
Natural Seed Limitation and Effectiveness of Forest Plantations to Restore Semiarid Abandoned Metal Mining Areas in SE Spain



Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Natural Plant Colonization in the Mine Tailings
2.3. Plantation of the Tetraclinis Mixed Forest in a Mine Tailing
2.4. Statistical Analyses
2.4.1. Natural Colonization in the Mine Tailings
2.4.2. Plantation of the Tetraclinis Mixed Forest in Mine Tailing
3. Results
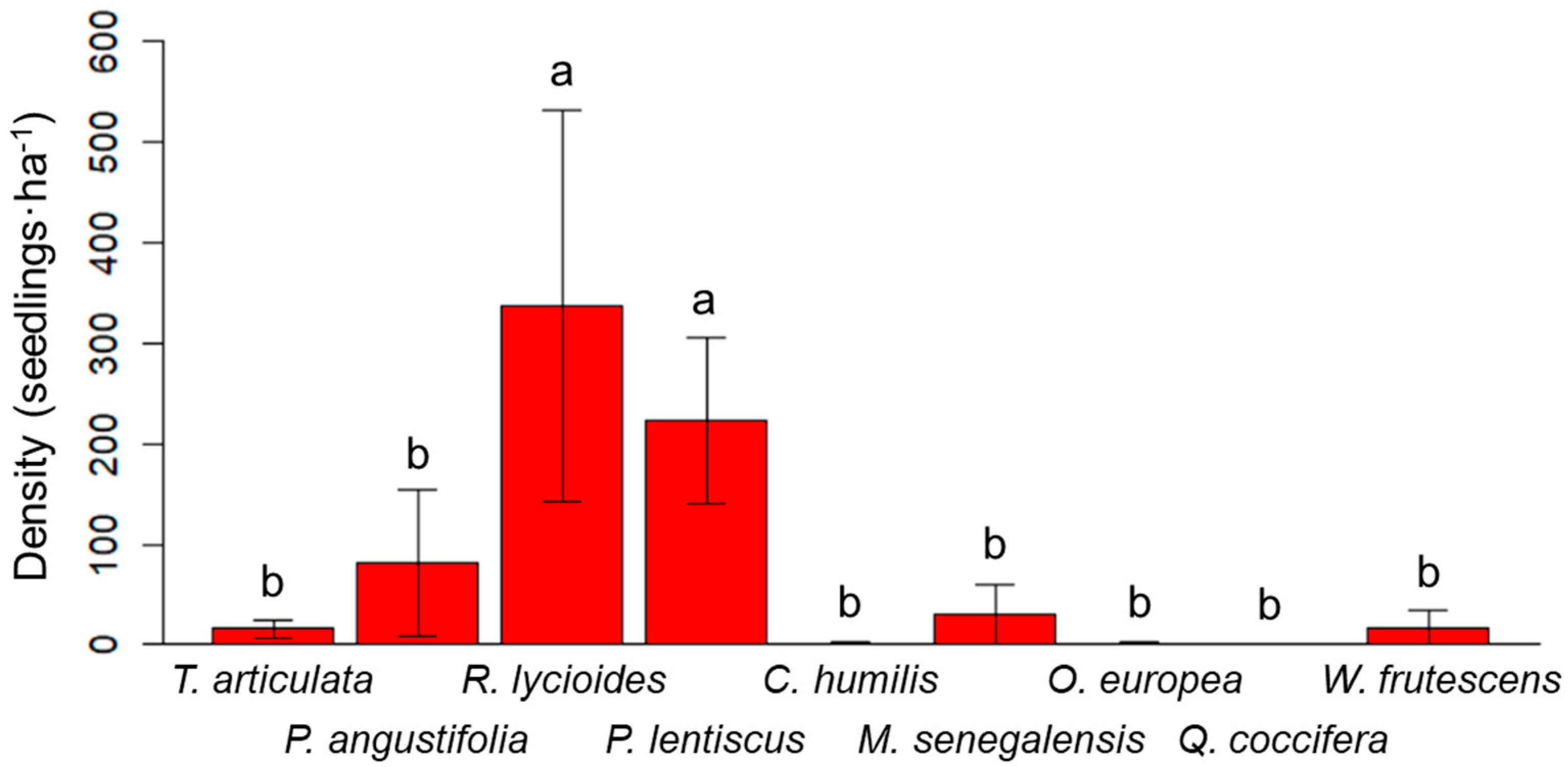
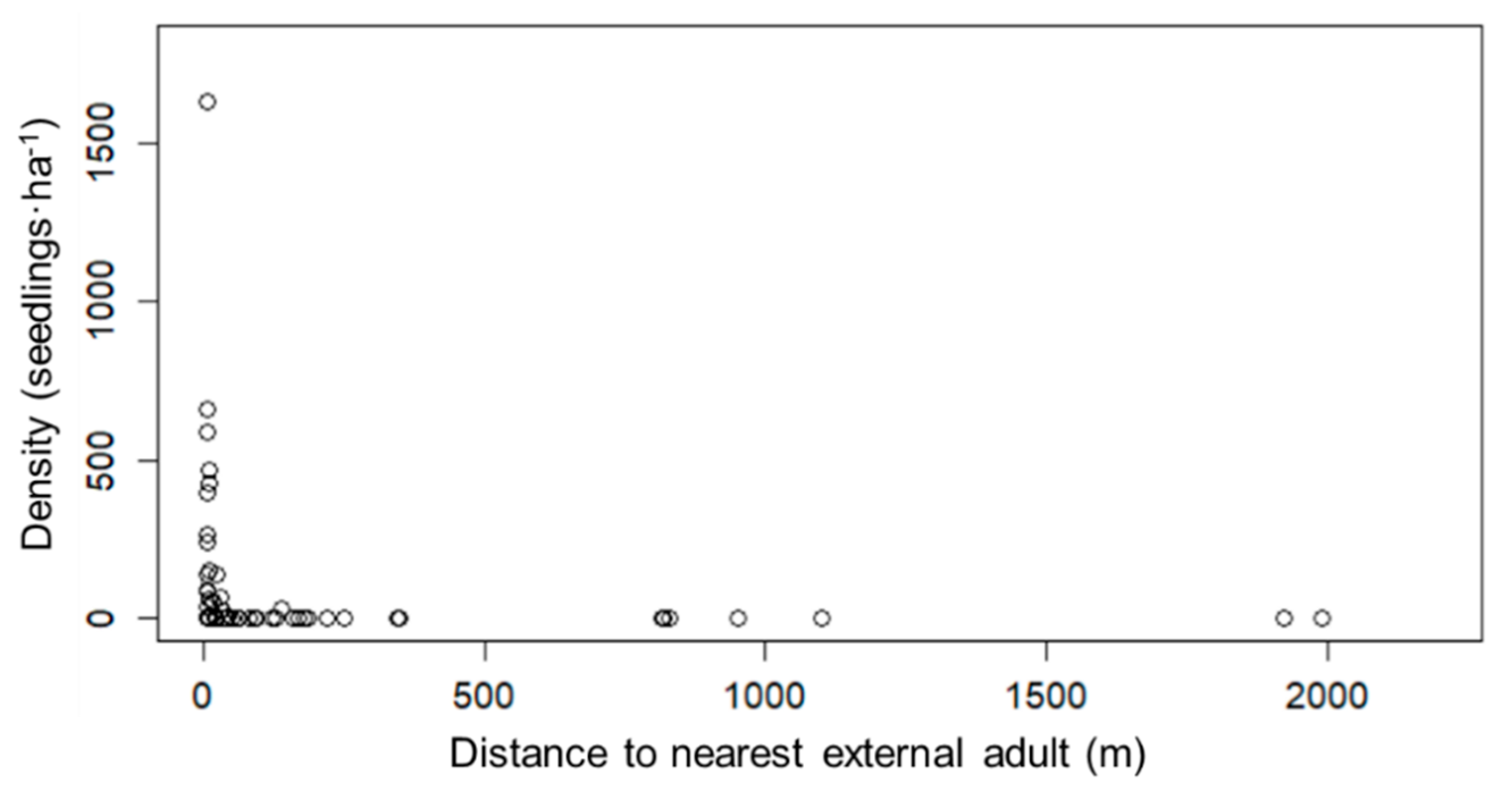
3.1. Natural Plant Colonization in the Mine Tailings
3.2. Plantation of the Tetraclinis Mixed Forest in a Mine Tailing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Metsaranta, J.M.; Beauchemin, S.; Langley, S.; Tisch, B.; Dale, P. Assessing the Long-Term Ecosystem Productivity Benefits and Potential Impacts of Forests Re-Established on a Mine Tailings Site. Forests 2018, 9, 707. [Google Scholar] [CrossRef] [Green Version]
- Panagos, P.; Van Liedekerke, M.; Yigini, Y.; Montanarella, L. Contaminated sites in Europe: Review of the current situation based on data collected through a European network. J. Environ. Public Health 2013, 2013, 158764. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cano, J.A.; Verdú, M.; Goberna, M. Trait-based selection of nurse plants to restore ecosystem functions in mine tailings. J. Appl. Ecol. 2018, 55, 1195–1206. [Google Scholar] [CrossRef]
- Colin, Y.; Goberna, M.; Verdú, M.; Navarro-Cano, J.A. Successional trajectories of soil bacterial communities in mine tailings: The role of plant functional traits. J. Environ. Manag. 2019, 241, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Oreja, B.; Goberna, M.; Verdú, M.; Navarro-Cano, J.A. Constructed pine log piles facilitate plant establishment in mining drylands. J. Environ. Manag. 2020, 271, 111015. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, S.; Clement, J.S.; Thibault, Y.; Langley, S.; Gregorich, E.G.; Tisch, B. Geochemical stability of acid-generating pyrrhotite tailings 4 to 5 years after addition of oxygen-consuming organic covers. Sci. Totol Environ. 2018, 645, 1643–1655. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Daniels, W.L. Estimation of carbon sequestration by pine (Pinus sylvestris L.) ecosystems developing on reforested post-mining sites in Poland on differing mine soil substrates. Ecol. Eng. 2014, 73, 209–218. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Krzaklewski, K. Potential for carbon sequestration in reclaimed mine soil on reforested surface mining areas in Poland. Nat. Sci. 2010, 2, 1015–1021. [Google Scholar] [CrossRef]
- Rodrigue, J.A.; Burger, J.A.; Oderwald, R.G. Forest productivity and commercial value of pre-law reclaimed mined land in the Eastern United States. North. J. App. For. 2002, 19, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.J.; Currey, N.A. Engineering closure of an open pit gold operation in a semi-arid climate. Int. J. Surf. Min. Reclam. Environ. 2002, 16, 270–288. [Google Scholar] [CrossRef]
- Mendez, M.O.; Maier, R.M. Phytostabilization of mine tailings in arid and semiarid environments—An emerging remediation technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Martínez, S.; Zornoza, R.; Gabarrón, M.; Gómez-Garrido, M.; Rosales, R.M.; Muñoz, M.A.; Gómez-López, M.D.; Soriano-Disla, J.M.; Faz, A.; Acosta, J.A. Is aided phytostabilization a suitable technique for the remediation of tailings? Eur. J. Soil Sci. 2019, 70, 862–875. [Google Scholar] [CrossRef]
- Navarro-Cano, J.A.; Goberna, M.; Valiente-Banuet, A.; Verdu, M. Same nurse but different time: Temporal divergence in the facilitation of plant lineages with contrasted functional syndromes. Funct. Ecol. 2016, 30, 1854–1861. [Google Scholar] [CrossRef]
- Gómez-Ros, J.M.; Garcia, G.; Peñas, J.M. Assessment of restoration success of former metal mining areas after 30 years in a highly polluted Mediterranean mining area: Cartagena-La Unión. Ecol. Eng. 2013, 57, 393–402. [Google Scholar] [CrossRef]
- Shuman, L.M. Organic waste amendments effect on zinc fractions of two soils. J. Environ. Qual. 1999, 28, 1442–1447. [Google Scholar] [CrossRef]
- Walker, D.J.; Clemente, R.; Bernal, M.P. Contrasting effects of manure and compost on soil pH, heavy metal availability and growth of Chenopodium album L. in a soil contaminated by pyritic mine waste. Chemosphere 2004, 57, 215–224. [Google Scholar] [CrossRef]
