
Botryosphaeria Dothidea and Neofusicoccum Yunnanense Causing Canker and Die-Back of Sequoiadendron Giganteum in Croatia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Fungal Isolation, and Morphological Characterization
2.2. DNA Extraction, PCR Amplification and Phylogenetic Analyses
2.3. Path ogenicity Test and Statistical Analysis
3. Results
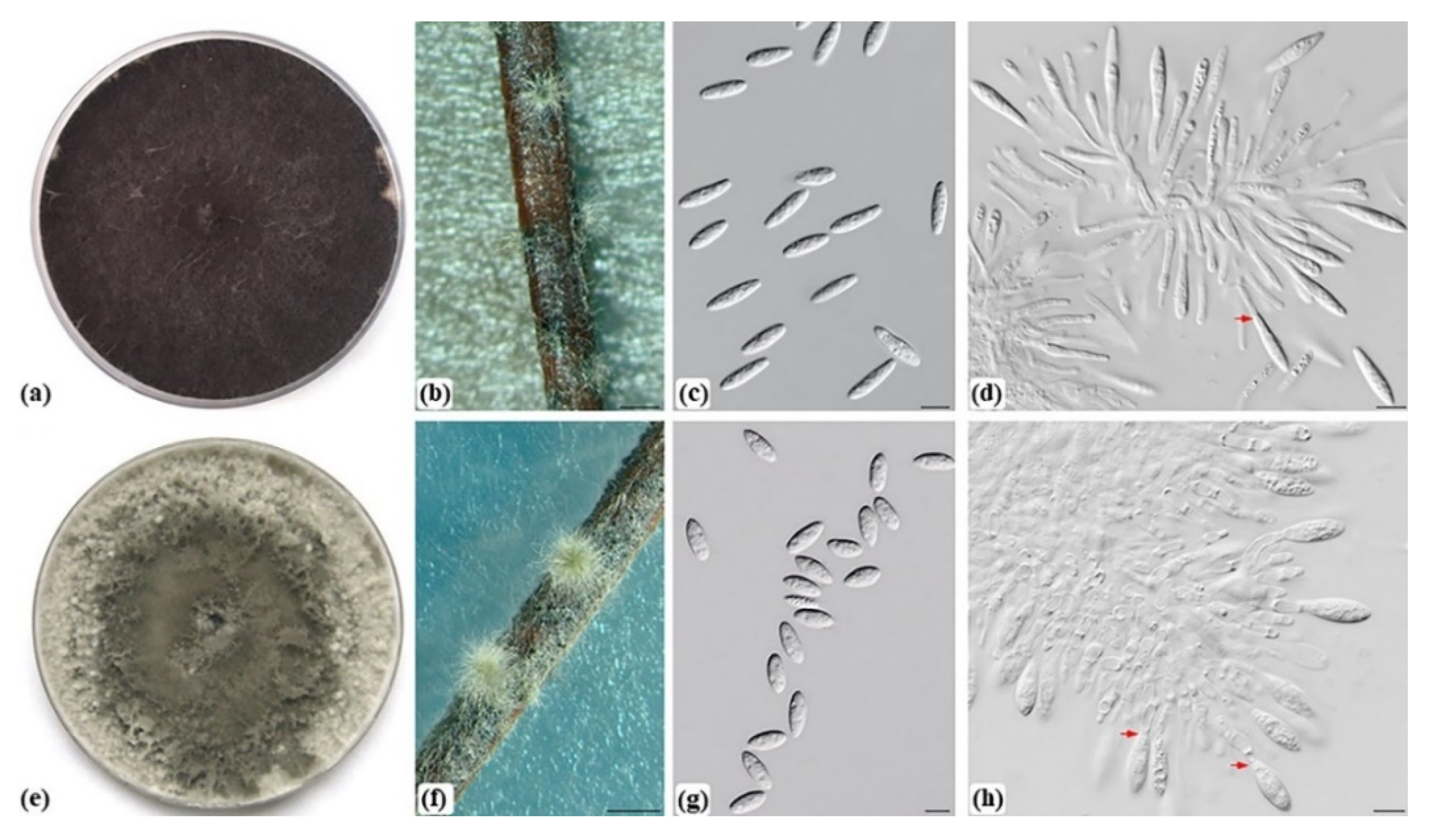
3.1. Fungal Isolation, and Morphological Characterization
3.2. Phylogenetic Analyses
3.3. Pathogenicity Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harvey, H.T.; Shellhammer, H.S.; Stecker, R.E. Giant Sequoia Ecology: Fire and Reproduction; Scientific Monograph Series No. 12, U.S. Department of the Interior, National Park Service: Washington, DC, USA, 1980; p. 221. [Google Scholar]
- Paz-Kagan, T.; Vaughn, N.R.; Martin, R.E.; Brodrick, P.G.; Stephenson, N.L.; Das, A.J.; Nydick, K.R.; Asner, G.P. Landscape-scale variation in canopy water content of giant sequoias during drought. For. Ecol. Manag. 2018, 419, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Dodd, R.S.; DeSilva, R. Long-term demographic decline and late glacial divergence in a Californian paleoendemic: Sequoiadendron giganteum (giant sequoia). Ecol. Evol. 2016, 6, 3342–3355. [Google Scholar] [CrossRef] [Green Version]
- IUCN Red List of Threatened Species (Ver. 3.1). Available online: http://www.iucnredlist.org (accessed on 23 March 2021).
- Nydick, K.R.; Stephenson, N.L.; Ambrose, A.R.; Asner, G.P.; Baxter, W.B.; Das, A.J.; Dawson, T.; Martin, R.E.; Paz-Kagan, T. Leaf to landscape responses of giant sequoia to hotter drought: An introduction and synthesis for the special section. For. Ecol. Manag. 2018, 419–420, 249–256. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Tidwell, T.E.; Blomquist, C.L.; Peek, K.S. First report of Neofusicoccum nonquaesitum causing branch cankers on giant sequoia (Sequoiadendron giganteum) in North America. Plant Dis. 2012, 96, 905. [Google Scholar] [CrossRef] [PubMed]
- Morelet, M.; Andréoli, C.; Chandelier, P.; Ménard, J.-E. Botryosphaeria dothidea, agent de chancre sur Sequoiadendron giganteum. Rev. For. Franc. 1993, 45, 37–42. (In French) [Google Scholar] [CrossRef] [Green Version]
- Kehr, R. Branch dieback of giant sequoia (Sequoiadendron giganteum) in Germany caused by Botryosphaeria dothidea. Nachr. Des. Dtsch. Pflanzenschutzd. 2004, 56, 37–43. (In German) [Google Scholar]
