
Enhancing the Resilience of a Mediterranean Forest to Extreme Drought Events and Climate Change: Pinus—Tetraclinis Forests in Europe




Abstract
:1. Introduction
1.1. Solar Irradiation and Reproduction
1.2. Interspecific Competition
2. Materials and Methods
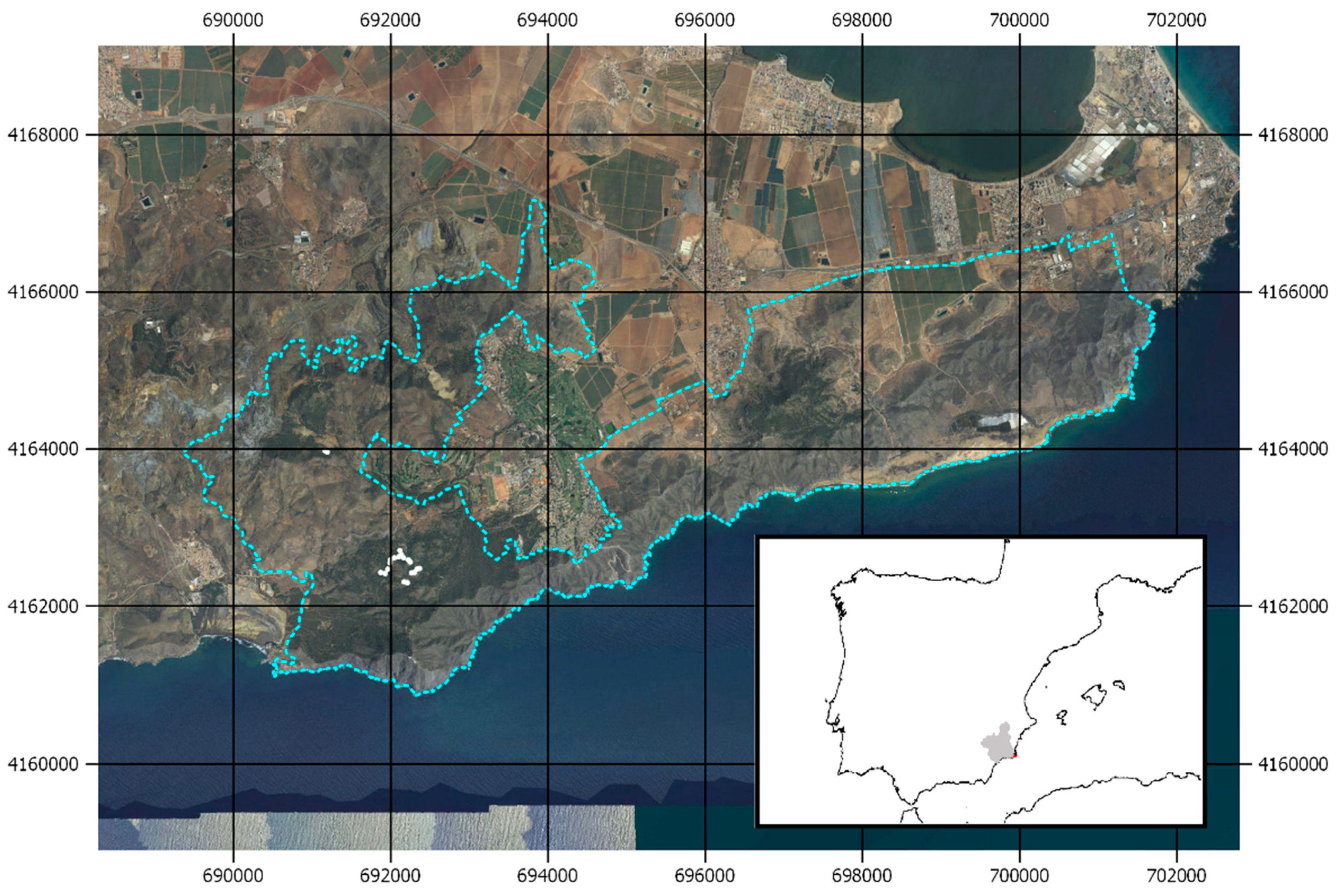
2.1. Experimental Design
2.2. Data Collection
2.3. LiDAR-Based Light Model
2.4. BACI Analysis Design
2.5. Reproductive Status Variation
2.6. Demographic Response Observed in the Experimental Plots
3. Results
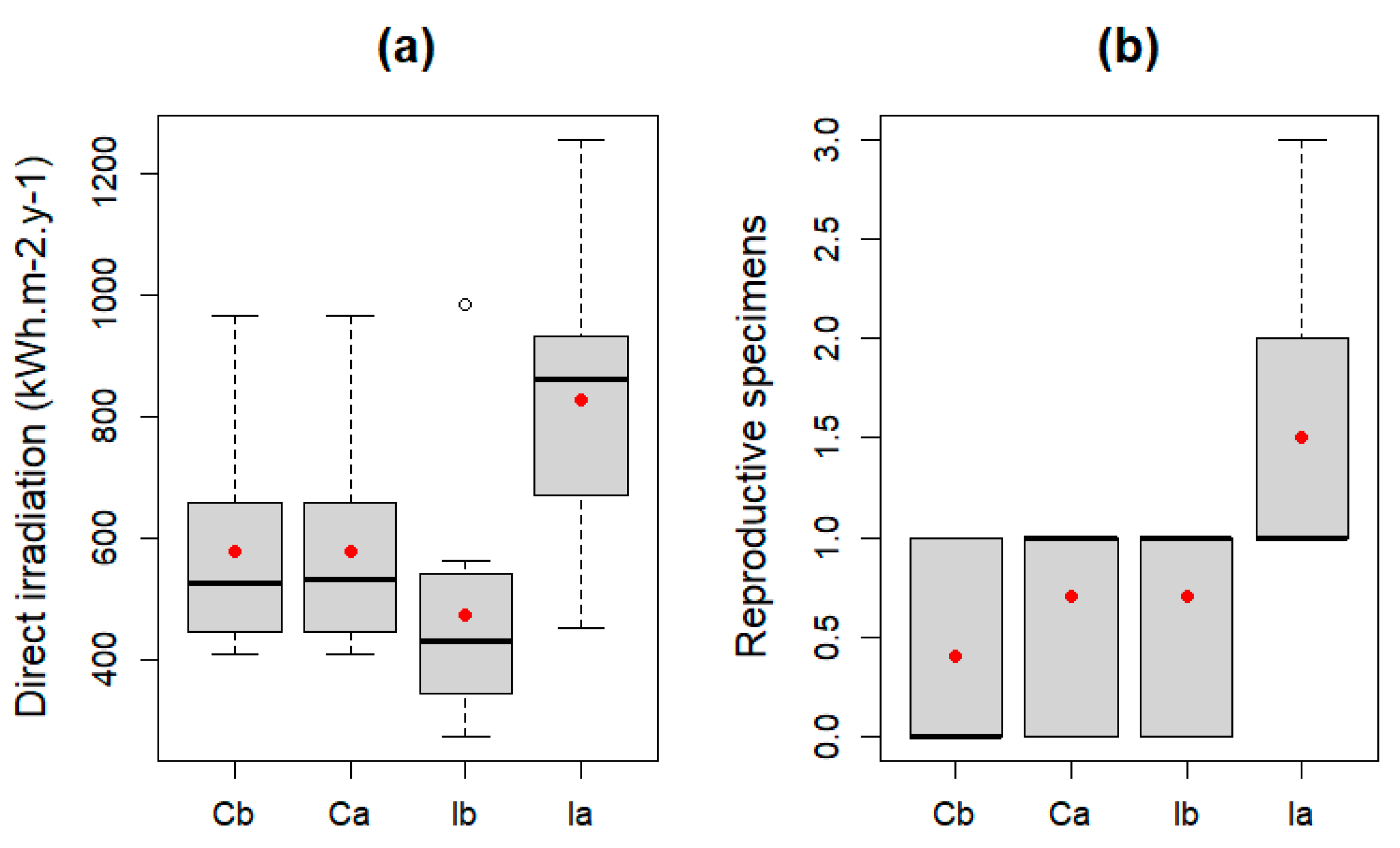
3.1. BACI Analysis
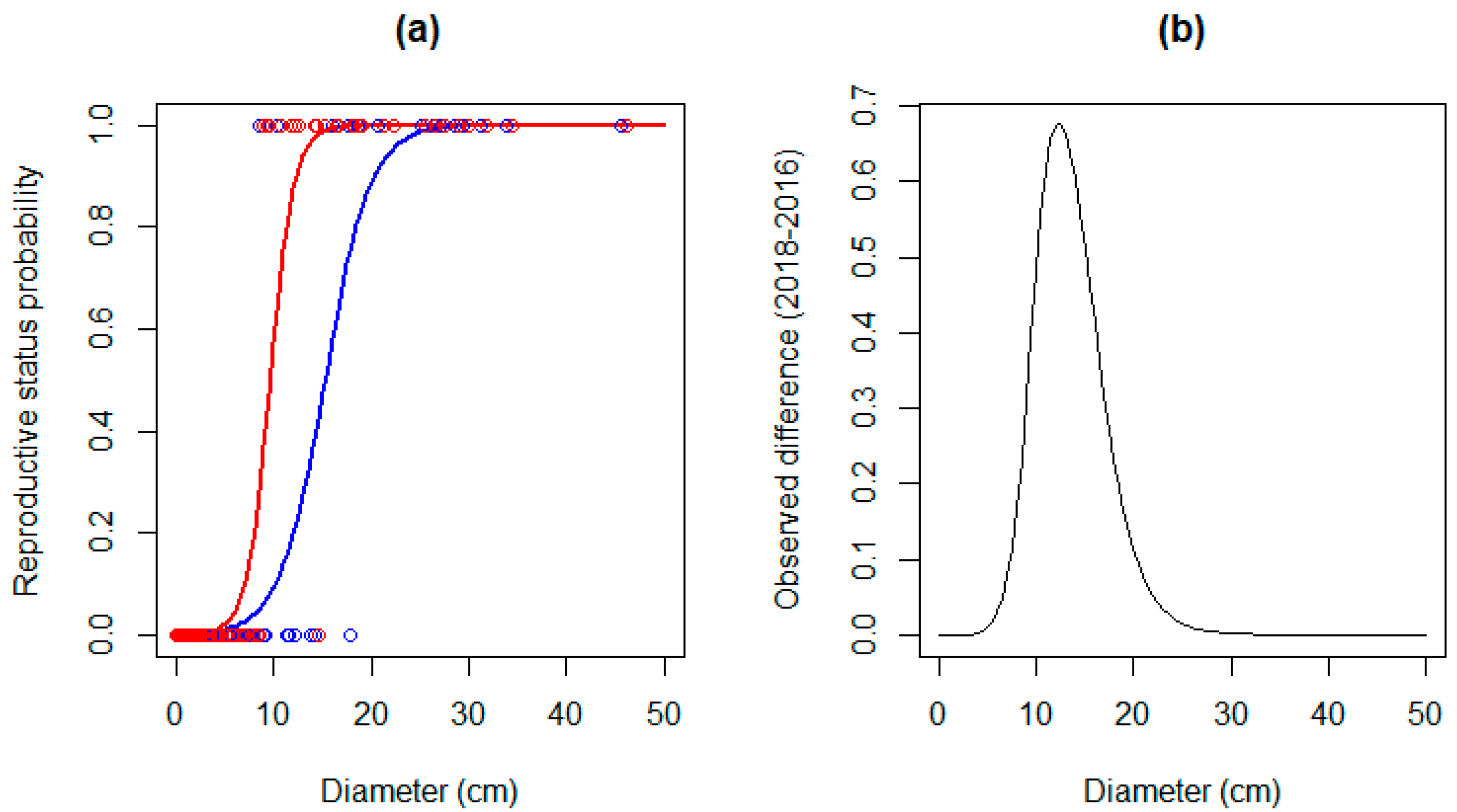