- Zornoza, R.; Faz, A.; Carmona, D.M.; Martínez-Martínez, S.; Acosta, J.A. Plant cover and soil biochemical properties in a mine tailing pond five years after application of marble wastes and organic amendments. Pedosphere 2012, 22, 22–32. [Google Scholar] [CrossRef]
- Park, J.H.; Lamb, D.; Paneerselvam, P.; Choppala, G.; Bolan, N.; Chung, J.W. Role of organic amendments on enhanced bioremediation of heavy metal (loid) contaminated soils. J. Hazard. Mater. 2011, 185, 549–574. [Google Scholar] [CrossRef] [PubMed]
- Tordof, G.M.; Baker, A.J.M.; Willis, A.J. Current approaches to the revegetation and reclamation of metalliferous mine wastes. Chemosphere 2000, 41, 219–228. [Google Scholar] [CrossRef]
- Pond, A.P.; White, S.A.; Milczarek, M.; Thompson, T.L. Accelerated weathering of biosolid-amended copper mine tailings. J. Environ. Qual. 2005, 34, 1293–1301. [Google Scholar] [CrossRef] [Green Version]
- Parra, A.; Zornoza, R.; Conesa, E.; Faz, A.; Gómez-López, M.D. Nutritional status and its interaction with soil properties and trace elements in six Mediterranean shrub species grown in reclaimed pyritic tailings. Ecol. Eng. 2017, 109, 25–34. [Google Scholar] [CrossRef]
- Bainbridge, D.A. A Guide for Desert and Dryland Restoration: New Hope for Arid Lands; Island Press: Washington, DC, USA, 2007. [Google Scholar]
- Hou, X.; Liu, S.; Zhao, S.; Beazley, R.; Cheng, F.; Wu, X.; Xu, J.; Dong, S. Selection of suitable species as a key factor for vegetation restoration of degraded areas in an open-pit manganese-ore mine in Southern China using multivariate-analysis methods. Land Degrad. Dev. 2019, 30, 942–950. [Google Scholar] [CrossRef]
- Párraga-Aguado, I.; González-Alcaraz, M.N.; Álvarez-Rogel, J.; Jiménez-Cárceles, F.J.; Conesa, H.M. The importance of edaphic niches and pioneer plant species succession for the phytomanagement of mine tailings. Environ. Pollut. 2013, 176, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cano, J.A.; Goberna, M.; Verdú, M. Using plant functional distances to select species for restoration of mining sites. J. Appl. Ecol. 2019, 56, 2353–2362. [Google Scholar] [CrossRef]
