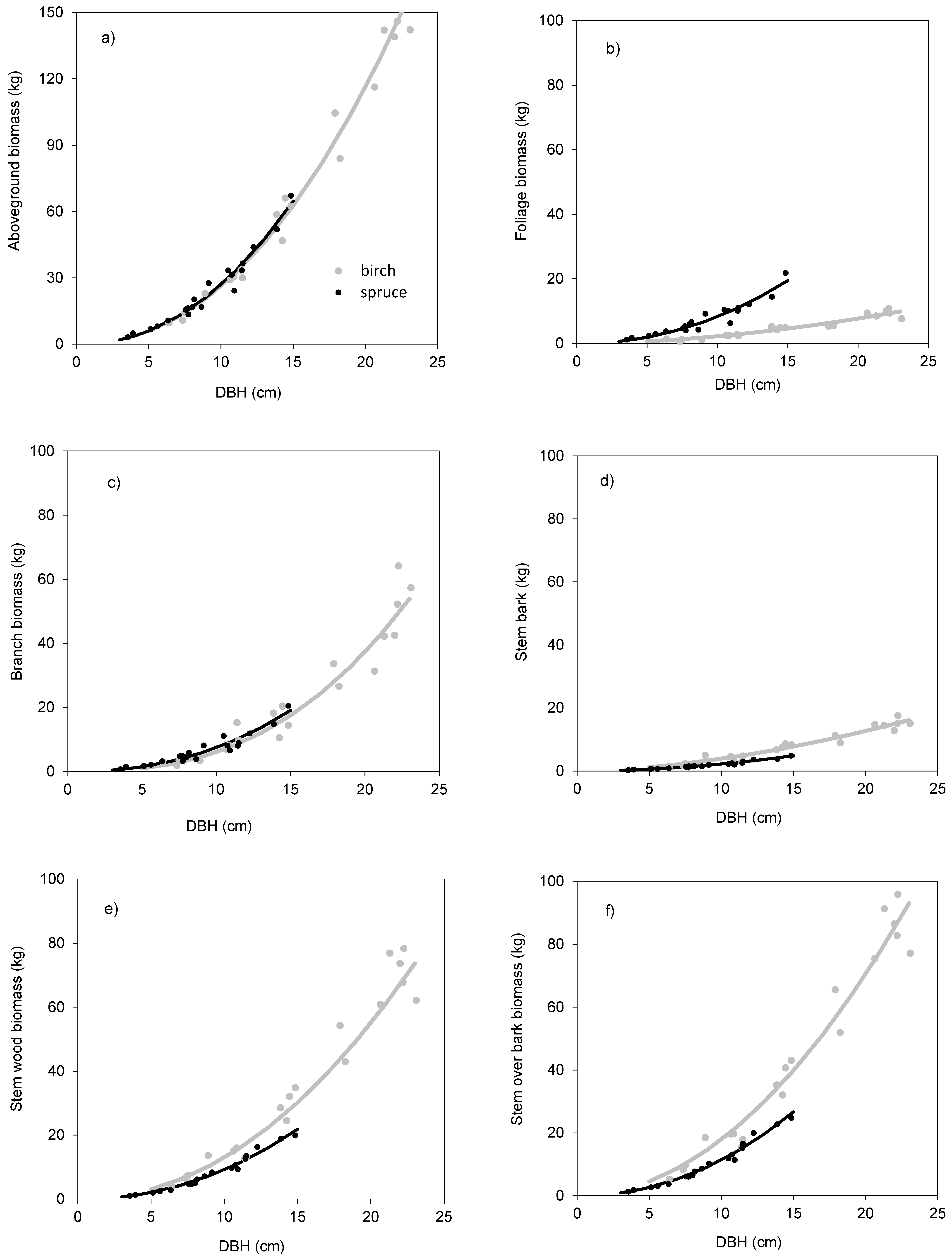
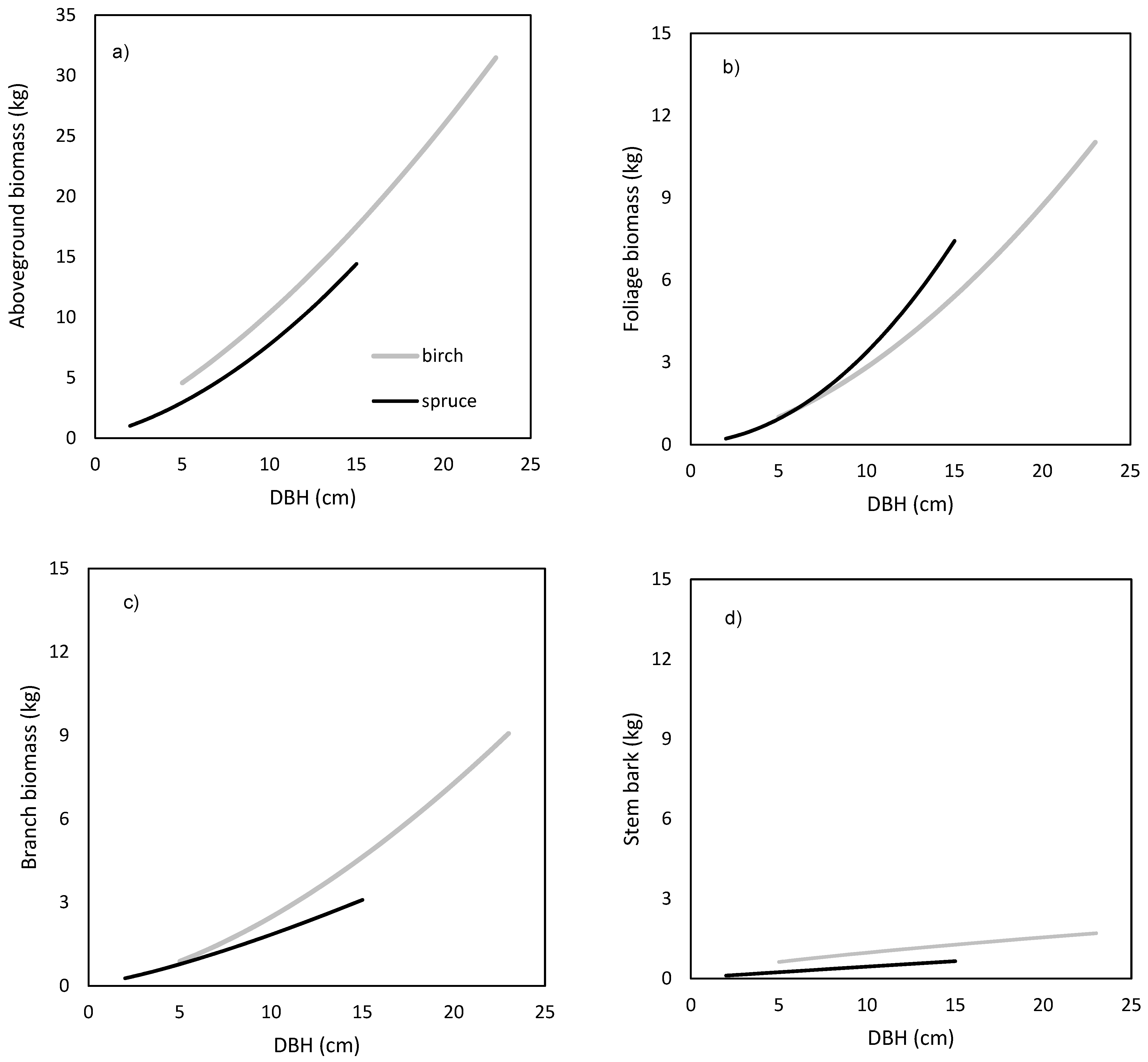
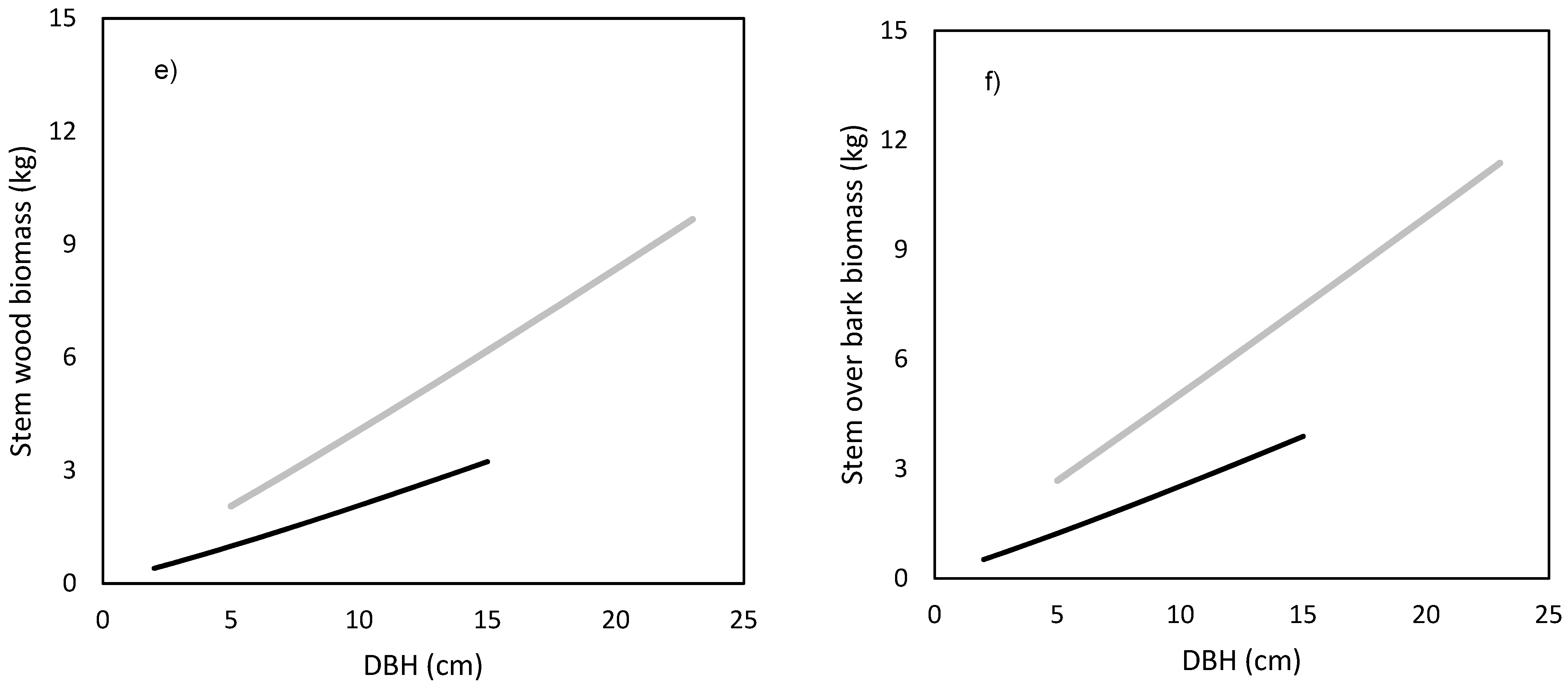
The models showed that birch and spruce trees with identical DBH had nearly the same amounts of aboveground biomass (
Figure 1a). For instance, the aboveground tree biomass of an individual tree with DBH of 15 cm was about 70 kg irrespective of tree species. The derived models showed also very similar branch biomass of both species (