- Cech, T.L.; Christian, T. Increasing prevalence of Botryosphaeria-canker (Botryosphaeria dothidea) of giant sequoia in Eastern Austria. Forstsch. Aktuell 2013, 57, 37–40, (In German with an English abstract). [Google Scholar]
- Vajna, L.; Schwarczinger, I. Fungi causing branch dieback of giant sequoia (Sequoiadendron giganteum (Lindl.) Buchholcz) in Hungary. Növényvédelem 1998, 34, 133–136, (In Hungarian with an English abstract). [Google Scholar]
- Tsopelas, P.; Slippers, B.; Wingfield, M.J.; Zagkou, Z.G. Infection of Sequoiadendron giganteum in Greece by the fungus Neofusicoccum parvum. In Proceedings of the 14th National Congress of Hellenic Phytopathological Society, Argolida, Greece, 7–10 October 2008. [Google Scholar]
- Georgieva, M. First record of the pathogen Botryosphaeria dothidea associated with Sequoiadendron giganteum in Bulgaria. Silva Balcanica 2016, 17, 57–62. [Google Scholar]
- Zlatković, M.; Keča, N.; Wingfield, M.J.; Jami, F.; Slippers, B. Botryosphaeriaceae associated with the die-back of ornamental trees in the Western Balkans. Antonie Van Leeuwenhoek 2016, 109, 543–564. [Google Scholar] [CrossRef] [Green Version]
- Zlatković, M.; Wingfield, M.J.; Jami, F.; Slippers, B. Host specificity of co-infecting Botryosphaeriaceae on ornamental and forest trees in the Western Balkans. For. Path. 2018, 48, e12410. [Google Scholar] [CrossRef] [Green Version]
- Zlatković, M. Botryosphaeriaceae species associated with canker and die-back disease of conifers in urban environments in Serbia. Topola/Poplar 2017, 199, 55–75, (In Serbian with an English summary). [Google Scholar]
- Haenzi, M.; Cochard, B.; Chablais, R.; Crovadore, J.; Lefort, F. Neofusicoccum parvum, a new agent of sequoia canker and dieback identified in Geneva, Switzerland. Forests 2021, 12, 434. [Google Scholar] [CrossRef]
- Pavlic-Zupanc, D.; Maleme, H.M.; Piškur, B.; Wingfield, B.D.; Wingfield, M.J.; Slippers, B. Diversity, phylogeny and pathogenicity of Botryosphaeriaceae on non-native Eucalyptus grown in an urban environment: A case study. Urban For. Urban Green. 2017, 26, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Moricca, S.; Uccello, A.; Ginetti, B.; Ragazzi, A. First report of Neofusicoccum parvum associated with bark canker and dieback of Acer pseudoplatanus and Quercus robur in Italy. Plant Dis. 2012, 96, 1699. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.A.; Gelfand, D.H.; Sninsky, J.J.; White, T.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications Innis MA; Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Darren, K.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nature Protocols 2008, 3, 1452. [Google Scholar] [CrossRef]
