
Chitosan Oligosaccharides Stimulate the Efficacy of Somatic Embryogenesis in Different Genotypes of the Liriodendron Hybrid

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Initiation
2.2. Configuration of COS Stock Solution
2.3. Concentration of COS
2.4. In Vitro Growth of Somatic Embryo Seedlings
2.5. Ex Vitro Adaptation of Somatic Embryo Seedlings
2.6. Somatic Embryo Observation
2.7. Acquisition and Growth Potential Measurement of Regenerated Plants
2.8. Seedling Effects of COS
2.9. Analysis of Endogenous Hormone Levels
3. Data Analysis
4. Results
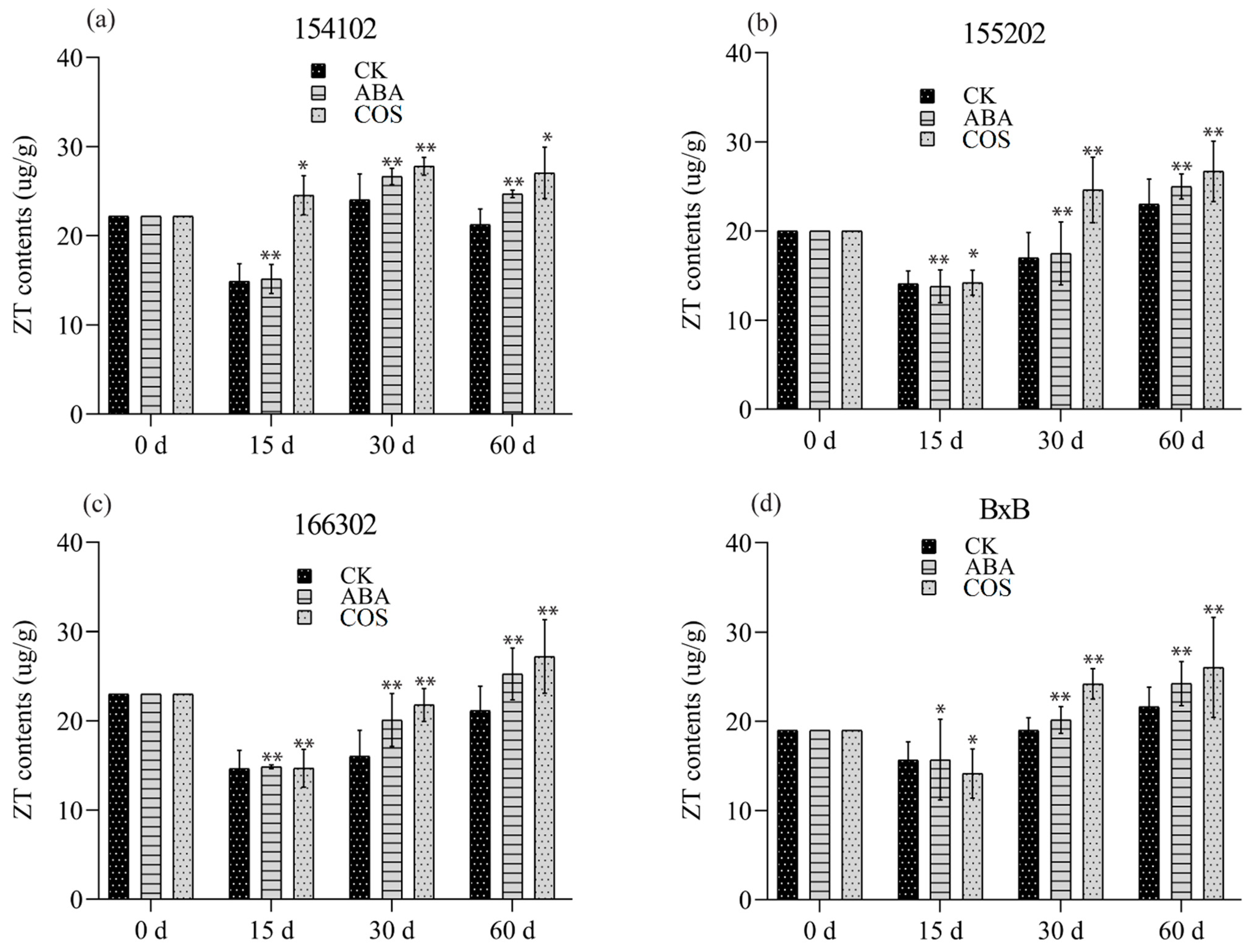
4.1. COS Regulating the Level of Endogenous Hormones
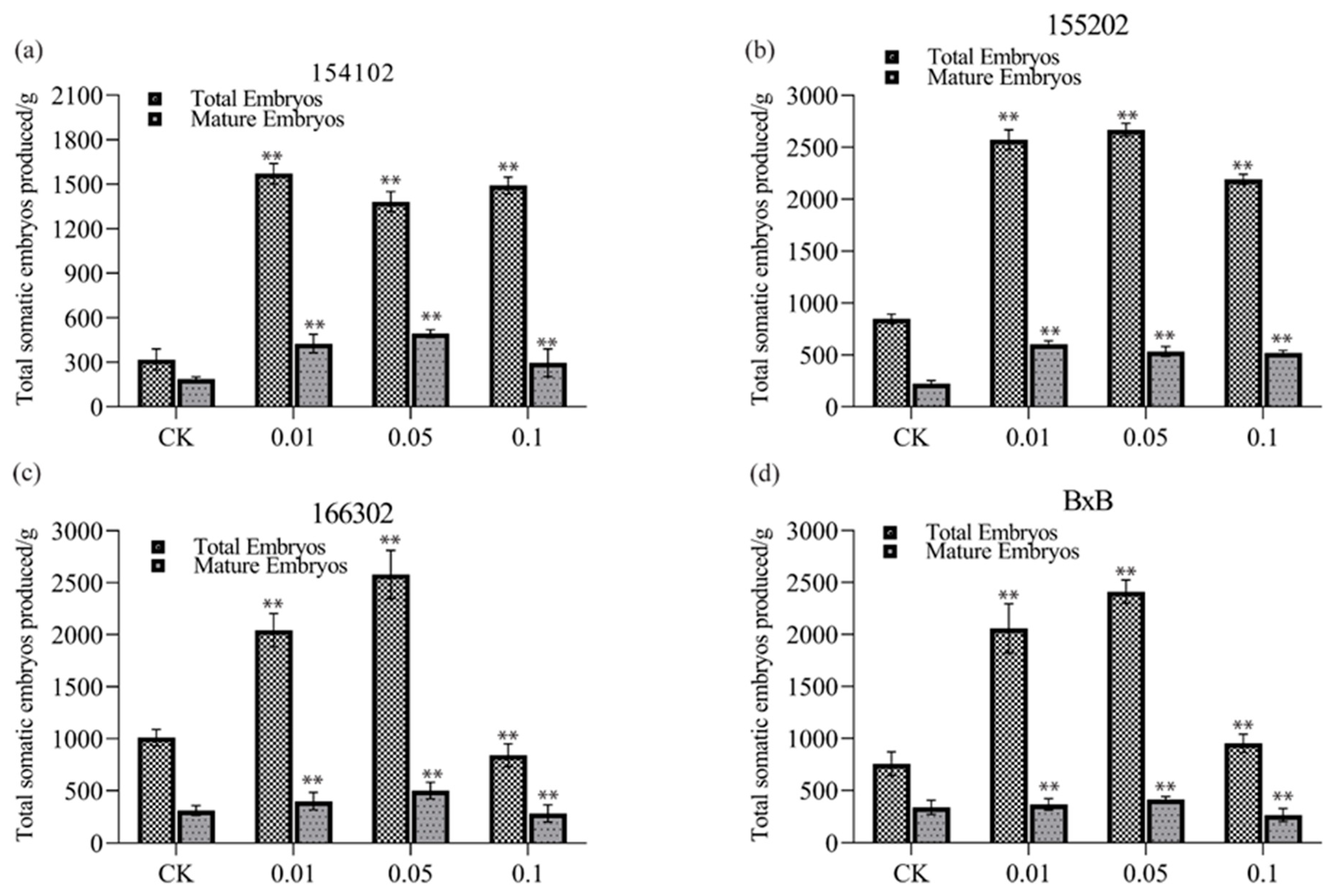
4.2. Germination Ability
4.3. Acquisition and Growth Potential Measurement of Somatic Embryo Regenerated Plants
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Hao, Z.; Guang, X.; Zhao, C.; Wang, P.; Xue, L.; Zhu, Q.; Yang, L.; Sheng, Y.; Zhou, Y.; et al. Liriodendron genome sheds light on angiosperm phylogeny and species–pair differentiation. Nat. Plants 2019, 5, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Parks, C.R.; Wendel, J.F. Molecular divergence between Asian and North American species of Liriodendron (Magnoliaceae) with implications for interpretation of fossil floras. Am. J. Bot. 1990, 77, 1243–1256. [Google Scholar] [CrossRef]
- Zhen, Y.; Li, C.; Chen, J.; Chen, Q.; Shi, J. Proteomics of embryogenic and non-embryogenic calli of a liriodendron hybrid. Acta Physiol. Plant. 2015, 37, 211. [Google Scholar] [CrossRef]
- Shang, C.; Wang, Z. A new scientific name of hybrid Liriodendron-L. Sino-americanum. J. Nanjing For. Univ. 2012, 36, 1–2. [Google Scholar]