3.2. Observed Variations in the Reproductive Status
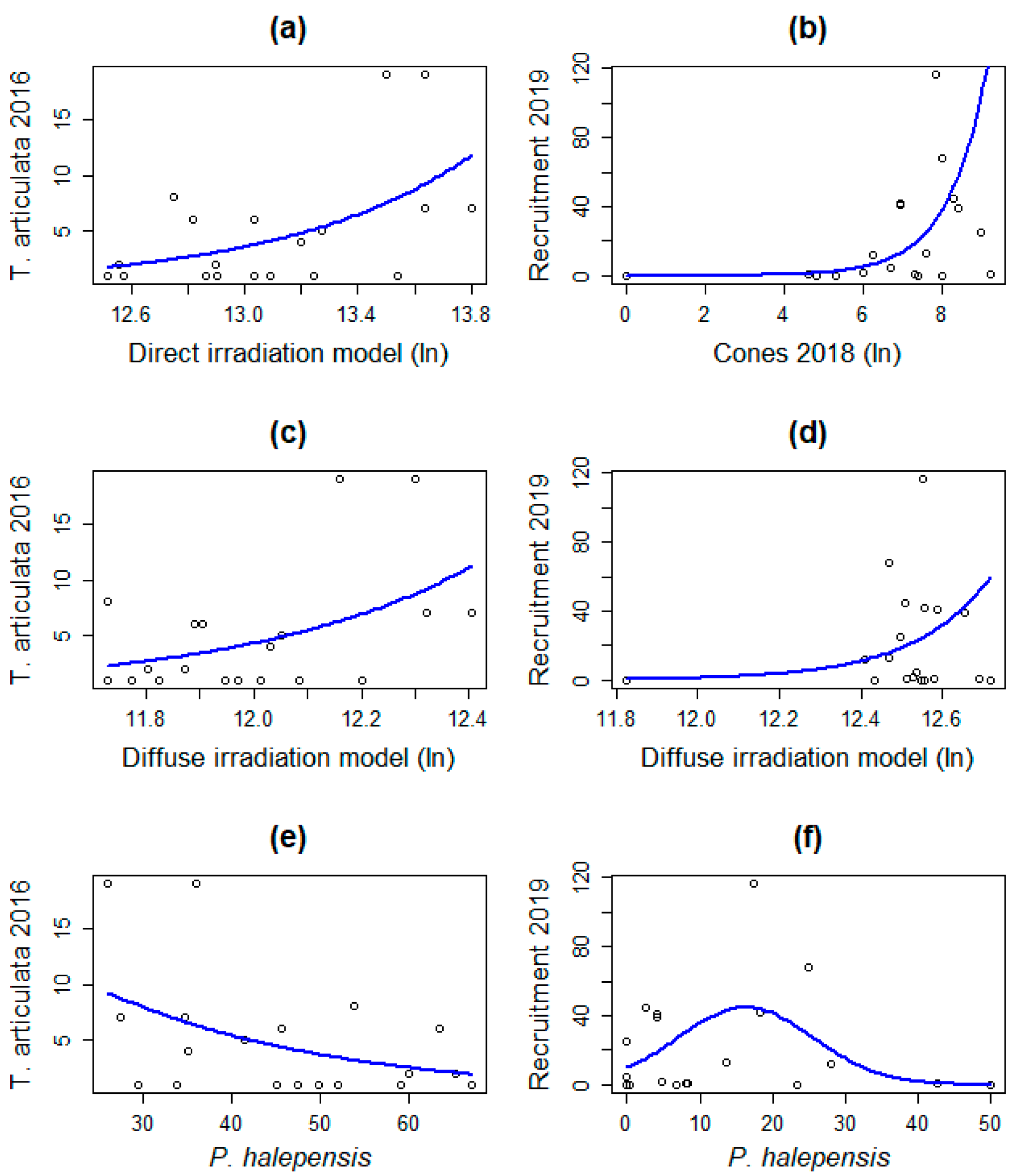
3.3. Demographic Response Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change. Climate Change 2013—The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef] [Green Version]
- Beniston, M.; Stephenson, D.B.; Christensen, O.B.; Ferro, A.T.; Frei, C.; Goyette, S.; Halsnaes, K.; Holt, T.; Jylhä, K.; Koffi, B.; et al. Future extreme events in European climate: An exploration of regional climate model projections. Clim. Chang. 2007, 81, 71–95. [Google Scholar] [CrossRef] [Green Version]
