
Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland



Abstract
:1. Introduction
2. Materials and Methods
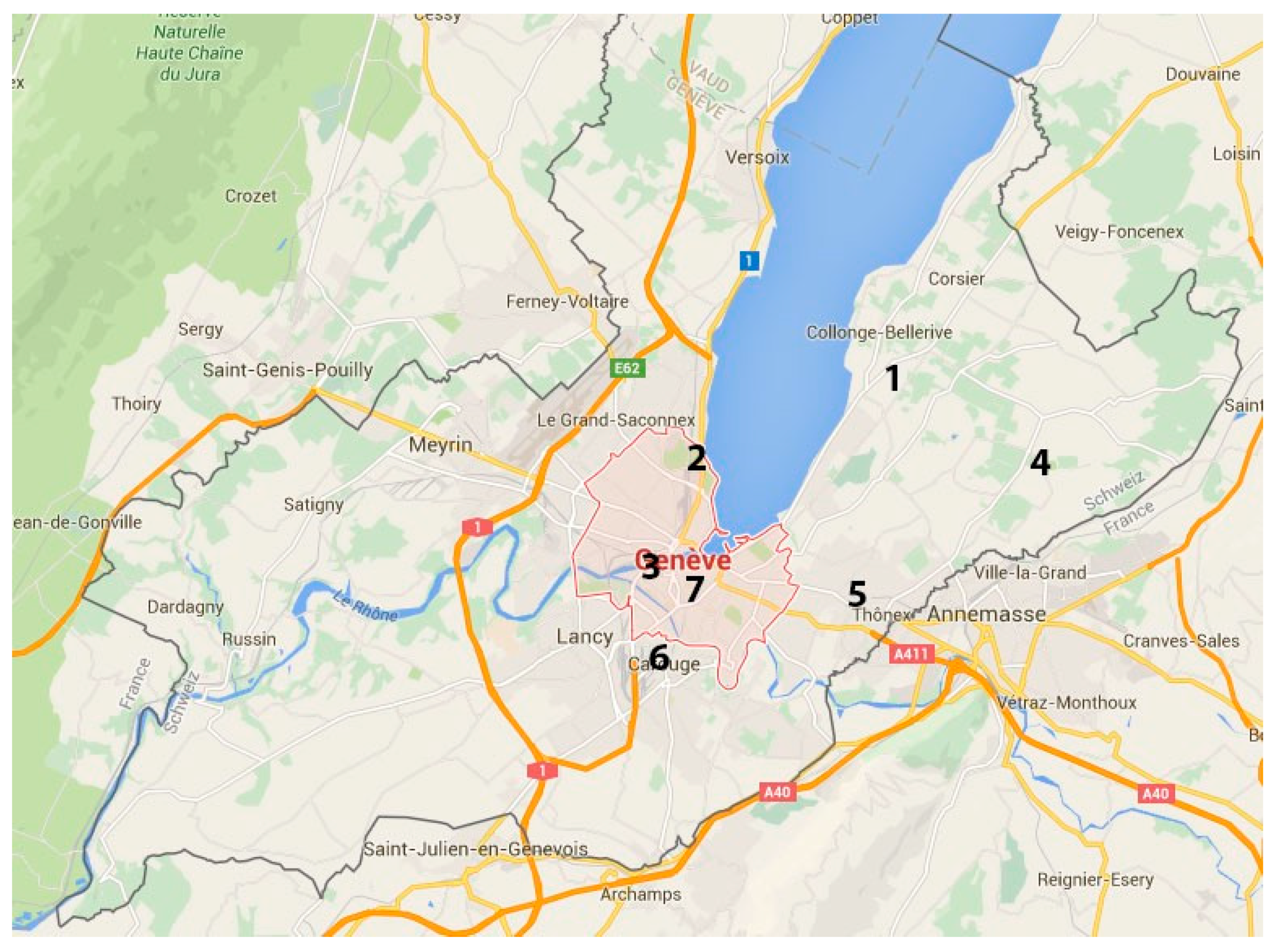
2.1. Sampled Trees
2.2. Isolation of Microorganisms
2.3. Genetic Identification
2.4. Production of Pycnidia and Measurement of Conidia Dimensions
2.5. Pathogenicity Testing According Henle-Koch’s Postulates
3. Results
3.1. Isolation and Genetic Identification
3.2. Pycnidia Production and Conidia Dimensions
3.3. Pathogenicity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesati, V.; De Notaris, G. Schema di classificazione degle sferiacei italici aschigeri piu’ o meno appartenenti al genere Sphaeria nell’antico significato attribuitoglide Persono. Comm. Soc. Critt. Ital. 1863, 1, 177–420. [Google Scholar]
- Bega, R.V. Diseases of Pacific Coast Conifers. Agriculture Handbook No 521; Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1978; p. 75. [Google Scholar]
- Worrall, J.J.; Correll, J.C.; McCain, A.H. Pathogenicity and teleomorph-anamorph connection of Botryosphaeria dothidea on Sequoiadendron giganteum and Sequoia sempervirens. Plant Dis. 1986, 70, 757–759. [Google Scholar] [CrossRef]
- Wood, D.L. Pests of the Native California Conifers. In California Natural History Guides; University of California Press: Oakland, CA, USA, 2003. [Google Scholar]
- Kehr, R. Triebschäden an Mammutbaum (Sequoiadendron giganteum) durch Botryosphaeria dothidea auch in Deutschland nachgewiesen. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 2004, 56, 37–43. [Google Scholar]
- Cech, T.; Tomiczek, C. Botryosphaeria-Krebs (Botryosphaeria dothidea) des Mammutbaumes nimmt in Ostösterreich zu. Forstsch. Aktuell 2013, 57/58, 37–40. [Google Scholar]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea, a latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef]
- Pennycook, S.R.; Samuels, G.J. Botryosphaeria and Fusicoccum species associated with ripe fruit rot of Actinidia deliciosa (kiwifruit) in New Zealand. Mycotaxon 1985, 24, 445–458. [Google Scholar]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Tsopelas, B.; Slippers, B.; Wingfield, M.J.; Gonou-Zagou, Z. Infection of Sequoiadendron giganteum in Greece by the fungus Neofusicoccum parvum. Phytopathol. Mediterr. 2009, 48, 311. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants, diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–108. [Google Scholar] [CrossRef]
- Golzar, H.; Burgess, T.I. Neofusicoccum parvum, a causal agent associated with cankers and decline of Norfolk Island pine in Australia. Austral. Plant Pathol. 2011, 40, 484–489. [Google Scholar] [CrossRef]