- Cheng, Y.; Li, H. Interspecies evolutionary divergence in Liriodendron, evidence from the nucleotide variations of the lcdhn-like gene. BMC Evol. Biol. 2018, 18, 195. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Xu, M.; Luo, Q.; Wang, J.; Li, H. De novo transcriptome analysis of Liriodendron chinense petals and leaves by Illumina sequencing. Gene 2014, 534, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Rudall, P. Comparative floral anatomy and ontogeny in Magnoliaceae. Plant Syst. Evol. 2006, 258, 1–15. [Google Scholar] [CrossRef]
- Endress, P.K. The evolution of floral biology in basal angiosperms. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 411–421. [Google Scholar] [CrossRef]
- Li, M.; Wang, K.; Wang, X.; Yang, P. Morphological and proteomic analyses reveal the role of pistil under pollination in Liriodendron chinense (hemsl.) Sarg. PLoS ONE 2014, 9, e99970. [Google Scholar]
- Li, B.; Li, Y.; Cai, Q.; Lin, F.; Meng, Q.; Zheng, Y. The complete chloroplast genome of a Tertiary relict species Liriodendron chinense (Magnoliaceae). Conserv. Genet. Resour. 2016, 8, 279–281. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. The origin and diversification of angiosperms. Am. J. Bot. 2004, 91, 1614–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Z.-L.; Wen, J.; Azuma, H.; Qiu, Y.-L.; Sun, H.; Meng, Y.; Sun, W.-B.; Zimmer, E.A. Phylogenetic and biogeographic complexity of Magnoliaceae in the Northern Hemisphere inferred from three nuclear data sets. Mol. Phylogenetics Evol. 2008, 48, 1027–1040. [Google Scholar] [CrossRef] [PubMed]