- Blenkinsop, S.; Fowler, H.J. Changes in European drought characteristics projected by the PRUDENCE regional climate models. Int. J. Climatol. 2007, 27, 1595–1610. [Google Scholar] [CrossRef]
- García, R.; Vilagrosa, A.; Alloza, J.A. Pine mortality in southeast Spain after an extreme dry and warm year: Interactions among drought stress, carbohydrates and bark beetle attack. Trees 2015, 29, 1791–1804. [Google Scholar] [CrossRef]
- Esteve-Selma, M.A.; Carreño-Fructuoso, M.F.; Moya-Pérez, J.M.; Montoya Bernabéu, P.F.; Martínez Fernández, J.; Pérez Navarro, M.A.; Lloret, F. Respuesta de los bosques de Pinus halepensis del sureste ibérico al cambio climático: Los eventos de sequía extrema. In El Clima: Aire, Agua, Tierra y Fuego, 1st ed.; Montávez, J.P., Gómez, J.J., López, J.M., Palacios, L., Turco, M., Jerez, S., Lorente, R., Jiménez Guerrero, P., Eds.; Publicaciones de la Asociación Española de Climatología (AEC) Serie A, no. 11. NIPO. Coed; Asociación Española de Climatología: Santander, Spain; Agencia Estatal de Meteorología: Leganés, Spain, 2018; pp. 1023–1033. Available online: http://aeclim.org/wp-content/uploads/2019/07/1023-ESTEVE.pdf (accessed on 14 September 2020).