Figure 1c), moderate differences in stem bark biomass (
Figure 1d), as well as in stem biomass under bark (
Figure 1e) and over bark (
Figure 1f), but very contrasting amounts in foliage (
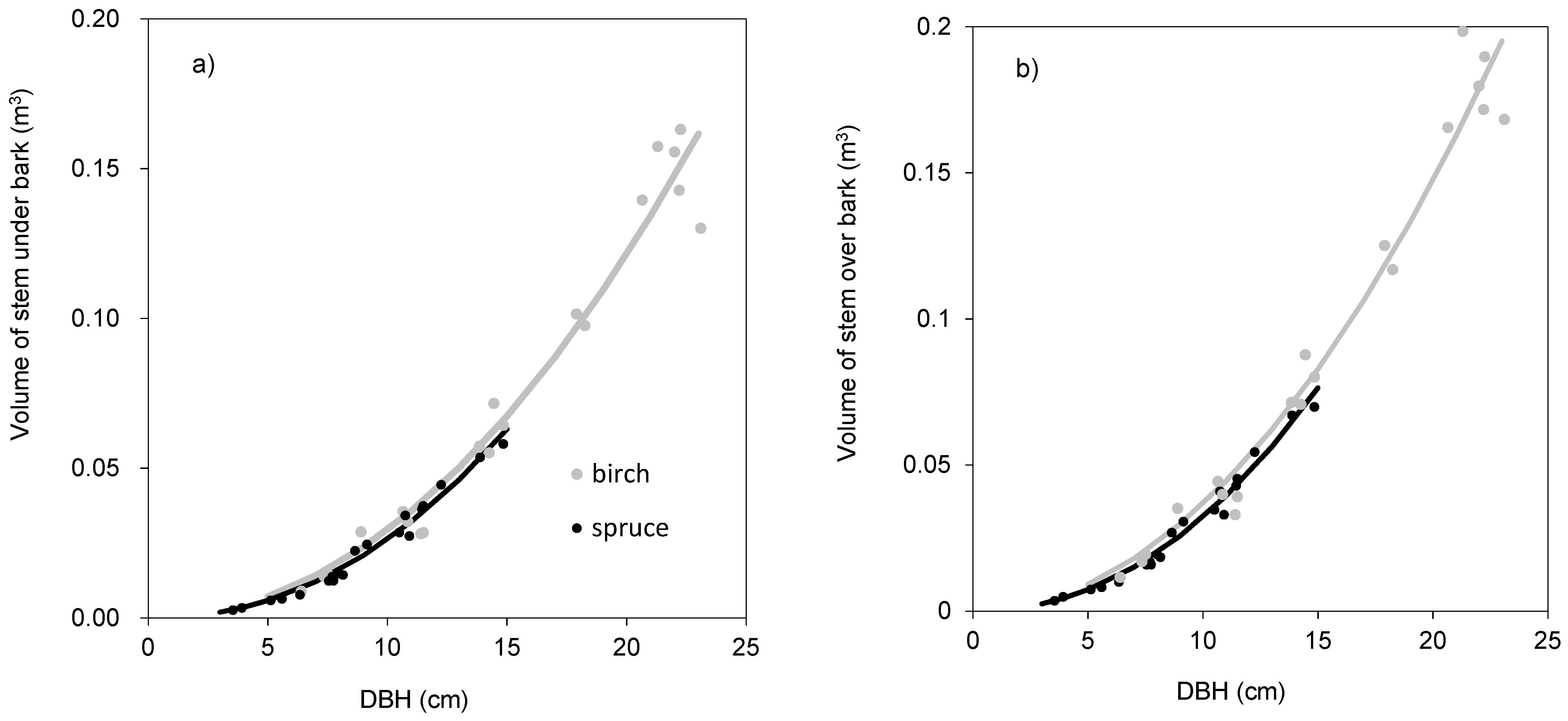
Figure 1b). For instance, a spruce tree with DBH equal to 15 cm had as much as 20 kg of foliage biomass, while the foliage amount of the birch tree with the same DBH was only 5 kg. As for stem volume, the two species had very similar relationships between DBH and stem volume under bark (
Figure 2a) or over bark (
Figure 2b). Derived models for stem volume prediction using DBH as an explanatory variable were significant and explained more than 90% of volume variability for both birch (
Table 5) and spruce (
Table 6). The models clearly showed that for the specific DBH the compared tree species were characterized by almost identical values of stem volume, both under and over bark.