- Park, S.; Kim, S.-H.; Back, C.-G.; Lee, S.-Y.; Kang, I.-K.; Jung, H.-Y. First Report of Botryosphaeria parva causing stem blight on Rubus crataegifolius in Korea. Res. Plant Dis. 2016, 22, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Massonnet, M.; Figueroa-Balderas, R.; Galarneau, E.R.A.; Miki, S.; Lawrence, D.P.; Sun, Q.; Wallis, C.M.; Baumgartner, K.; Cantu, D. Neofusicoccum parvum colonization of the grapevine woody stem triggers asynchronous host responses at the site of infection and in the leaves. Front. Plant Sci. 2017, 8, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.; Raimondo, M.; Cibelli, F.; Phillips, A.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum; associated with a decline of olives in Southern Italy. Phytopathol. Mediterr. 2013, 52, 517–527. [Google Scholar] [CrossRef]
- Lefort, F.; Douglas, G.C. An efficient micro-method of DNA isolation from mature leaves of four hardwood tree species Acer, Fraxinus, Prunus and Quercus. Ann. For. Sci. 1999, 56, 259–263. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.; Fourie, P.H.; Halleen, F. DNA phylogeny, morphology and pathogenicity of Botryosphaeria species on grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef]
- Henle, J. Pathologische Untersuchungen—Von den Miasmen und Contagien und von den Misamatisch-Contagiösen Krankheiten; Verlag von August Hirschwald: Berlin, Germany, 1840. [Google Scholar]
- Koch, R. Die Aetiologie der Tuberculose. Mitt. Kaiser. Gesundh. 1884, 2, 1–88. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Meier, F.; Engesser, R.; Forster, B.; Odermatt, O.; Angst, A. Protection des Forêts—Vue D’ensemble 2009; Institut fédéral de Recherches sur la Forêt, la Neige et le Paysage WSL: Birmensdorf, Switzerland, 2010. [Google Scholar]
- Lehoczky, J. Black dead-arm disease of grapevine caused by Botryosphaeria stevensii infection. Acta Phytopathol. Acad. Sci. Hung. 1974, 9, 319–327. [Google Scholar]
- Pelleteret, P.; Crovadore, J.; Cochard, B.; Pasche, S.; Bovigny, P.-Y.; Chablais, R.; Lefort, F. Urban London plane tree dieback linked to fungi in the Botryosphaeriaceae. Urban Urban Green 2017, 22, 74–83. [Google Scholar] [CrossRef]
- Haleem, R.A.; Abdullah, S.K.; Jubrael, J.M.S. Identification and pathogenicity of Botryosphaeria parva associated with grapevine decline in Kurdistan region—Iraq. Acta Agrobot. 2012, 65, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Grossenbacher, J.G.; Duggar, B.M. A contribution to the life history, parasitism, and biology of Botryosphaeria ribis. N. Y. Agric. Exp. Stn. 1911, 18, 114–188. [Google Scholar]
- Phillips, A.J.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.; Stanosz, G. Molecular and morphological differentiation of Botryosphaeria dothidea (anamorph Fusicoccum aesculi) from some other fungi with Fusicoccum anamorphs. Mycologia 2001, 93, 505–515. [Google Scholar] [CrossRef]