- Conesa, H.M.; Schulin, R. The Cartagena–La Unión mining district (SE Spain): A review of environmental problems and emerging phytoremediation solutions after fifteen years research. J. Environ. Monit. 2010, 12, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Rogel, J.; Peñalver-Alcalá, A.; Jiménez-Cárceles, F.J.; Tercero, M.C.; González-Alcaraz, M.N. Evidence supporting the value of spontaneous vegetation for phytomanagement of soil ecosystem functions in abandoned metal(loid) mine tailings. Catena 2021, 201, 105191. [Google Scholar] [CrossRef]
- Lee, J.S.; Chon, H.T. Exposure assessment of heavy metals on abandoned metal mine areas by ingestion of soil, crop plant and groundwater. J. Phys. IV 2003, 107, 757–760, EDP sciences. [Google Scholar] [CrossRef]
- González-Alcaraz, M.N.; van Gestel, C.A.M. Climate change effects on enchytraeid performance in metal-polluted soils explained from changes in metal bioavailability and bioaccumulation. Environ. Res. 2015, 142, 177–184. [Google Scholar] [CrossRef]
- Gutiérrez, M.; Mickus, K.; Camacho, L.M. Abandoned PbZn mining wastes and their mobility as proxy to toxicity: A review. Sci. Total Environ. 2016, 565, 392–400. [Google Scholar] [CrossRef]
- Blondet, I.; Schreck, E.; Viers, J.; Casas, S.; Jubany, I.; Bahí, N.; Zouiten, C.; Dufréchou, G.; Freydier, R.; Galy-Lacaux, C.; et al. Atmospheric dust characterisation in the mining district of Cartagena-La Unión, Spain: Air quality and health risks assessment. Sci. Total Environ. 2019, 693, 133496. [Google Scholar] [CrossRef]
- Wang, P.; Sun, Z.; Hu, Y.; Cheng, H. Leaching of heavy metals from abandoned mine tailings brought by precipitation and the associated environmental impact. Sci. Total Environ. 2019, 695, 133893. [Google Scholar] [CrossRef]
- Esteve-Selma, M.A.; Martínez-Fernández, J.; Hernández-García, I.; Montávez, J.P.; López-Hernández, J.J.; Calvo, J.F. Potential effects of climatic change on the distribution of Tetraclinis articulata, an endemic tree from arid Mediterranean ecosystems. Clim. Chang. 2012, 113, 663–678. [Google Scholar] [CrossRef]