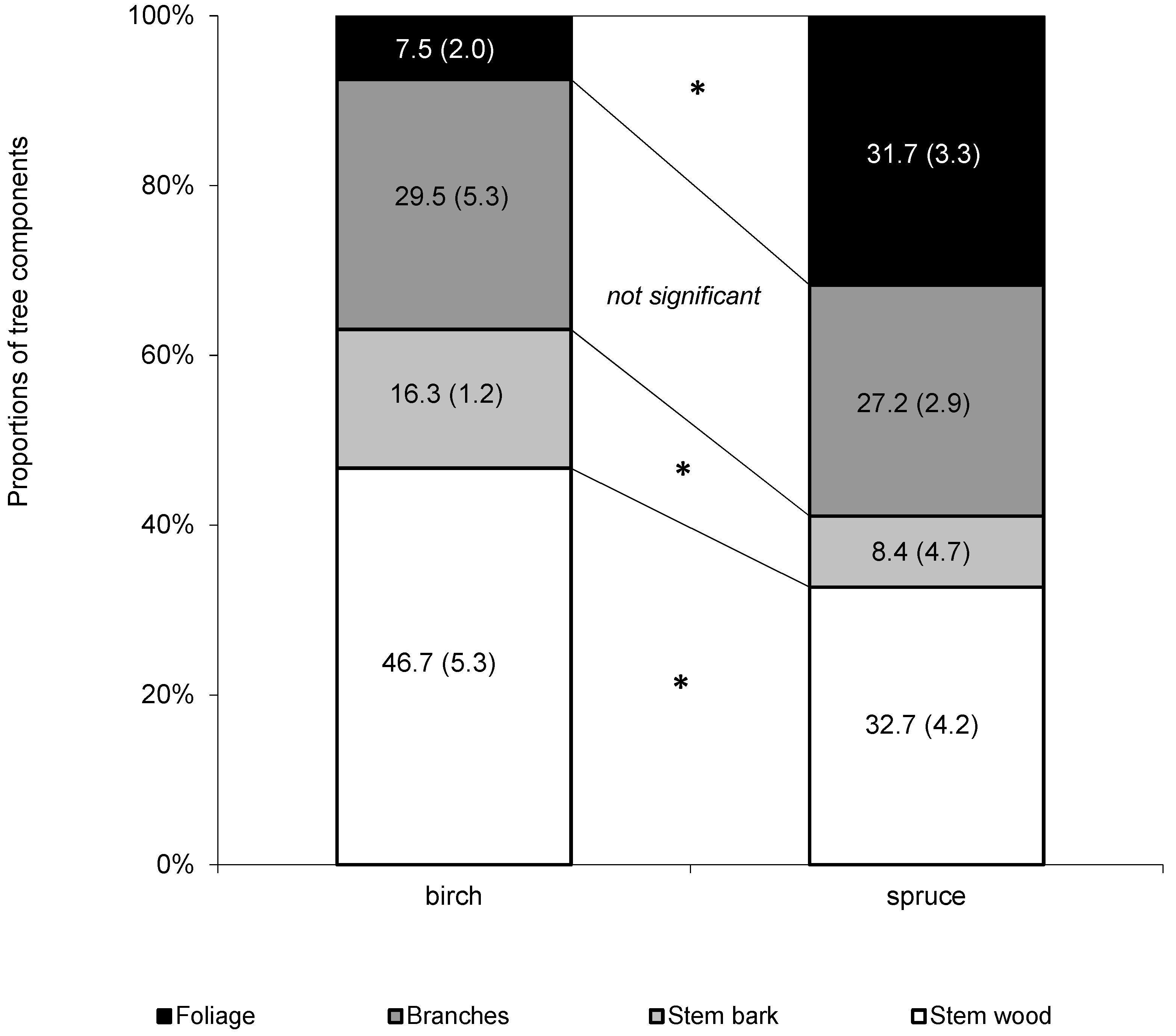
Further, we used the data from the sampled trees to derive mean tree compartments contributions to aboveground tree biomass (
Figure 3). The results showed that only mean contribution of branches to aboveground biomass was insignificantly different between both species (29.5% and 27.2% for birch and spruce, respectively). The difference in foliage contribution was fourfold (7.5% and 31.7% for birch and spruce, respectively), and nearly the double difference was revealed in stem bark (13.3% and 8.4% for birch and spruce, respectively). Stem wood contributed to aboveground biomass most regardless of species, but its contribution in birch was significantly higher (46.7%) than in spruce (32.7%).