- Pérez-Navarro, M.A.; Sapes, G.; Batllori, E.; Serra-Diaz, J.M.; Esteve, M.A.; Lloret, F. Climatic Suitability Derived from Species Distribution Models Captures Community Responses to an Extreme Drought Episode. Ecosystems 2019, 22, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Lloret, F. Vulnerability and resilience of forest ecosystems to extreme drought episodes. Ecosistemas 2012, 21, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Dargie, T.C.D. An ordination analysis of vegetation patterns on topoclimate gradients in South-east Spain. J. Biogeogr. 1987, 14, 197–211. [Google Scholar] [CrossRef]
- Chaparro, J. Distribución Potencial del Bosque y de Sus Especies Arbóreas en Zonas Mediterráneas Semiáridas: Modelos y Aplicaciones. Ph.D. Thesis, University of Murcia, Murcia, Spain, 1996. [Google Scholar]
- Esteve-Selma, M.A.; Chaparro, J.; Pardo, M.; Vives, R. Los sistemas forestales desde una perspectiva histórica: Las repoblaciones forestales. In Los Recursos Naturales de la Región de Murcia: Un Análisis Interdisciplinar, 1st ed.; Esteve-Selma, M.A., Lloréns, M., Martínez Gallur, C., Eds.; EDITUM; University of Murcia: Murcia, Spain, 2003. [Google Scholar]
- Oliveras, I.; Martínez-Vilalta, J.; Jimenez-Ortiz, T.; Lledó, M.J.; Escarré, A.; Piñol, J. Hydraulic properties of Pinus halepensis, Pinus pinea and Tetraclinis articulata in a dune ecosystem of Eastern Spain. Plant Ecol. 2003, 169, 131–141. [Google Scholar] [CrossRef]
- Esteve-Selma, M.A.; Martínez-Fernández, J.; Hernández-García, I.; Montávez, J.P.; López-Hernández, J.J.; Calvo, J.F. Potential effects of climatic change on the distribution of Tetraclinis articulata, an endemic tree from arid Mediterranean ecosystems. Clim. Chang. 2012, 113, 663–678. [Google Scholar] [CrossRef]
- Esteve-Selma, M.A.; Moya-Pérez, J.M.; Navarro-Cano, J.A. Manual de Evaluación y Gestión del Hábitat 9570*: Bosques de Tetraclinis Articulata, 1st ed.; Dirección General del Medio Natural, Región de Murcia: Murcia, Spain, 2019; p. 87. Available online: http://www.murcianatural.carm.es/c/document_library/get_file?uuid=15ccbf52-1211-4174-bc86-5572c2f662bc&groupId=14 (accessed on 10 November 2020).
- Simpson, J.D.; Powell, G.R. Some Factors Influencing Cone Production on Young Black Spruce in New Brunswick. For. Chron. 1981, 57, 267–269. [Google Scholar] [CrossRef]
- Despland, E.; Houle, G. Aspect influences cone abundance within the crown of Pinus banskiana Lamb. trees at the limit of the species distribution in northern Quebec (Canada). Écoscience 1997, 4, 521–525. [Google Scholar] [CrossRef]
- Nanda, K.K. Some observations on growth, branching behaviour and flowering of Teak (Tectona grandis LF.) in relation to light. Indian For. 1962, 88, 207–218. [Google Scholar]
- Chalupkla, W.; Giertych, M. Effect of polyethylene covers on the flowering of Norway spruce (Picea abies (L.) Karst.) grafts. Arbor. Korn. 1977, 22, 185–191. [Google Scholar] [CrossRef]
- Santos-del-Blanco, L.; Bonser, S.P.; Valladares, F.; Chambel, M.R.; Climent, J. Plasticity in reproduction and growth among 52 range-wide populations of a Mediterranean conifer: Adaptive responses to environmental stress. J. Evol. Biol. 2013, 26, 1912–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.G. Conifer Reproductive Biology, 1st ed.; Springer: Dordrecht, The Netherlands, 2009; p. 169. [Google Scholar] [CrossRef]
- Day, M.E.; Greenwood, M.S. Regulation of ontogeny in temperate conifers. In Size- and Age-Related Changes in Tree Structure and Function; Meinzer, F., Lachenbruch, B., Dawson, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 4, pp. 91–119. [Google Scholar] [CrossRef]
- Nyman, B. Effect of red and far-red irradiation on the germination process in seeds of Pinus sylvestris L. Nature 1961, 191, 1219–1220. [Google Scholar] [CrossRef]
- Loisel, R. Germination du pin d’Alep dans certaines associations végétales de Basse-Provence. Bull. Soc. Bot. Fr. 1967, 113, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Zamora, R.; Hodar, J.A.; Gómez, J.M. Ecology of seed germination of Pinus sylvestris L. at its southern, Mediterranean distribution range. For. Syst. 2005, 14, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Thanos, C.A.; Skordilis, A. The effects of light, temperature and osmotic stress on the germination of Pinus halepensis and Pinus brutia seeds. Seed Sci. Technol. 1987, 15, 163–174. [Google Scholar]