- European Commission. Interpretation Manual of European Union Habitats; EC DG Environment: Brussels, Belgium, 2013. [Google Scholar]
- Esteve-Selma, M.A.; Moya-Pérez, J.M.; Navarro-Cano, J.A. Manual de Evaluación y Gestión del Hábitat 9570: Bosques de Tetraclinis Articulata; Dirección General del Medio Natural: Murcia, Spain, 2019; p. 87. [Google Scholar]
- Navarro-Cano, J.A.; Goberna, M.; Valiente-Banuet, A.; Verdú, M. Phenotypic structure of plant facilitation networks. Ecol. Lett. 2021, 24, 509–519. [Google Scholar] [CrossRef]
- Navarro-Cano, J.A.; Verdú, M.; García, C.; Goberna, M. What nurse shrubs can do for barren soils: Rapid productivity shifts associated with a 40 years ontogenetic gradient. Plant. Soil 2015, 388, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Prikhodko, V.E.; Azarenko, Y.A.; Shayakhmetov, M.R.; Tishkin, A.A.; Gorbunov, V.V.; Pivovarova, E.G. Reconstruction of the climate of the Medieval Epoch based on soil and geochemical studies of kurgans of the Srostki Culture in the South of Western Siberia. Eurasian Soil Sci. 2020, 53, 267–282. [Google Scholar] [CrossRef]
- Crommentuijn, T.; Polder, M.D.; van de Plassche, E.J. Maximum Permissible Concentrations and Negligible Concentrations for Metals, Taking Background Concentrations into Account; RIVM Report Number 601501001; National Institute of Public Health and the Environment Bilthoven: Bilthoven, The Netherlands, 1997; p. 260. Available online: https://rivm.openrepository.com/handle/10029/10113 (accessed on 27 January 2021).
- Aguilar, J.; Dorronsoro, C.; Galán, E.; Gómez Ariza, J.L. Los criterios y estándares para declarar un suelo como contaminado en Andalucía y la metodología y técnica de toma de muestras y análisis para su investigación. In Investigación y Desarrollo Medioambiental en Andalucía; OTRI. Universidad de Sevilla: Sevilla, Spain, 1999; p. 253. Available online: http://www.ugr.es/~fjmartin/INFORMES/Criterios%20y%20estandares.pdf (accessed on 28 January 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 20 September 2020).
- Oksanen, J.; Guillaume, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5–6. Available online: https://CRAN.Rproject.org/package=vegan (accessed on 1 October 2020).