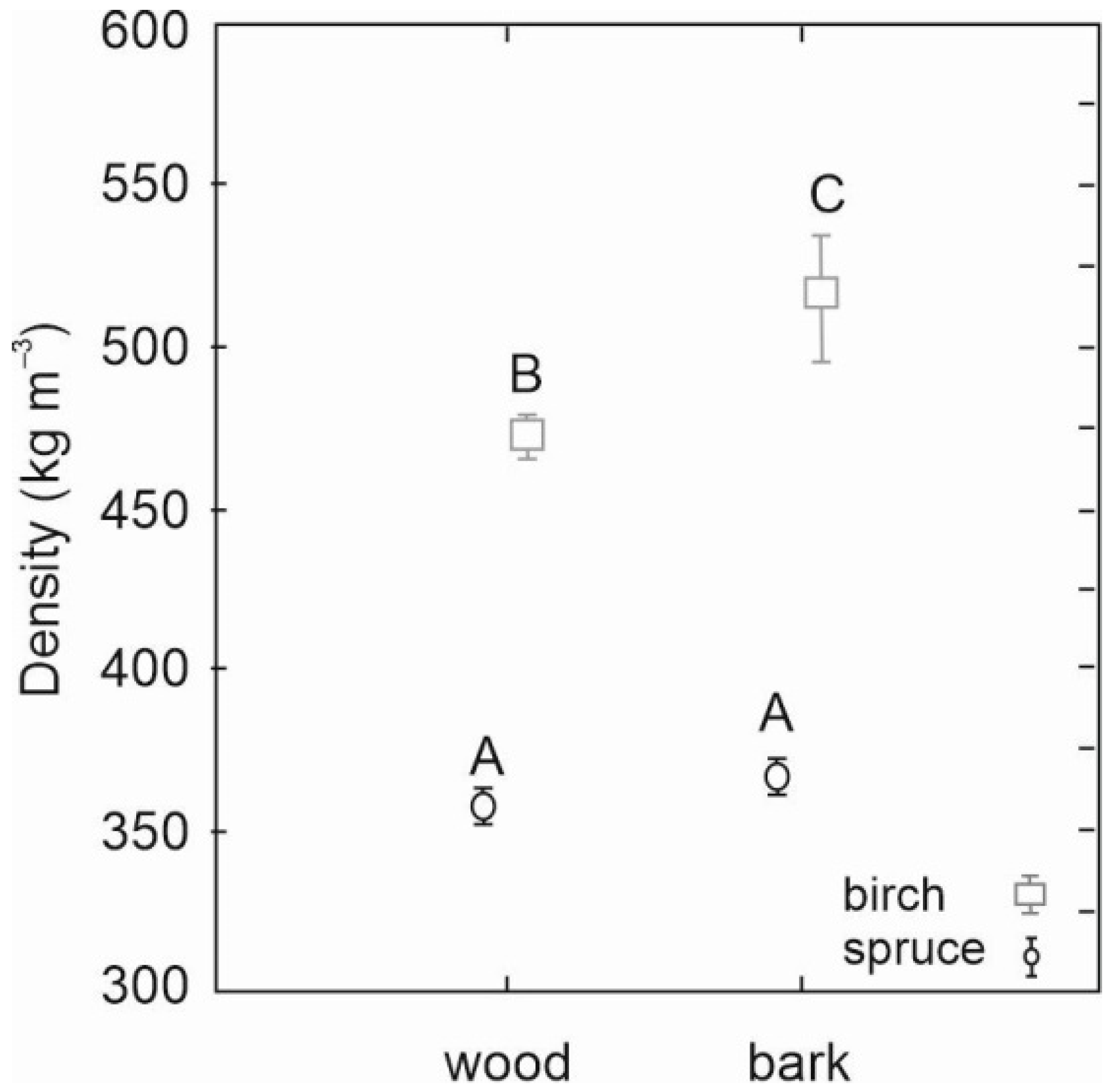
As for density, significantly higher density values of both stem wood and stem bark were revealed for birch than spruce (
Figure 4). While spruce wood and bark densities were similar (360 versus 370 kg m
−3), birch wood density was significantly lower than its bark density (470 versus 520 kg m
−3). Moreover, the results also revealed that the variability of birch bark density was greater than the variability of birch or spruce wood or bark density (
Figure 4).
