
Estimation of the Productivity Potential of Mountain Sites (Mixed Beech-Coniferous Stands) in the Romanian Carpathians

 ,
, 
Abstract
:1. Introduction
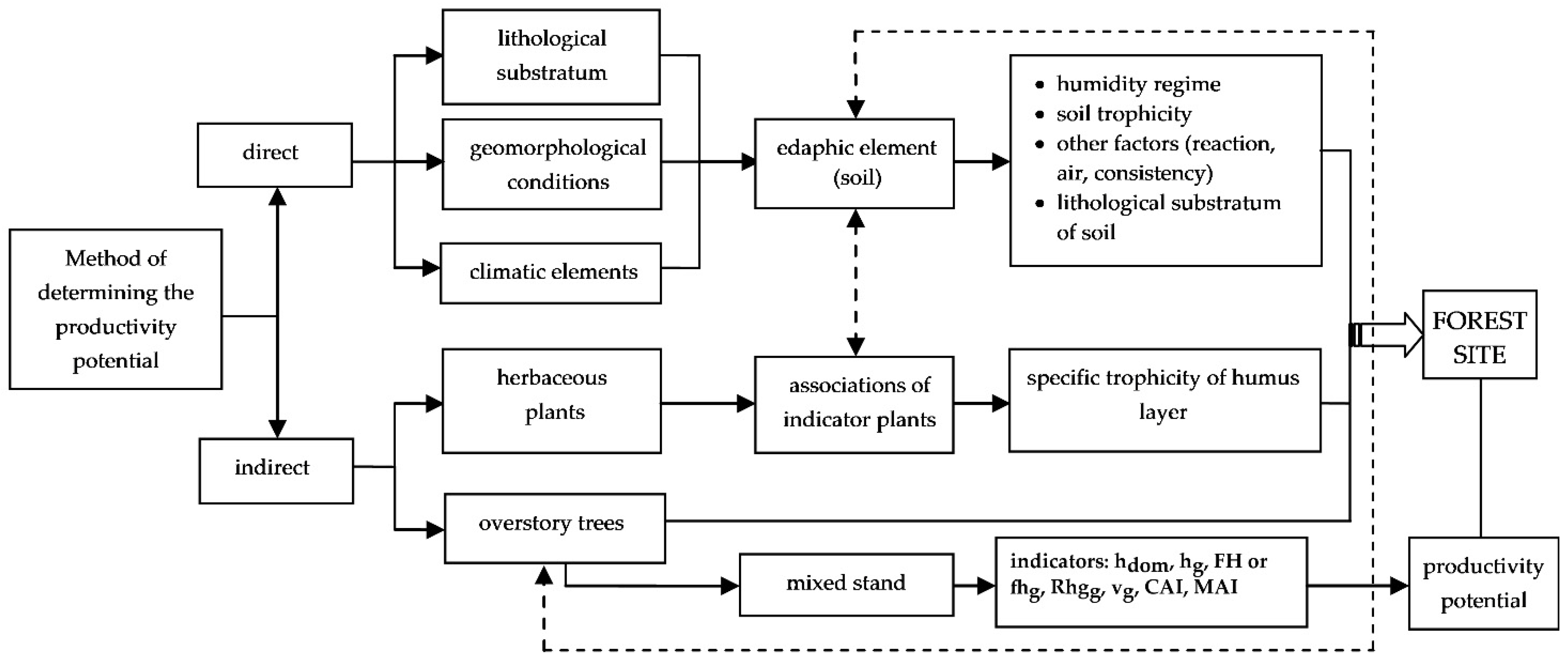
2. Materials and Methods
- -
- Model 1: f (x) = a + bx + cx2, between the variables: hdom and T, hg and T, fhg and T, hg and dg, fhg and dg, vg and gg, with x being the age T, dg, or gg;
- -
- Model 2: f (x) = ae-bx, between the variables: Rhgg and dg, Rhgg and T, with x being dg or T;
- -
- Model 3: f (x) = a + bx + cx2 + dx3, between the variables: current annual volume increment (CAI) and T, MAI and T, with x being T.