- Masetti, C.; Mencuccini, M. Régénération naturelle du Pin pignon (Pinus pinea L.) dans la Pineta Granducale di Alberese (Parco Naturale della Maremma, Toscana, Italie). Ecol. Mediterr. 1991, 17, 103–118. [Google Scholar] [CrossRef]
- Skordilis, A.; Thanos, C.A. Seed stratification and germination strategy in the Mediterranean pines Pinus brutia and Pinus halepensis. Seed Sci. Res. 1995, 5, 151–160. [Google Scholar] [CrossRef]
- Escudero, A.; Perez-Garcia, F.; Luzurlaga, A.L. Effects of lights, temperature and population variability on the germination of seven Spanish pines. Seed Sci. Res. 2002, 12, 261–271. [Google Scholar] [CrossRef]
- Hadjadj Aoul, S.; Chouieb, M.; Loisel, R. Effet des facteurs environnementaux sur les premiers stades de la régénération naturelle de Tetraclinis articulata (Vahl, Master) en Oranie, Algérie. Ecol. Mediterr. 2009, 35, 19–30. [Google Scholar] [CrossRef]
- Haddouche, I.; Benhanifia, K.; Gacemi, M. Analyse spatiale de la régénération forestière postincendie de la forêt de Fergoug à Mascara, Algérie. Bois For. Trop. 2011, 307, 23–31. [Google Scholar] [CrossRef]
- Dallahi, Y.; Chahhou, D.; El Aboudi, A.; Aafi, A.; Abbas, Y.; Mounir, F.; Abidine, M.M.O. The dynamics of natural regeneration of Tetraclinis articulata (Vahl) Masters in the Moroccan Central Plateau. Plant Sociol. 2017, 54, 37–41. [Google Scholar] [CrossRef]
- Nicolás, M.J.; Esteve, M.A.; Palazón, J.A.; López-Hernández, J.J. Modelo sobre las preferencias de hábitat a escala local de Tetraclinis articulata (Vahl) Masters en una población de su área de distribución. An. Biol. 2004, 26, 157–167. Available online: https://revistas.um.es/analesbio/article/view/30571 (accessed on 3 November 2020).
- Esteve Selma, M.A.; Montoya, P.; Moya, J.M.; Miñano, J.; Hernández, I.; Carrión, J.S.; Charco, J.; Fernández, S.; Munuera, M.; Ochando, J. Tetraclinis Articulata: Biogeografía, Ecología, Amenazas y Conservación, 1st ed.; Dirección General de Medio Natural, Región de Murcia: Murcia, Spain, 2017; p. 248. Available online: http://www.murcianatural.carm.es/c/document_library/get_file?uuid=6eeb4eb9-b1d1-4695-81e3-49fea2a002e8&groupId=14 (accessed on 4 February 2021).
- Petras, V.; Petrasova, A.; GRASS Development Team. Addon r.sun.daily. Geographic Resources Analysis Support System (GRASS) Software, Version 7.8. 2021. Available online: https://grass.osgeo.org/grass78/manuals/addons/r.sun.daily.html (accessed on 1 March 2021).
- Barilotti, A.; Turco, S.; Alberti, G. LAI determination in forestry ecosystem by LiDAR data analysis. In Proceedings of the International Workshop 3D Remote Sensing in Forestry, Vienna, Austria, 14–15 February 2006; Koukal, T., Scneider, W., Eds.; Institute of Surveying, Remote Sensing and Land Information, University of Natural Resources and Applied Life Sciences (BOKU): Vienna, Austria, 2006; pp. 259–263. Available online: https://boku.ac.at/fileadmin/data/H03000/H85000/H85700/workshops/3drsforestry/Proceedings_3D_Remote_Sensing_2006_rev_20070129.pdf (accessed on 20 September 2020).
- Bode, C.A.; Limm, M.P.; Power, M.E.; Finlay, J.C. Subcanopy solar radiation model: Predicting solar radiation across a heavily vegetated landscape using LiDAR and GIS solar radiation models. Remote Sens. Environ. 2014, 154, 387–397. [Google Scholar] [CrossRef]
- Mcgaughey, R.J. FUSION/LDV: Software for LIDAR data analysis and visualization; Version 4.20; USDA Forest Service, Pacific Northwest Research Station: Seattle, WA, USA, 2021; Available online: http://forsys.cfr.washington.edu/fusion/fusionlatest.html (accessed on 22 January 2021).
- Green, R.H. Sampling Design and Statistical Methods for Environmental Biologists, 1st ed.; John Wiley & Sons: Chichester, UK, 1979. [Google Scholar]
- Fox, J.; Weisberg, S.A. R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 16 July 2020).
- Nelder, J.; Wedderburn, R. Generalized Linear Models. J. R. Stat. Soc. 1972, 135, 370–384. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; Available online: http://www.stats.ox.ac.uk/pub/MASS4/ (accessed on 20 June 2020).