- Zlatković, M.; Tenorio-Baigorria, I.; Lakatos, T.; Tóth, T.; Koltay, A.; Pap, P.; Marković, M.; Orlović, S. Bacterial canker disease on Populus × euramericana caused by Lonsdalea populi in Serbia. Forests 2020, 11, 1080. [Google Scholar] [CrossRef]
- Taylor, D.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Zlatković, M.; Keča, N.; Wingfield, M.W.; Jami, F.; Slippers, B. New and unexpected host associations for Diplodia sapinea in the Western Balkans. For. Path. 2017, 47, e12328. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Jurković, M.; Jurković-Bevilacqua, B. Golemi mamutovac- Sequoiadendron giganteum (Lindl.) Buchh. u zagrebačkim parkovima. Sumar. List 1996, 1–2, 47–53. (In Croatian) [Google Scholar]
- Worral, J.J.; Correll, J.C.; McCain, A.H. Pathogenicity and teleomorph-anamorph connection of Botryosphaeria dothidea on Sequoiadendron giganteum and Sequoia sempervirens. Plant Dis. 1986, 70, 757–759. [Google Scholar] [CrossRef]
- Kaliterna, J.; Miličević, T. In Proceedings of the Incidence and Distribution of Fungi from Diaporthaceae and Botryosphaeriaceae on Grapevine in Croatia. In Proceedings of the 11th Conference of the European Foundation for Plant Pathology: Healthy Plants—Healthy People, Krakov, Poland, 8–13 September 2014. [Google Scholar]
- Inderbitzin, P.; Bostock, R.M.; Trouillas, F.P.; Michailides, T.J. A six locus phylogeny reveals high species diversity in Botryosphaeriaceae from California almond. Mycologia 2010, 102, 1350–1368. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Multiple gene sequences delimit Botryosphaeria australis sp. nov. from B. lutea. Mycologia 2004, 96, 1030–1041. [Google Scholar] [CrossRef]
- Guoqing, L.; Slippers, B.; Wingfield, M.J.; Shuaifei. C. Variation in Botryosphaeriaceae from Eucalyptus plantations in YunNan Province in southwestern China across a climatic gradient. IMA Fungus 2020, 11, 1–49. [Google Scholar] [CrossRef]
- Aćimović, S.; Rooney-Latham, S.; Albu, S.; Grosman, D.M.; Doccola, J.J. Characterization and pathogenicity of Botryosphaeriaceae fungi associated with declining urban stands of coast redwood in California. Plant Dis. 2018, 102, 1950–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Path. 2008, 57, 948–956. [Google Scholar] [CrossRef]
- State Hydrometeorological Institute. Available online: www.meto.hr (accessed on 13 April 2021).