3. Results
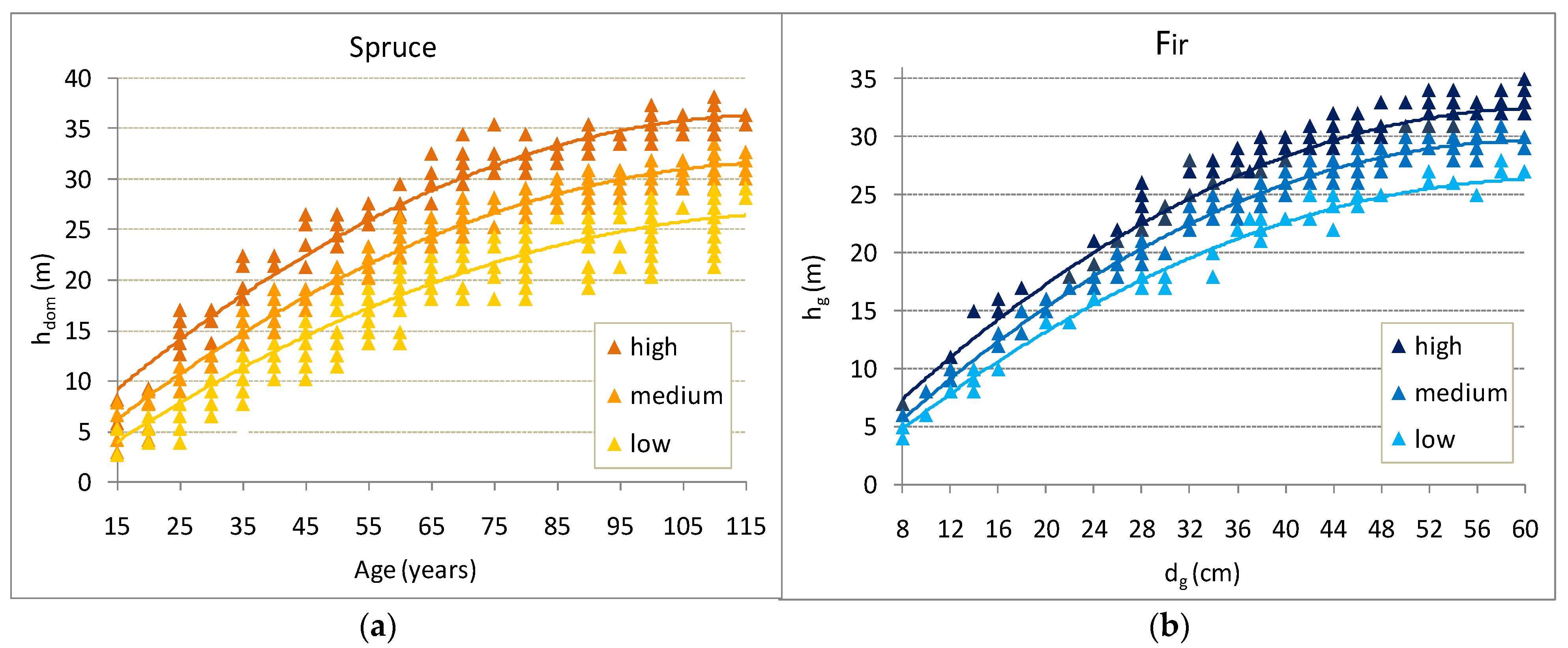
3.1. Height and Form Height (FH) of Stand as an Indicator of Site Productivity
3.2. Ratio between Height of the Mean-Basal-Area Tree of the Stand and Its Basal Area (Rhgg)
3.3. Volume of the Mean Tree (vg)
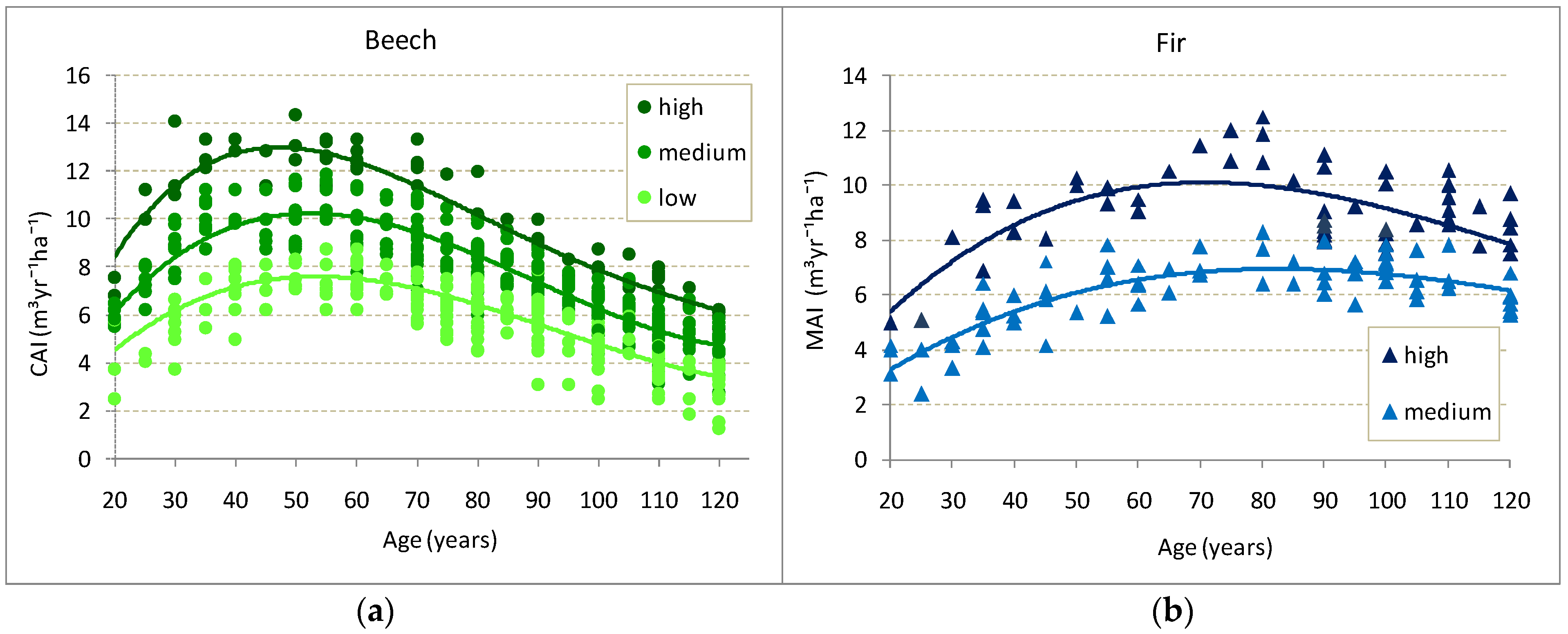
3.4. Volume Increment of the Stand
3.5. Precision of Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiriță, C.; Vlad, I.; Păunescu, C.; Pătrășcoiu, N.; Roșu, C.; Iancu, I. Stațiuni Forestiere; Editura Academiei Republicii Socialiste România: București, Romania, 1977; pp. 20–100, 158–169. [Google Scholar]
- Târziu, D.; Spârchez, G. Soluri și Stațiuni Forestiere; Editura Universității Transilvania: Brașov, Romania, 2013; pp. 104–109. ISBN 978-606-19-0260-6. [Google Scholar]
- Seynave, I.; Gégout, J.C.; Hervé, J.C.; Dhôte, J.F.; Drapier, J.; Bruno, E.; Dumé, G. Picea abies site index prediction by environmental factors and understorey vegetation: A two-scale approach based on survey databases. Can. J. For. Res. 2005, 35, 1669–1678. [Google Scholar] [CrossRef]
- Jensen, J.; Rasmussen, L.; Raulund-Rasmussen, K.; Borggaard, O. Influence of soil properties on the growth of sycamore (Acer pseudoplatanus L.) in Denmark. Eur. J. For. Res. 2008, 127, 263–274. [Google Scholar] [CrossRef]
- Kobal, M.; Grčman, H.; Zupan, M.; Levanič, T.; Simončič, P.; Kadunc, A.; Hladnik, D. Influence of soil properties on silver fir (Abies alba Mill.) growth in the Dinaric Mountains. For. Ecol. Manag. 2015, 337, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Fajardo, A.; Mcintire, E.J.B. Distinguishing microsite and competition processes in tree growth dynamics: An a priori. Am. Nat. 2007, 169, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.D.; Marion, D.A. Biomechanical effects, lithological variations, and local pedodiversity in some forest soils of Arkansas. Geoderma 2005, 124, 73–89. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Bockheim, J.G. Pedodiversity in an old-growth northern hardwood forest in the Huron Mountains, Upper Peninsula, Michigan. Can. J. For. Res. 2007, 37, 1106–1117. [Google Scholar] [CrossRef]
- La Roi, G.H.; Strong, W.L.; Pluth, D.J. Understory plant community classifications as predictors of forest site quality for lodgepole pine and white spruce in west-central Alberta. Can. J. For. Res. 1988, 18, 875–887. [Google Scholar] [CrossRef]
- Strong, W.L.; Pluth, D.J.; La Roi, G.H.; Corns, I.G.W. Forest understory plants as predictors of lodgepole pine and white spruce site quality in west-central Alberta. Can. J. For. Res. 1991, 21, 1675–1683. [Google Scholar] [CrossRef]
- Bergès, L.; Chevalier, R.; Dumas, Y.; Franc, A.; Gilbert, J.M. Sessile oak (Quercus petraea Liebl.) site index variations in relation to climate, topography and soil in even-aged high-forest stands in northern France. Ann. For. Sci. 2005, 62, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Beldie, A.; Chiriță, C. Flora Indicatoare din Pădurile Noastre; Editura Agro-Silvică: București, Romania, 1960; pp. 15–25, 113–141, 159–161. [Google Scholar]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of spatial and temporal variability in natural site conditions. Forestry 2013, 86, 305–315. [Google Scholar] [CrossRef]
- Berrill, J.; O’Hara, K.L. Estimating site productivity in irregular stand structures by indexing the basal area or volume increment of the dominant species. Can. J. For. Res. 2014, 44, 92–100. [Google Scholar] [CrossRef]