- Smith, D.R.; Michailides, T.J.; Stanosz, G.R. Differentiation of a Fusicoccum species causing panicle and shoot blight on California pistachio trees from Botryosphaeria dothidea. Plant Dis. 2001, 85, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Michailides, T.J. Characterization of Botryosphaeria dothidea isolates collected from pistachio and other plant hosts in California. Phytopathology 2002, 92, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.; Crous, P.W.; Taylor, J.E.; Kang, J.; Pascoe, I.; Wingfield, M.J. An overview of the taxonomic history of Botryosphaeria and a re-evaluation of its anamorphs based on morphology and ITS rDNA phylogeny. Stud. Mycol. 2000, 45, 129–140. [Google Scholar]
- Ogata, T.; Sano, T.; Harada, Y. Botryosphaeria spp. isolated from apple and several deciduous fruit trees are divided into three groups based on the production of warts on twigs, size of conidia, and nucleotide sequences of nuclear ribosomal DNA ITS regions. Mycoscience 2000, 41, 331–337. [Google Scholar] [CrossRef]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Multiple gene genealogies and phenotypic data reveal cryptic species of the Botryosphaeriaceae: A case study on the Neofusicoccum parvum/N. ribis complex. Mol. Phylogenet. Evol. 2009, 51, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Molecular and phenotypic characterisation of three phylogenetic species discovered within the Neofusicoccum parvum/N. ribis complex. Mycologia 2009, 101, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Inderbitzin, P.; Bostock, R.M.; Trouillas, F.P.; Michailides, T.J. A six-locus phylogeny reveals high species diversity in Botryosphaeriaceae from California almond. Mycologia 2010, 102, 1350–1368. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Tidwell, T.E.; Blomquist, C.L.; Peek, K.S. First report of Neofusicoccum nonquaesitum causing branch cankers on giant sequoia (Sequoiadendron giganteum) in North America. Plant Dis. 2012, 96, 905. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Multiple gene sequences delimit Botryosphaeria australis sp. nov. from B. lutea. Mycologia 2004, 96, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, N.L.; Das, A.J.; Ampersee, N.J.; Cahill, K.G.; Caprio, A.C.; Sanders, J.E.; Williams, A.P. Patterns and correlates of giant sequoia foliage dieback during California’s 2012–2016 hotter drought. For. Ecol. Manag. 2018, 419–420, 268–278. [Google Scholar] [CrossRef]
- DeSilva, R.; Dodd, R.S. Association of genetic and climatic variability in giant sequoia, Sequoiadendron giganteum, reveals signatures of local adaptation along moisture-related gradients. Ecol. Evol. 2020, 10, 10619–10632. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree | Localisation | Disease Severity 1 | Sampled Material | Isolate Nb | Pathogen Species | GenBank Accession | Isolate Accession |