- Azcon-Bieto, J.; Talón, M. Fundamentos de Fisiología Vegetal; McGraw-Hill: Barcelona, Spain, 2000. [Google Scholar]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 3rd ed.; CRC Press LLC: Boca Raton, FL, USA, 2001. [Google Scholar]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M.I.R.; Syeed, S.; Khan, N.A. Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant. Sci. 2012, 3, 1476–1489. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jia, Y.; Dong, R.; Huang, R.; Liu, P.; Li, X.; Wang, Z.; Liu, G.; Chen, Z. Advances in the mechanisms of plant tolerance to manganese toxicity. Int. J. Mol. Sci. 2019, 20, 5096. [Google Scholar] [CrossRef] [Green Version]
- Anjum, N.A.; Singh, H.P.; Khan, M.I.R.; Masood, A.; Per, T.S.; Negi, A.; Batish, D.R.; Khan, N.A.; Duarte, A.C.; Pereira, E.; et al. Too much is bad—An appraisal of phytotoxicity of elevated plant-beneficial heavy metal ions. Environ. Sci. Pollut. Res. 2015, 22, 3361–3382. [Google Scholar] [CrossRef]
- Rey, P.J.; Alcántara, J.M. Recruitment dynamics of a fleshy-fruited plant (Olea europaea): Connecting patterns of seed dispersal to seedling establishment. J. Ecol. 2000, 88, 622–633. [Google Scholar] [CrossRef]
- Young, T.P.; Petersen, D.A.; Clary, J.J. The ecology of restoration: Historical links, emerging issues and unexplored realms. Ecol. Lett. 2005, 8, 662–673. [Google Scholar] [CrossRef]
- Hardesty, B.D.; Hubbell, S.P.; Bermingham, E. Genetic evidence of frequent long-distance recruitment in a vertebrate-dispersed tree. Ecol. Lett. 2006, 9, 516–525. [Google Scholar] [CrossRef]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef]
- Walker, L.R.; Del Moral, R. Primary Succession and Ecosystem Rehabilitation; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Aradottir, A.L.; Halldorsson, G. Colonization of woodland species during restoration: Seed or safe site limitation? Rest. Ecol. 2008, 26, S73–S83. [Google Scholar] [CrossRef]
- Herrera, C.M. Plant-vertebrate seed dispersal systems in the Mediterranean: Ecological, evolutionary, and historical determinants. Ann. Rev. Ecol. System. 1995, 26, 705–727. [Google Scholar] [CrossRef]
- Debussche, M.; Escarré, J.; Lepart, J. Ornithochory and plant succession in Mediterranean abandoned orchards. Vegetatio 1982, 48, 255–266. [Google Scholar]
- Jordano, P.; García, C.; Godoy, J.A.; García-Castaño, J.L. Differential contribution of frugivores to complex seed dispersal patterns. Proc. Nat. Acad. Sci. USA 2007, 104, 3278–3282. [Google Scholar] [CrossRef] [Green Version]
- Zapata, V.M.; Robledano, F.; Ramos, V.; Martínez-López, V. Bird-mediated seed dispersal of fleshy fruits of Mediterranean shrubs in semiarid forest patches: The role of Pinus halepensis Miller trees as seed receptors. Plant. Ecol. 2014, 215, 1337–1350. [Google Scholar] [CrossRef]
- McConkey, K.R.; Prasad, S.; Corlett, R.T.; Campos-Arceiz, A.; Brodie, J.F.; Rogers, H.; Santamaria, L. Seed dispersal in changing landscapes. Biol. Conserv. 2012, 146, 1–13. [Google Scholar] [CrossRef]
- Padilla, F.M.; Pugnaire, F.I. El uso de especies arbustivas para la restauración de la cubierta vegetal en ambientes semiáridos. Cuad. Soc. Esp. Cie. For. 2004, 17, 103–107. [Google Scholar]
- Padilla, F.M.; Ortega, R.; Sánchez, J.; Pugnaire, F.I. Rethinking species selection for restoration of arid shrublands. Basic Appl. Ecol. 2009, 10, 640–647. [Google Scholar] [CrossRef] [Green Version]