- Jiang, H.; Radtke, P.J.; Weiskittel, A.R.; Coulston, J.W.; Guertin, P.J. Climate- and soil-based models of site productivity in eastern US tree species. Can. J. For. Res. 2015, 45, 325–342. [Google Scholar] [CrossRef]
- Del Río, M.; Pretzsch, H.; Alberdi, I.; Bielak, K.; Bravo, F.; Brunner, A.; Condés, S.; Ducey, M.J.; Fonseca, T.; Von Lüpke, N.; et al. Characterization of the structure, dynamics, and productivity of mixed-species stands: Review and perspectives. Eur. J. For. Res. 2016, 135, 23–49. [Google Scholar] [CrossRef]
- Socha, J.; Tyminska-Czabanscka, L.; Grabska, E.; Orzel, S. Site index models for main forest-forming tree species in Poland. Forests 2020, 11, 301. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Sharma, R.P.; Zhu, G.; Li, H.; Hong, L.; Guo, H.; Duan, G.; Shen, C.; Lei, Y.; Li, Y.; et al. Basal Area Increment-Based Approach of Site Productivity Evaluation for Multi-Aged and Mixed Forests. Forests 2017, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Pokharel, B.; Froese, R.E. Representing site productivity in the basal area increment model for FVS-Ontario. For. Ecol. Manag. 2009, 258, 657–666. [Google Scholar] [CrossRef]
- Vanclay, J.K. Assessing site productivity in tropical moist forests. For. Ecol. Manag. 1992, 54, 257–287. [Google Scholar] [CrossRef] [Green Version]
- Tudoran, G.; Zotta, M. Adapting the planning and management of Norway spruce forests in mountain areas of Romania to environmental conditions including climate change. Sci. Total Environ. 2019. [Google Scholar] [CrossRef]
- Marin, D. Amenajamentul U.P. I și II (O.S. Făgăraș), 2018. Amenajamentul U.P. I și II (O.S. Brașov), Romania, 2015. Romanian National Institute for Research and Development in Forestry. Available online: http://www.rosilva.ro/articole/amenajamente_silvice__p_2645.htm (accessed on 26 October 2020).