- Eschen, R.; O’Hanlon, R.; Santini, A.; Vannini, A.; Roques, A.; Kirichenko, N.; Kenis, M. Safeguarding global plant health: The rise of sentinels. J. Pest Sci. 2019, 92, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Orlović, S.; Zlatković, M.; (Institute of Lowland Forestry and Environment (ILFE), University of Novi Sad, Novi Sad, Serbia). Personal communication, 2021.
- Aćimović, S.G.; Martin, D.K.H.; Turcotte, R.M.; Meredith, C.L.; Munck, I.A. Choosing an adequate pesticide delivery system for managing pathogens with difficult biologies: Case studies on Diplodia corticola, Venturia inaequalis and Erwinia amylovora. Plant Dis. Curr. Threat. Manag. Trends 2019. [Google Scholar] [CrossRef] [Green Version]
- Karen, J.; Paine, T.D. Consumer preferences and willingness to pay for biological control in the urban landscape. Biol. Cont. 2004, 30, 312–322. [Google Scholar] [CrossRef]
- Karličić, V.; Jovičić-Petrović, J.; Marojević, V.; Zlatković, M.; Orlović, S.; Raičević, V. Potential of Trichoderma spp. and Pinus sylvestris bark extracts as biocontrol agents against fungal pathogens residing in the Botryosphaeriales. Environ. Sci. Proc. 2020, 3, 99. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Abou-Mansour, E.; Courteaux, B.; Rabenoelina, F.; Clément, C.; Fontaine, F.; Aziz, A. Bacillus subtilis PTA-271 counteracts Botryosphaeria dieback in grapevine, triggering immune responses and detoxification of fungal phytotoxins. Front. Plant Sci. 2019, 10, 25. [Google Scholar] [CrossRef]
- Zhai, L.; Xiang, J.; Zhang, M.; Fu, M.; Yang, Z.; Hong, N.; Wang, G. Characterization of a novel double-stranded RNA mycovirus conferring hypovirulence from the phytopathogenic fungus Botryosphaeria dothidea. Virology 2016, 493, 75–85. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate No. 1,2 | Identity | Host | Location | Collector | GenBank Accession No. 3,4 | ||
|---|---|---|---|---|---|---|---|
| ITS | tef 1-α | TUB2 | |||||
| ILFE 2 | Botryosphaeria dothidea | Sequoiadendron giganteum | Zagreb, Croatia | M. Kovač | MW033525 | MW051688 | MW051690 |
| ILFE 3 | B. dothidea | S. giganteum | Zagreb, Croatia | M. Kovač | MW033526 | MW051689 | MW051691 |
| CMW 39308 | B. dothidea | S. giganteum | Valjevo, Serbia | N. Keča | KF575008 | KF575040 | KF575104 |
| CBS 110302 | B. dothidea | Vitis vinifera | Portugal | A.J.L. Phillips | AY259092 | AY573218 | EU673106 |
| CMW 8000 | B. dothidea | Prunus sp. | Crocifisso, Switzerland | B. Slippers | AY236949 | AY236898 | AY236927 |
| CERC 2947 | B. qingyuanensis | Eucalyptus hybrid | China | S.F.Chen & G.Q.Li | KX278001 | KX278106 | KX278210 |
| CERC 2946 | B. qingyuanensis | Eucalyptus hybrid | China | S.F.Chen & G.Q.Li | KX278000 | KX278105 | KX278209 |
| CBS 135219 | B. kuwatsukai | Malus domestica | China | C.S. Wang | KJ433388 | KJ433410 | ‒ |
| LSP 5 | B. kuwatsukai | Pyrus sp. | China | C.S. Wang | KJ433395 | KJ433417 | ‒ |
| CBS 127194 | B. fabicerciana | Eucalyptus sp. | China | M.J. Wingfield | HQ332198 | HQ332214 | KF779069 |