- Vilagrosa, A.; Caturla, R.N.; Hernández, N.; Cortina, J.; Bellot, J.; Vallejo, V.R. Reforestación en ambiente semiárido del sureste peninsular. Resultados de las investigaciones desarrolladas para optimizar la supervivencia y el crecimiento de especies autóctonas. In Montes para la sociedad del nuevo milenio. III Congreso Forestal Español. Junta Andal. Cons. Medio Ambiente 2001, 4, 5–10. [Google Scholar]
- Maestre, F.T.; Bautista, S.; Cortina, J.; Bellot, J. Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecol. Appl. 2001, 11, 1641–1655. [Google Scholar] [CrossRef]
- Párraga-Aguado, I.; Querejeta, J.J.; González-Alcaraz, M.N.; Jiménez-Cárceles, F.J.; Conesa, H.M. Usefulness of pioneer vegetation for the phytomanagement of metal(loid)s enriched tailings: Grasses vs. shrubs vs. trees. J. Environ. Manag. 2014, 133, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Oliet, J.A.; Puértolas, J.; Planelles, R.; Jacobs, D.F. Nutrient loading of forest tree seedlings to promote stress resistance and field performance: A Mediterranean perspective. New For. 2013, 44, 649–669. [Google Scholar] [CrossRef]
- Achak, N.; Romane, A.; Alifriqui, M.; Markouk, M. Chemical composition, organic and mineral contents of leaves of Tetraclinis articulata (Vahl) masters from the Tensift-Al Haouz, Marrakech region (Morocco). J. Essent. Oil-Bear. Plants 2009, 12, 198–204. [Google Scholar] [CrossRef]
- Párraga-Aguado, I.; González-Alcaraz, M.N.; Álvarez-Rogel, J.; Conesa, H.M. Assessment of the employment of halophyte plant species for the phytomanagement of mine tailings in semiarid areas. Ecol. Eng. 2014, 71, 598–604. [Google Scholar] [CrossRef]
- Domínguez, M.T.; Marañón, T.; Murillo, J.M.; Schulin, R.; Robinson, B.H. Nutritional status of Mediterranean trees growing in a contaminated and remediated area. Water Air Soil Poll. 2010, 205, 305–321. [Google Scholar] [CrossRef]
- Hu, Y.L.; Mgelwa, A.S.; Singh, A.N.; Zeng, D.H. Differential responses of the soil nutrient status, biomass production, and nutrient uptake for three plant species to organic amendments of placer gold mine-tailing soils. Land Degrad. Dev. 2018, 29, 2836–2845. [Google Scholar] [CrossRef]
- Arco-Lázaro, E.; Martínez-Fernández, D.; Bernal, M.P.; Clemente, R. Response of Piptatherum miliaceum to co-culture with a legume species for the phytostabilisation of trace elements contaminated soils. J. Soils Sediments 2017, 17, 1349–1357. [Google Scholar] [CrossRef]
- Ross, S.M. Retention, transformation and mobility of toxic metals in soils. In Toxic Metals in Soil-Plant Systems; Ross, S.M., Ed.; John Wiley and Sons Ltd.: Chichester, UK, 1994; pp. 63–152. [Google Scholar]
- Disante, K.B.; Fuentes, D.; Cortina, J. Sensitivity to zinc of Mediterranean woody species important for restoration. Sci. Total Environ. 2010, 408, e2225. [Google Scholar] [CrossRef]
- Almås, Å.; Singh, B.R.; Salbu, B. Mobility of cadmium-109 and zinc-65 in soil influenced by equilibration time, temperature, and organic matter. J. Environ. Qual. 1999, 28, 1742–1750. [Google Scholar] [CrossRef]
- Kohler, J.; Caravaca, F.; Azcón, R.; Díaz, G.; Roldán, A. The combination of compost addition and arbuscular mycorrhizal inoculation produced positive and synergistic effects on the phytomanagement of a semiarid mine tailing. Sci. Total Environ. 2015, 514, 42–48. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mine Tailing | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Composted Waste | Clarkelithosphere | MPC | MRV | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil Properties | |||||||||||||