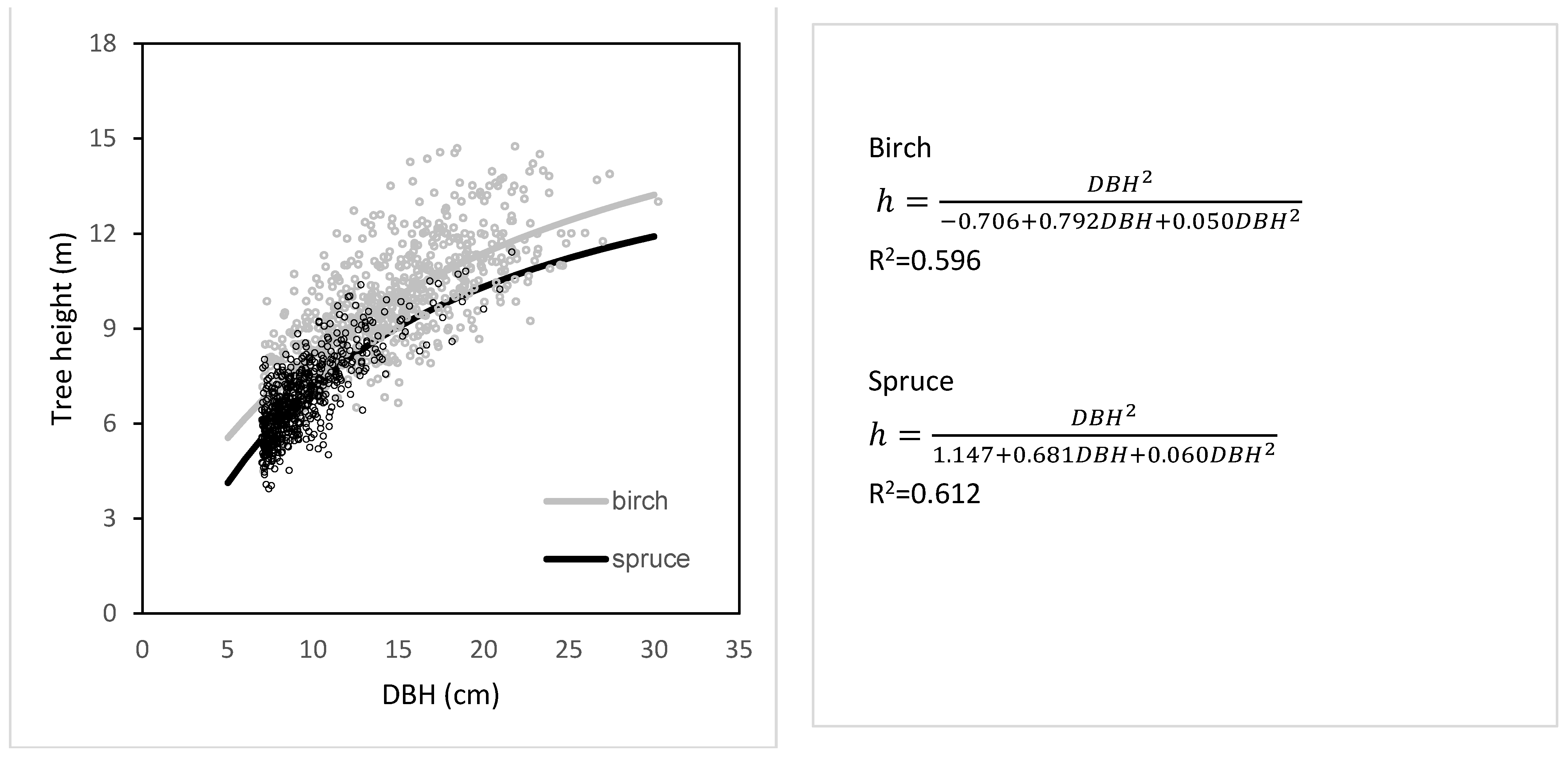
The relationships between measured tree heights and DBH of birch and spruce trees growing at transects showed differences between the two species (
Figure 5). Specifically, birch trees were by about 1.0 m higher than spruce trees with the same DBH. Even greater differences occurred between highest birches (about 15 m) and highest spruces (approx. 10 m). The data also showed that, while DBH of the thickest birches was about 28 cm, the greatest measured DBH of spruce was only 23 cm.