- Bosela, M.; Lukac, M.; Castagneri, D.; Sedmák, R.; Biber, P.; Carrer, M.; Konôpka, B.; Nola, P.; Nagel, T.A.; Popa, I.; et al. Contrasting effects of environmental change on the radial growth of co-occurring beech and fir trees across Europe. Sci. Total Environ. 2018, 615, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Hilmers, T.; Avdagić, A.; Bartkowicz, L.; Bielak, K.; Binder, F.; Bončina, A.; Dobor, L.; Forrester, D.I.; Hobi, M.L.; Ibrahimspahić, A.; et al. The productivity of mixed mountain forests comprised of Fagus sylvatica, Picea abies, and Abies alba across Europe. Forestry 2019, 92, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Hilmers, T.; Uhl, E.; Bielak, K.; Bosela, M.; del Rio, M.; Dobor, L.; Forrester, D.I.; Nagel, T.A.; Pach, M.; et al. European beech stem diameter grows better in mixed than in mono-specifc stands at the edge of its distribution in mountain forests. Eur. J. For. Res. 2020, 140, 127–145. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef] [Green Version]
- Bosela, M.; Tobin, B.; Šebeň, V.; Petráš, R.; Larocque, G.R. Different mixtures of Norway spruce, silver fir, and European beech modify competitive interactions in central European mature mixed forests. Can. J. For. Res. 2015, 45, 1577–1586. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dauber, E. Long-term stand dynamics of managed spruce–fir–beech mountain forests in Central Europe: Structure, productivity and regeneration success. Forestry 2015, 88, 407–428. [Google Scholar] [CrossRef]
- Strimbu, B.M.; Amarioarei, A.; Paun, M. Nonlinear parsimonious forest modeling assuming normal distribution of residuals. Eur. J. For. Res. 2021. [Google Scholar] [CrossRef]
- Giurgiu, V.; Drăghiciu, D. Modele Matematico-Auxologice și Tabele de Producție Pentru Arborete; Editura Ceres: București, Romania, 2004; pp. 544–623. [Google Scholar]
- Giurgiu, V. Dendrometrie şi Auxologie Forestieră; Editura Ceres: București, Romania, 1979; pp. 126–568. [Google Scholar]
- Kerr, G. Effects of spacing on the early growth of planted Fraxinus excelsior L. Can. J. For. Res. 2003, 33, 1196–1207. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation Variables | Units | Description |
|---|---|---|
| hdom | m | Average height of dominant trees |
| hg | m | Height of tree with dg (height of mean-basal-area tree) |
| fhg or FH | – | Form height of mean-basal-area tree () or form height of stand () |
| V | m3ha−1 | Volume of stand |
| G | m2ha−1 | Basal area of stand |
| Rhgg | – | Height to basal area ratio (hg/gg) |
| v | m3 | Volume of tree |
| CAI | m3ha−1year−1 | Current annual volume increment |
| MAI | m3ha−1year−1 | Mean annual volume increment |
| T | years | Stand age (or age of mean-basal-area tree) |
| dg | cm | Mean squared diameter or quadratic mean diameter (diameter corresponding to mean basal area of stand or diameter of the mean-basal-area tree) |
| vg | m3 | Volume of mean-basal-area tree |
| g | m2 | Basal area of tree (g or ba) |
| gg | m2 | Mean basal area of stand () |
| Parameters | Classes | Total | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Altitude (m) | 800–1000 | 1001–1200 | 1201–1400 | 1401–1600 | – | |||||||||||||