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model. Dev. 2015, 8, 2271–2312. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In Selected Papers of Hirotugu Akaike; Parzen, E., Tanabe, K., Kitagawa, G., Eds.; Springer: New York, NY, USA, 1998. [Google Scholar] [CrossRef]
- Messier, C.; Doucet, R.; Ruel, J.C.; Claveau, Y.; Kelly, C.; Lechowicz, M.J. Functional ecology of advance regeneration in relation to light in boreal forests. Can. J. For. Res. 1999, 29, 812–823. [Google Scholar] [CrossRef]
- Frochot, H.; Armand, G.; Gama, A.; Nouveau, M.; Wehrlen, L. La gestion de la vegetation accompagnatrice: État et perspective. Rev. For. Fr. 2002, 54, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Novoplansky, A. Picking battles wisely: Plant behavior under competition. Plant. Cell Environ. 2009, 32, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Yamawo, A. Relatedness of Neighboring Plants Alters the Expression of Indirect Defense Traits in an Extrafloral Nectary-Bearing Plant. Evol. Biol. 2015, 42, 12–19. [Google Scholar] [CrossRef]
- Moya-Pérez, J.M.; (University of Murcia, Murcia, Spain); Esteve-Selma, M.A.; (University of Murcia, Murcia, Spain). Personal communication. 2020. [Google Scholar]
- Belmonte-Serrato, F.; Romero-Díaz, A.; López-Bermúdez, F. Influence of the throughfall in the volume and water availability distribution in the mediterranean scrubland. In Proceedings of the Conference on Erosion and Land Degradation in the Mediterranean, Aveiro, Portugal, 14–18 June 1995; pp. 531–541. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. halepensis Canopy Cover (%) | Number of Plots (2016) | Number of Plots (2018) |
|---|---|---|
| 00–10 | 0 | 11 |
| 10–20 | 0 | 3 |
| 20–30 | 3 | 3 |
| 30–40 | 4 | 0 |
| 40–50 | 5 | 2 |
| 50–60 | 3 | 0 |
| >60 | 4 | 0 |
| ANOVA Test | Variable | F Value | p-Value |
|---|---|---|---|
| Control–Impact before | Direct solar irradiation | 1.5244 | 0.2328 |
| Reproductive specimens | 1.8 | 0.1963 | |
| Control–Impact after | Direct solar irradiation | 7.2772 | 0.0147 * |
| Reproductive specimens | 6.6977 | 0.0186 * | |
| Control before–Control after | Direct solar irradiation | 0.0002 | 0.9886 |
| Reproductive specimens | 1.8 | 0.1964 | |
| Impact before–Impact after | Direct solar irradiation | 13.038 | 0.0019 ** |
| Reproductive specimens | 6.6977 | 0.0186 * |
| Year | Total Reproductive Specimens | Specimens with Cone Production | Estimated Cone Production | Production Ratio Per Active Specimen | Recruited Seedlings |
|---|---|---|---|---|---|
| 2016 | 18 | 11 | 15,900 | 1445 | 19 |
| 2018 | 28 | 24 | 44,070 | 1836 | 41 * |
| 2019 | 28 | 6 | 7900 | 1317 | 411 |
| Variable | Coefficient | Std. Error | p-Value | Deviance |
|---|---|---|---|---|
| Diameter 2016 | Int. −6.5789 | Int. 1.7427 | Int. 0.000160 *** | 73.78 |
| Pr. 0.4318 | Pr. 0.1202 | Pr. 0.000329 *** | ||
| Diameter 2018 | Int. −7.8666 | Int. 2.2374 | Int. 0.000438 *** | 86.51 |
| Pr. 0.8167 | Pr. 0.2424 | Pr. 0.000756 *** |
| 2016 Predictors | Direct Irradiation | Diffuse Irradiation | Pine Canopy | Reproductive Status | Cone Production | Specimens Number |
|---|---|---|---|---|---|---|
| Direct irradiation | 1 | 0.8969 | −0.5934 | −0.0137 | 0.1335 | 0.5398 |
| Diffuse irradiation | - | 1 | −0.7631 | 0.0869 | 0.1264 | 0.4823 |
| Pine canopy | - | - | 1 | −0.2023 | −0.0399 | −0.4469 |
| Reproductive | - | - | - | 1 | 0.1577 | 0.4798 |
| Cone production | - | - | - | - | 1 | 0.0167 |
| Specimens number | - | - | - | - | - | 1 |
| 2019 Predictors | Direct Irradiation | Diffuse Irradiation | Pine Canopy | Reproductive Status | Cone Production | Recruits Number |