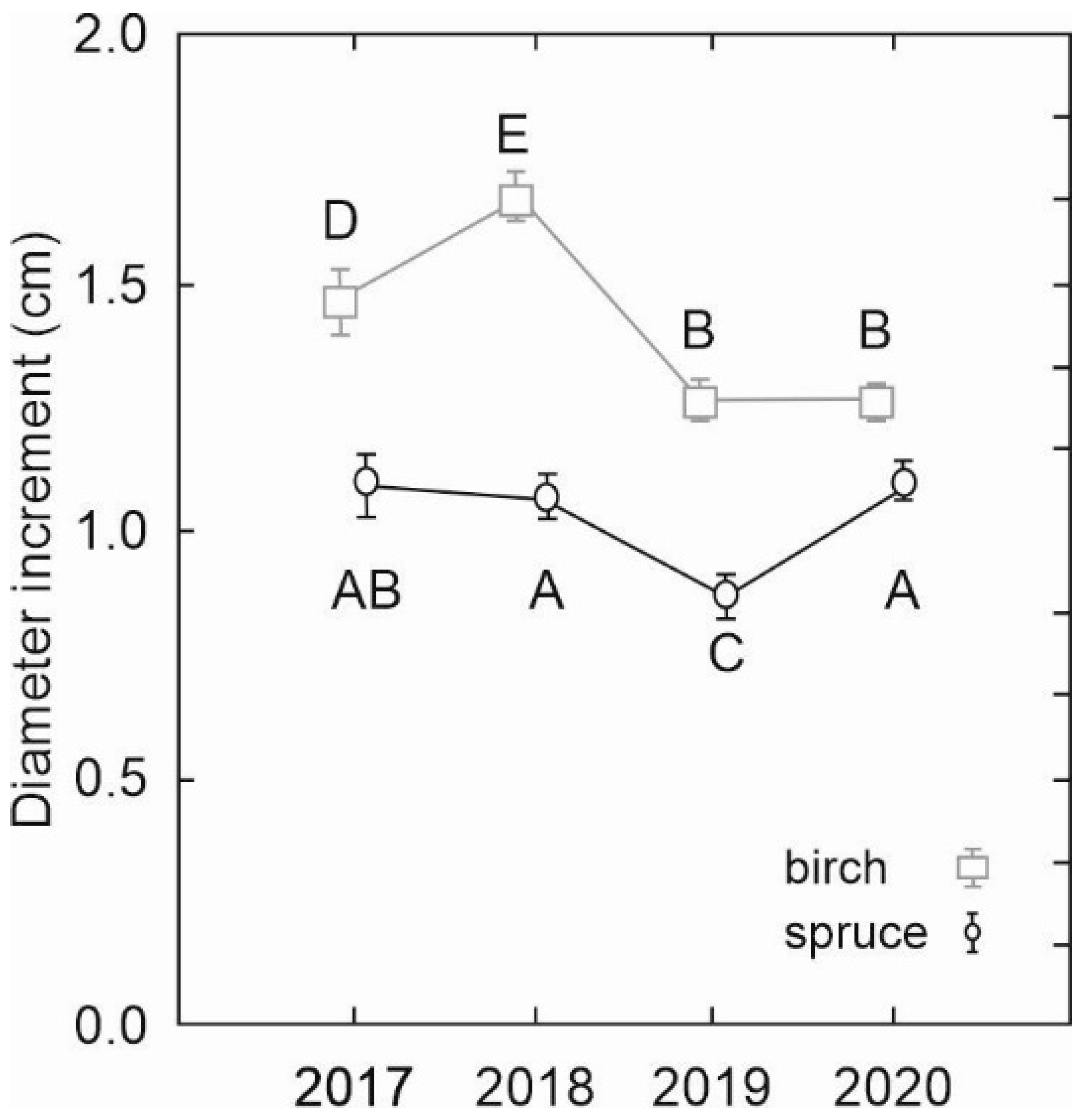
The analysis of mean annual diameter increments derived from consecutive DBH measurements revealed significantly faster diameter growth of birch trees than spruce (
Figure 6). The increments differed significantly not only between tree species but also between some years. While in the case of spruce, only one mean annual diameter increment in the year 2019 was significantly lower than in other years, the differences between the annual increments of birch were more pronounced, except for similar increments in the last two years (2019 and 2020).
Moreover, inter-specific comparisons of annual production of aboveground biomass and of individual tree components were performed (
Figure 7). The comparisons considered real mean values of diameter increment for each species derived as averages of four mean annual diameter increments, i.e., 1.4 cm and 1.0 for birch and spruce, respectively. The results indicated that 14-year-old birch trees produced annually slightly more aboveground biomass than the spruce trees of the same age with the same DBH (
Figure 7a). The opposite situation was found for foliage production, especially for trees with greater DBH (
Figure 7b). Birch produced annually more branch biomass than spruce (
Figure 7c), as well as more stem bark (
Figure 7d), stem wood (
Figure 7e) and, hence, also more stem under bark biomass (
Figure 7f). The interspecific differences in the production of tree component biomass increased with DBH (
Figure 7).