|---|---|---|---|---|---|---|---|
| No1 | Vésenaz | ++ | twig | C | N. parvum | NA | NA |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R1 | N. parvum | MH500829 | UASWS1756 |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R2 | N. parvum | MH500830 | UASWS1757 |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R3 | N. parvum | MH500831 | UASWS1758 |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R4 | N. parvum | MH500832 | UASWS1759 |
| No3 | Geneva (Parc Barton) | +++ | twig | B1 | N. parvum | MH500826 | UASWS1753 |
| No3 | Geneva (Parc Barton) | +++ | twig | B2 | N. parvum | MH500827 | UASWS1754 |
| No3 | Geneva (Parc Barton) | +++ | twig | B3 | N. parvum | MH500828 | UASWS1755 |
| No4 | Lullier | + | twig | A1 | Fusicoccum aesculi | MH500836 | UASWS1763 |
| No5 | Lullier | +/− | twig | B1 Lullier | Sordaria sp. | MH500837 | UASWS1764 |
| No6 | Lullier | + | twig | C1 | N. parvum | MH500838 | UASWS1765 |
| No6 | Lullier | bark | D1 | Diplodia mutila | MH500839 | UASWS1766 | |
| No7 | Chêne-Bourg | ++ | stroma on trunk | E1 | N. parvum | MH500840 | UASWS1767 |
| No8 | Carouge | + | twig | FV2 | Diplodia seriata | MH500841 | UASWS1768 |
| No8 | Carouge | + | twig | 1.1. | N. parvum | MH500822 | UASWS1749 |
| No8 | Carouge | + | twig | 2.1 | Diplodia mutila | MH500823 | UASWS1750 |
| No8 | Carouge | + | twig | 2.2 | Sordaria sp. | MH500824 | UASWS1751 |
| No8 | Carouge | + | twig | 3.1 | Phomopsis sp. | MH500825 | UASWS1752 |
| No9 | Geneva (Parc des Bastions) | ++ | twig | Bastions 1.3 | N. parvum | MH500833 | UASWS1760 |
| No9 | Geneva (Parc des Bastions) | ++ | twig | Bastions 3.1 | Fusicoccum aesculi | MH500834 | UASWS1761 |
| No9 | Geneva (Parc des Bastions) | ++ | twig | Bastions 4.1 | Fusicoccum aesculi | MH500835 | UASWS1762 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haenzi, M.; Cochard, B.; Chablais, R.; Crovadore, J.; Lefort, F. Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland. Forests 2021, 12, 434. https://doi.org/10.3390/f12040434
Haenzi M, Cochard B, Chablais R, Crovadore J, Lefort F. Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland. Forests. 2021; 12(4):434. https://doi.org/10.3390/f12040434
Chicago/Turabian StyleHaenzi, Martine, Bastien Cochard, Romain Chablais, Julien Crovadore, and François Lefort. 2021. "Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland" Forests 12, no. 4: 434. https://doi.org/10.3390/f12040434
APA StyleHaenzi, M., Cochard, B., Chablais, R., Crovadore, J., & Lefort, F. (2021). Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland. Forests, 12(4), 434. https://doi.org/10.3390/f12040434