| Direct irradiation | 1 | 0.7200 | −0.4608 | 0.4222 | 0.3106 | −0.2014 |
| Diffuse irradiation | - | 1 | −0.6844 | 0.2503 | 0.3486 | 0.0947 |
| Pine canopy | - | - | 1 | −0.2175 | −0.3020 | −0.0058 |
| Reproductive | - | - | - | 1 | −0.0841 | 0.0706 |
| Cone production | - | - | - | - | 1 | 0.1469 |
| Recruits number | - | - | - | - | - | 1 |
| Predictors | Statistics | 2016 (Specimens) | 2019 (Recruitment) |
|---|---|---|---|
| Direct solar irradiation (ln) | Coefficients | Int. −18.0208 Pr. 1.4845 | - |
| Std. Errors | Int. 6.8406 Pr. 0.5193 | - | |
| p-values | Int. 0.00843 ** Pr. 0.00426 ** | - | |
| AIC | 100.28 | - | |
| Deviance | 32.83 | - | |
| Diffuse solar irradiation (ln) | Coefficients | Int. −26.992 Pr. 2.371 | Int. −63.365 Pr. 5.304 |
| Std. Errors | Int. 12.353 Pr. 1.027 | Int. 33.071 Pr. 2.691 | |
| p-values | Int. 0.0289 * Pr. 0.0209 * | Int. 0.0601 Pr. 0.0487 * | |
| AIC | 102.44 | 144.89 | |
| Deviance | 24.27 | 5.2 | |
| P. halepensis canopy (x + x2) | Coefficients | Int. 3.23552 Pr. −0.03858 | Int. 2.295034 Pr. 0.185123 Pr.2 −0.005683 |
| Std. Errors | Int. 0.75714 Pr. 0.01651 | Int. 0.628018 Pr. 0.089193 Pr.2 0.0023 | |
| p-values | Int. 1.93 × 10−5 *** Pr. 0.0195 * | Int. 0.000258 *** Pr. 0.037937 * Pr.2 0.013465 * | |
| AIC | 102.38 | 142.56 | |
| Deviance | 24.52 | 22.99 | |
| Σ Previous cones (ln) | Coefficients | - | Int. −4.4104 Pr. 1.0079 |
| Std. Errors | - | Int. 2.0563 Pr. 0.2805 | |
| p-values | - | Int. 0.031965 * Pr. 0.000327 *** | |
| AIC | - | 138.4 | |
| Deviance | - | 30.53 |
| Year | Predictors | Coefficients | Std. Errors | p-Values | VIF | AIC | Deviance |
|---|---|---|---|---|---|---|---|
| 2016 (specimens) | Direct solar irradiation (ln) Reproductive specimens | Int. −17.5961 Pr.a 1.4115 Pr.b 0.4881 | Int. 6.0281 Pr.a 0.4571 Pr.b 0.2297 | Int. 0.00351 ** Pr.a 0.00201 ** Pr.b 0.03362 * | Pr.a 1.0006 Pr.b 1.0006 | 97.8 | 48.15 |
| 2019 (recruitment) | Σ Previous cones (ln) P. halepensis canopy (x + x2) | Int. −4.7882 Pr.a 0.9022 Pr.b 0.2244 Pr.b2 −0.0062 | Int. 1.89764 Pr.a 0.24202 Pr.b 0.07978 Pr.b2 0.002129 | Int. 0.011627 * Pr.a 0.000193 *** Pr.b 0.004920 ** Pr.b2 0.003706 ** | Pr.a 1.0158 Pr.b 7.9568 Pr.b2 7.9198 | 134.91 | 52.97 |
| Year | Predictor | Coefficient | p-Value | AIC | Deviance |
|---|---|---|---|---|---|
| 2018 (recruitment) | Σ Flow lines (x + x2) | Int. 1.4662283 Pr. −0.0749441 Pr.2 0.004082 | Int. 0.0303 * Pr. 0.0235 * Pr.2 0.0104 * | 64.864 | 43.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya-Pérez, J.M.; Carreño, M.F.; Esteve-Selma, M.Á. Enhancing the Resilience of a Mediterranean Forest to Extreme Drought Events and Climate Change: Pinus—Tetraclinis Forests in Europe. Forests 2021, 12, 487. https://doi.org/10.3390/f12040487
Moya-Pérez JM, Carreño MF, Esteve-Selma MÁ. Enhancing the Resilience of a Mediterranean Forest to Extreme Drought Events and Climate Change: Pinus—Tetraclinis Forests in Europe. Forests. 2021; 12(4):487. https://doi.org/10.3390/f12040487
Chicago/Turabian StyleMoya-Pérez, Juan Miguel, María Francisca Carreño, and Miguel Ángel Esteve-Selma. 2021. "Enhancing the Resilience of a Mediterranean Forest to Extreme Drought Events and Climate Change: Pinus—Tetraclinis Forests in Europe" Forests 12, no. 4: 487. https://doi.org/10.3390/f12040487
APA StyleMoya-Pérez, J. M., Carreño, M. F., & Esteve-Selma, M. Á. (2021). Enhancing the Resilience of a Mediterranean Forest to Extreme Drought Events and Climate Change: Pinus—Tetraclinis Forests in Europe. Forests, 12(4), 487. https://doi.org/10.3390/f12040487