| TOC (g 100 g−1) | 0.836 | 0.664 | 0.670 | 0.542 | 0.592 | 0.782 | 0.670 | 0.153 | 19.42 | ||||
| N (g 100 g−1) | 0.054 | 0.047 | 0.079 | 0.045 | 0.087 | 0.044 | 0.045 | 0.018 | 1.89 | ||||
| P (g 100 g−1) | 0.023 | 0.001 | 0.044 | 0.001 | 0.043 | 0.001 | 0.026 | 0.005 | 0.35 | ||||
| K (g 100 g−1) | 0.187 | 0.255 | 0.327 | 0.093 | 0.486 | 0.222 | 0.247 | 0.071 | 0.71 | ||||
| pH | 7.92 | 7.86 | 7.05 | 7.69 | 6.69 | 7.31 | 7.66 | 6.68 | 6.5 | ||||
| EC (dS m−1) | 1.43 | 3.89 | 1.06 | 3.15 | 1.80 | 2.58 | 2.48 | 1.48 | 4.1 | ||||
| As (mg kg−1) | 355 | 167 | 232 | 120 | 260 | 153 | 197 | 93 | <0.01 | 5.6 | 34 | 100 | |
| Cd (mg kg−1) | 10.8 | 36.0 | 2.3 | 37.0 | 23.3 | 20.7 | 15.2 | 15.6 | 2.7 | 0.09 | 1.6 | 15 | |
| Cu (mg kg−1) | 19.2 | 86.5 | 35.0 | 106.6 | 94.2 | 404.0 | 38.0 | 46.8 | 110.0 | 27 | 40 | 500 | |
| Fe (mg kg−1) | 124,554 | 129,326 | 97,636 | 184,264 | 95,165 | 144,816 | 136,255 | 146,313 | 9689 | 465 | |||
| Mn (mg kg−1) | 4529 | 5246 | 2413 | 9086 | 1695 | 7386 | 6189 | 8148 | 186 | 7.7 | |||
| Zn (mg kg−1) | 4851 | 13,288 | 638 | 14,619 | 10,681 | 10,405 | 6653 | 8082 | 272 | 75 | 160 | 1000 | |
| Pb (mg kg−1) | 2076 | 5430 | 499 | 21241 | 4347 | 8915 | 3791 | 4913 | 79 | 17 | 140 | 1000 | |
| Factors | Macronutrients | ||||||||
| N | P | K | C:N | ||||||
| Organic Amendment | 5.27 * | 0.29 | 0.72 | 6.34 * | |||||
| Species Identity | 4.70 *** | 5.12 *** | 6.62 *** | 5.14 *** | |||||
| Amendment x Species | 0.35 | 3.15 ** | 0.50 | 0.44 | |||||
| Factors | Metal(oid)s | ||||||||
| As | Cd | Cu | Fe | Mn | Pb | Zn | |||
| Organic Amendment | 0.28 | 0.03 | 5.58 * | 0.02 | 0.09 | 0.02 | 0.12 | ||
| Species Identity | 5.11 *** | 9.99 *** | 13.78 *** | 5.16 *** | 18.91 *** | 8.95 *** | 4.28 *** | ||
| Amendment x Species | 1.21 | 0.81 | 0.92 | 2.18 * | 2.01 ˙ | 1.98 ˙ | 0.43 | ||
| Species | Macronutrients | Metal(oid)s | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | C:N | As | Cd | Cu | Fe | Mn | Pb | Zn | |
| Tetraclinis articulata | ↑ | ↓ | |||||||||
| Rhamnus lycioides | ↓ | ↑ | |||||||||
| Pistacia lentiscus | ↓ | ||||||||||
| Olea europaea | ↑ | ||||||||||
| Chamaerops humilis | |||||||||||
| Maytenus senegalensis | ↓ | ↑ | |||||||||
| Periploca angustifolia | ↑ | ↑ | ↑ | ↑ | |||||||
| Quercus coccifera | ↓ | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oná, M.B.; Goberna, M.; Navarro-Cano, J.A. Natural Seed Limitation and Effectiveness of Forest Plantations to Restore Semiarid Abandoned Metal Mining Areas in SE Spain. Forests 2021, 12, 548. https://doi.org/10.3390/f12050548
Oná MB, Goberna M, Navarro-Cano JA. Natural Seed Limitation and Effectiveness of Forest Plantations to Restore Semiarid Abandoned Metal Mining Areas in SE Spain. Forests. 2021; 12(5):548. https://doi.org/10.3390/f12050548
Chicago/Turabian StyleOná, Marta Bindang, Marta Goberna, and Jose Antonio Navarro-Cano. 2021. "Natural Seed Limitation and Effectiveness of Forest Plantations to Restore Semiarid Abandoned Metal Mining Areas in SE Spain" Forests 12, no. 5: 548. https://doi.org/10.3390/f12050548
APA StyleOná, M. B., Goberna, M., & Navarro-Cano, J. A. (2021). Natural Seed Limitation and Effectiveness of Forest Plantations to Restore Semiarid Abandoned Metal Mining Areas in SE Spain. Forests, 12(5), 548. https://doi.org/10.3390/f12050548