| Area (%) | 60 | 15 | 21 | 4 | 100 | |||||||||||||
| Species | European beech | Silver fir | Norway spruce | Deciduous species | – | |||||||||||||
| Area (%) | 42 | 11 | 44 | 3 | 100 | |||||||||||||
| Density | 0.1–0.2 | 0.3–0.4 | 0.5–0.6 | 0.7–0.8 | 0.9–1.0 | – | ||||||||||||
| Area (%) | – | 2 | 5 | 77 | 16 | 0.78 | ||||||||||||
| Exposure | Shaded (N, NE) | Sunny (S, SW) | Semi-shaded (E, SE, W, NW) | – | ||||||||||||||
| Area (%) | 11 | 49 | 40 | 100 | ||||||||||||||
| Inclination (degrees) | <15 | 16–30 | 31–40 | >40 | – | |||||||||||||
| Area (%) | 6 | 44 | 49 | 1 | 100 | |||||||||||||
| Age class (20 years) | I | II | III | IV | V | VI | VII | – | ||||||||||
| Area | ha | 336.16 | 613.81 | 1167.05 | 1475.32 | 1874.01 | 1456.44 | 2185.00 | 9107.79 | |||||||||
| % | 4 | 7 | 13 | 16 | 20 | 16 | 24 | 100 | ||||||||||
| Model | Variable | Species | Site | Equation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| y | x | Symbol | Level Quality | Number | a | b | c | d | ||
| 1 | hdom (m) | Age (years) | Spruce | 3.3.3.3 | High | (2) | 1.0323 | 0.5606 | −0.0024 | |
| hg (m) | Age (years) | Spruce | 3.3.3.3 | High | (8) | −1.2350 | 0.5693 | −0.0024 | – | |
| hg (m) | dg (cm) | Fir | 3.3.3.3 | High | (14) | −0.6348 | 1.0646 | −0.0086 | – | |
| fhg (m) | Age (years) | Beech | 3.3.3.1 | Low | (31) | 0.3495 | 0.1725 | −0.0006 | – | |
| fhg (m) | dg (cm) | Fir | 3.3.3.2 | Medium | (24) | −0.9849 | 0.5511 | −0.0057 | – | |
| vg (m3) | gg (m2) | Spruce | 3.3.3.3 | High | (62) | −0.1737 | 14.7945 | −5.6659 | – | |
| 2 | hg/gg | Age (years) | Beech | 3.3.3.2 | Medium | (48) | 1137.369 | –0.0157 | – | – |
| hg/gg | dg (cm) | Beech | 3.3.3.3 | High | (38) | 1462.454 | –0.0452 | – | – | |
| 3 | CAI (m3year−1ha−1) | Age (years) | Beech | 3.3.3.1 | Low | (67) | −1.3032 | 0.3843 | −0.0050 | 1.776 × 10−5 |
| MAI (m3year−1ha−1) | Age (years) | Fir | 3.3.3.2 | Medium | (77) | 0.4259 | 0.1785 | −0.0015 | 3.165 × 10−6 | |
| Model | Species | Variable | Site | Equation | R2 | R2 Adjusted | RMSE | MAE | MAPE(%) | AIC | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| y | x | Symbol | Level Quality | Number/(p) | a | b | c | d | ||||||||
| 1 | Fir | hdom (m) | Age (years) | 3.3.3.3 | High | (5) | 6.034 | 0.406 | 307 | – | 0.947 | 0.946 | 1.123 | 0.912 | 307 | 1.123 |
| p | 3.32 × 10−7 | 8.28 × 10−26 | 1.11 × 10−11 | |||||||||||||
| Spruce | hdom (m) | Age (years) | 3.3.3.2 | Medium | (9) | −3.562 | 0.624 | 1581 | – | 0.967 | 0.967 | 1.259 | 1.026 | 1581 | 1.259 | |
| p | 2.20 × 10−24 | 4.29 × 10−225 | 3.25 × 10−147 | |||||||||||||
| Beech | hg (m) | Age (years) | 3.3.3.3 | High | (11) | −1.7057 | 0.5316 | 4.05 | – | 0.977 | 0.977 | 1.072 | 0.825 | 4.05 | 734 | |
| p | 8.24 × 10−5 | 9.63 × 10−129 | 1.18 × 10−82 | |||||||||||||
| Fir | hg (m) | Age (years) | 3.3.3.2 | Medium | (15) | −2.7347 | 0.4649 | 4.88 | – | 0.976 | 0.975 | 1.13 | 0.932 | 4.88 | 281 | |
| p | 1.95 × 10−5 | 2.60 × 10−40 | 5.20 × 10−21 | |||||||||||||