To present an example of tree component biomass stock and production (
Table 7 and
Table 8) we selected DBH approximately in the middle of the interval of sampled trees (i.e., 10 cm). Aboveground biomass stocks of both species in the current (x) year and the previous (x − 1) year were very similar. On the other hand, annual aboveground biomass production of birch was by 33.6% greater than of spruce. The greatest inter-species differences in production were found for foliage (in favor of spruce) and stem bark and wood (in favor of birch, see
Table 7 and
Table 8). In addition, we also revealed the difference in the annual production of litter. While the annual foliage litter production of a birch tree was 2.23 kg year
−1, that of spruce was 1.53 kg year
−1 (or 1.69 kg year
−1 if applying the relationship derived by Pajtík et al. [
26] and Konôpka [
27]).
Finally, we focused on modeling the contribution of stem (both over and under bark) biomass to aboveground biomass production (
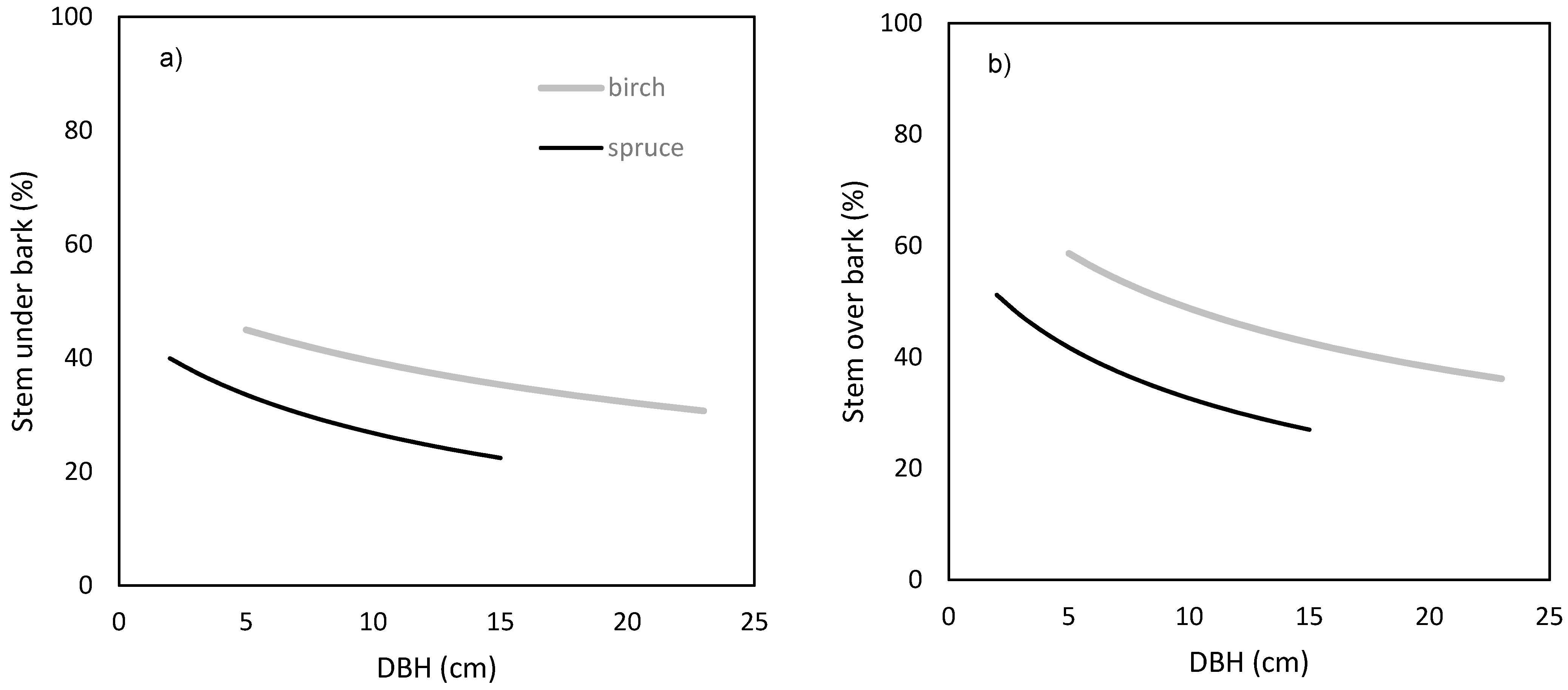
Figure 8) with respect to tree DBH. The results showed that the proportion of stem production on the annual aboveground biomass production had a decreasing trend with the increasing DBH. Moreover, birch trees invested a higher share of aboveground biomass production to stem than spruce ones.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}