| CBS 127193 | B. fabicerciana | Eucalyptus sp. | China | M.J. Wingfield | HQ332197 | HQ332213 | KF779068 |
| ATCC 22927 | B. corticis | Vaccinium sp. | North Carolina, USA | R.D. Millholland | DQ299247 | EU673291 | EU673108 |
| CBS 119047 | B. corticis | V. corymbosum | USA | P.V. Oudemans | DQ299245 | EU017539 | EU673107 |
| CBS 124702 | B. scharifii | Magnifera indica | Iran | J. Abdollahzadeh & A. Javadi | JQ772019 | JQ772056 | ‒ |
| CBS 124703 | B. scharifii | M. indica | Iran | J. Abdollahzadeh | JQ772020 | JQ772057 | ‒ |
| CGMCC 3.18004 | B. ramosa | Acacia sp. | Hainan, China | Z.P. Dou & W. He | KX197073 | KX197093 | KX197100 |
| CBS 122069 | B. ramosa | E. camaldulensis | Australia | T.I. Burgess | EU144055 | EU144070 | KF766132 |
| CBS 141505 | B. agaves | Agave sp. | France | P.W. Crous | KX306750 | MT592030 | MT592463 |
| CBS 133992 | B. agaves | Agave sp. | Thailand | R. Phookamsak | JX646791 | JX646856 | JX646841 |
| CBS 116131 | Neofusicoccum arbuti | Arbutus menziesii | USA | M. Elliott | AY819720 | KF531792 | KF531793 |
| CBS 117090 | N. arbuti | A. menziesii | USA | M. Elliott | AY819724 | KF531791 | KF531794 |
| CBS 126655 | N. nonquaesitum | Umbellularia californica | USA | F.P. Trouillas | GU251163 | GU251295 | GU251823 |
| CBS 133501 | N. nonquaesitum | Persea americana | USA | A. Eskalen | MT587498 | MT592213 | MT592705 |
| CGMCC 3.18313 | N. illicii | Illicium verum | Guangxi, China | L. Wang | KY350152 | KY817758 | KY350158 |
| CGMCC 3.18311 | N. illicii | I. verum | Guangxi, China | L. Wang | KY350150 | KY817756 | KY350156 |
| CBS 123634 | N. cordaticola | Syzygium cordatum | South Africa | D. Pavlić | EU821898 | EU821868 | EU821838 |
| CBS 123635 | N. cordaticola | S. cordatum | South Africa | D. Pavlić | EU821903 | EU821873 | EU821843 |
| CMM 1285 | N. brasilliense | Mangifera indica | Brazil | M.W. Marques | JX513628 | JX513608 | KC794030 |
| CMM 1338 | N. brasilliense | M. indica | Brazil | M.W. Marques | JX513630 | JX513610 | KC794031 |
| CBS 123639 | N. kwambonambiense | S. cordatum | South Africa | D. Pavlić | EU821900 | EU821870 | EU821840 |
| CBS 123641 | N. kwambonambiense | S. cordatum | South Africa | D. Pavlić | EU821919 | EU821889 | EU821859 |
| CBS 121.26 | N. ribis | Ribes rubrum | New York, USA | N.E. Stevens | AF241177 | AY236879 | AY236908 |
| CBS 115475 | N. ribis | Ribes sp. | USA | B. Slippers & G. Hudler | AY236935 | AY236877 | AY236906 |
| CERC 3416 | N. sinoeucalypti | Eucalyptus hybrid | Guangxi, China | S.F. Chen & G.Q. Li | KX278064 | KX278169 | KX278273 |
| CERC 2005 | N. sinoeucalypti | E. urophylla × E. grandis | Guangdong, China | S.F. Chen & G.Q. Li | KX278061 | KX278166 | KX278270 |
| CBS 128008 | N. occulatum | E. grandis hybrid | Australia | T.I. Burgess | EU301030 | EU339509 | EU339472 |
| MUCC 286 | N. occulatum | E. pellita | Australia | T.I. Burgess | EU736947 | EU339511 | EU339474 |
| CSF 5721 | N. dianense | E. globulus | YunNan, China | S.F. Chen & G.Q. Li | MT028608 | MT028774 | MT028940 |