| Spruce | hg (m) | Age (years) | 3.3.3.2 | Medium | (18) | −4.0748 | 0.5462 | 6.348 | – | 0.964 | 0.964 | 1.294 | 1.053 | 6.348 | 1566 | |
| p | 9.74 × 10−34 | 1.03 × 10−203 | 1.87 ×10−121 | |||||||||||||
| Spruce | hg (m) | dg (cm) | 3.3.3.3 | High | (26) | −2.0272 | 1.1912 | 3.752 | – | 0.944 | 0.944 | 1.327 | 1.014 | 3.752 | 934 | |
| p | 0.0003 | 3.93 × 10−93 | 1.46 × 10−60 | |||||||||||||
| Fir | fhg (m) | dg (cm) | 3.3.3.2 | Medium | (33) | −0.9849 | 0.5511 | 6.202 | – | 0.953 | 0.953 | 0.643 | 0.522 | 6.202 | 334 | |
| p | 8.34 × 10−10 | 2.80 × 10−62 | 5.18 × 10−39 | |||||||||||||
| Beech | fhg (m) | dg (cm) | 3.3.3.2 | Medium | (39) | −0.6427 | 0.2273 | 5.134 | – | 0.932 | 0.932 | 0.679 | 0.565 | 5.134 | 1307 | |
| p | 0.0004 | 4.91 × 10−212 | 1.40 × 10−118 | |||||||||||||
| Fir | fhg (m) | Age (years) | 3.3.3.2 | Medium | (42) | −1.1826 | 0.24 | 5.276 | – | 0.970 | 0.969 | 0.595 | 0.470 | 5.276 | 149 | |
| p | 0.0002 | 7.55 × 10−43 | 1.40 × 10−27 | |||||||||||||
| Spruce | fhg (m) | Age (years) | 3.3.3.2 | Medium | (45) | −1.1854 | 0.2537 | 5.83 | – | 0.948 | 0.948 | 0.559 | 0.455 | 5.83 | 695 | |
| p | 5.50 × 10−12 | 1.06 × 10−154 | 6.99 × 10−98 | |||||||||||||
| Beech | vg (m3) | gg (m2) | 3.3.3.3 | High | (65) | −0.0895 | 12.875 | 4.472 | – | 0.995 | 0.995 | 0.07 | 0.053 | 4.472 | −658 | |
| p | 5.27 × 10−14 | 7.51 × 10−108 | 4.96 × 10−14 | |||||||||||||
| Beech | vg (m3) | gg (m2) | 3.3.3.2 | Medium | (66) | −0.0756 | 11.9478 | 4.404 | – | 0.989 | 0.989 | 0.083 | 0.064 | 4.404 | −1368 | |
| p | 2.80 × 10−11 | 8.92 × 10−162 | 7.79 × 10−16 | |||||||||||||
| 3 | Beech | CAI (m3year−1ha−1) | Age (years) | 3.3.3.2 | Medium | (75) | −2.2068 | 0.5508 | 9.123 | 2.775 × 10−5 | 0.844 | 0.843 | 0.807 | 0.597 | 9.123 | 2078 |
| p | 2.41 × 10−5 | 8.98 × 10−107 | 6.52 × 10−98 | 3.48 × 10−74 | ||||||||||||
| Fir | CAI (m3year−1ha−1) | Age (years) | 3.3.3.2 | Medium | (78) | −4.288 | 0.7921 | 12.82 | 4.006 × 10−5 | 0.811 | 0.807 | 1.317 | 1.001 | 12.82 | 478 | |
| p | 9.08 × 10−7 | 7.51 × 10−25 | 8.24 × 10−22 | 7.51 × 10−17 | ||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cicșa, A.; Tudoran, G.-M.; Boroeanu, M.; Dobre, A.-C.; Spârchez, G. Estimation of the Productivity Potential of Mountain Sites (Mixed Beech-Coniferous Stands) in the Romanian Carpathians. Forests 2021, 12, 549. https://doi.org/10.3390/f12050549
Cicșa A, Tudoran G-M, Boroeanu M, Dobre A-C, Spârchez G. Estimation of the Productivity Potential of Mountain Sites (Mixed Beech-Coniferous Stands) in the Romanian Carpathians. Forests. 2021; 12(5):549. https://doi.org/10.3390/f12050549
Chicago/Turabian StyleCicșa, Avram, Gheorghe-Marian Tudoran, Maria Boroeanu, Alexandru-Claudiu Dobre, and Gheorghe Spârchez. 2021. "Estimation of the Productivity Potential of Mountain Sites (Mixed Beech-Coniferous Stands) in the Romanian Carpathians" Forests 12, no. 5: 549. https://doi.org/10.3390/f12050549
APA StyleCicșa, A., Tudoran, G. -M., Boroeanu, M., Dobre, A. -C., & Spârchez, G. (2021). Estimation of the Productivity Potential of Mountain Sites (Mixed Beech-Coniferous Stands) in the Romanian Carpathians. Forests, 12(5), 549. https://doi.org/10.3390/f12050549