| CSF 6075 | N. dianense | E. urophylla × E. grandis | YunNan, China | S.F. Chen & G.Q. Li | MT028605 | MT028771 | MT028937 |
| CSF 6142 | N. yunnanense | E. globulus | YunNan, China | S.F. Chen & G.Q. Li | MT028667 | MT028833 | MT028999 |
| CSF 6034 | N. yunnanense | E. urophylla × E. grandis | YunNan, China | S.F. Chen & G.Q. Li | MT028672 | MT028838 | MT029004 |
| ILFE 4 | N. yunnanense | S. giganteum | Zagreb, Croatia | M. Kovač | MW085805 | MW071142 | MW071144 |
| ILFE 5 | N. yunnanense | S. giganteum | Zagreb, Croatia | M. Kovač | MW085806 | MW071143 | MW071145 |
| CMW 39327 * | N. yunnanense | S. giganteum | Belgrade, Serbia | M. Zlatković | KF729050 | KF729380 | KF729340 |
| CMW 9080 | N. parvum | Populus nigra | New Zealand | G. J. Samuels | AY236943 | AY236888 | AY236917 |
| CMW 9081 | N. parvum | P. nigra | New Zealand | G.J. Samuels | AY236943 | AY236888 | AY236917 |
| CGMCC 3.18748 | N. hongkongense | Araucaria cunninghamii | Hong Kong, China | S.F. Chen | KX278051 | KX278156 | KX278260 |
| CGMCC 3.18747 | N. hongkongense | A. cunninghamii | Hong Kong, China | S.F. Chen | KX278050 | KX278155 | KX278259 |
| CBS 117448 | Pseudofusicoccum stromaticum | Eucalyptus hybrid | Venezuela | S. Mohali | AY693974 | AY693975 | EU673094 |
| CBS 117449 | P. stromaticum | Eucalyptus hybrid | Venezuela | S. Mohali | DQ436935 | DQ436936 | EU673093 |
| Isolates | Locus | ||||||
|---|---|---|---|---|---|---|---|
| ITS | tef 1-α | TUB2 | |||||
| 48 | 147 | 290 | 294 | 322 | 386 | 404 | |
| N. parvum CMW 9081 * | T | C | A | A | T | T | T |
| N. parvum CMW 9080 | T | C | A | A | T | T | T |
| N. yunnanense CSF 6142 * | A | T | G | C | C | C | C |
| N. yunnanense CSF 6034 | A | T | G | C | C | C | C |
| CMW 39327 | A | C | G | C | C | C | C |
| ILFE 4 | A | C | G | C | C | C | C |
| ILFE 5 | A | C | G | C | C | C | C |
| B. dothidea 8000 * | T | ||||||
| B. dothidea CBS 110302 | T | ||||||
| B. dothidea CMW 39308 | C | ||||||
| ILFE 2 | C | ||||||
| ILFE 3 | C | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovač, M.; Diminić, D.; Orlović, S.; Zlatković, M. Botryosphaeria Dothidea and Neofusicoccum Yunnanense Causing Canker and Die-Back of Sequoiadendron Giganteum in Croatia. Forests 2021, 12, 695. https://doi.org/10.3390/f12060695
Kovač M, Diminić D, Orlović S, Zlatković M. Botryosphaeria Dothidea and Neofusicoccum Yunnanense Causing Canker and Die-Back of Sequoiadendron Giganteum in Croatia. Forests. 2021; 12(6):695. https://doi.org/10.3390/f12060695
Chicago/Turabian StyleKovač, Marta, Danko Diminić, Saša Orlović, and Milica Zlatković. 2021. "Botryosphaeria Dothidea and Neofusicoccum Yunnanense Causing Canker and Die-Back of Sequoiadendron Giganteum in Croatia" Forests 12, no. 6: 695. https://doi.org/10.3390/f12060695
APA StyleKovač, M., Diminić, D., Orlović, S., & Zlatković, M. (2021). Botryosphaeria Dothidea and Neofusicoccum Yunnanense Causing Canker and Die-Back of Sequoiadendron Giganteum in Croatia. Forests, 12(6), 695. https://doi.org/10.3390/f12060695