- Hunt, D. Magnolias and Their Allies; Published for the International Dendrology Society and the Magnolia Society: London, UK, 1998. [Google Scholar]
- Moody, R.C. Yellow Popular Glulam Timber Beam Performance; U.S. Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 1993; Volume 520.
- Hernandez, R. Strength and Stiffness of Reinforced Yellow-Poplar Glued-Laminated Beams; U.S. Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 1997; Volume 554.
- Williams, R.S.; Feist, W.C. Durability of yellow-poplar and Sweetgum and service life of finishes after long-term exposure. For. Prod. J. 2004, 54, 96–101. [Google Scholar]
- Kim, K.D.; Lee, E.J. Potential Tree Species for Use in the Restoration of Unsanitary Landfills. Environ. Manag. 2005, 36, 1–14. [Google Scholar] [CrossRef]
- Moon, M.K.; Oh, H.M.; Kwon, B.-M.; Baek, N.-L.; Kim, S.-H.; Kim, J.S.; Kim, D.K. Farnesyl protein transferase and tumor cell growth inhibitory activities of lipiferolide isolated from Liriodendron tulipifera. Arch. Pharmacal Res. 2007, 30, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Kudi, A.; Umoh, J.; Eduvie, L.; Gefu, J. Screening of some Nigerian medicinal plants for antibacterial activity. J. Ethnopharmacol. 1999, 67, 225–228. [Google Scholar] [CrossRef]
- Celen, I.; Harper, D.; Labbe, N. A multivariate approach to the acetylated poplar wood samples by near-infrared spectroscopy. Holzforschung 2008, 62, 189–196. [Google Scholar] [CrossRef]
- Xiang, Q.; Lee, Y.Y.; Torget, R.W. Kinetics of glucose decomposition during dilute-acid hydrolysis of lignocellulosic biomass. In Proceedings of the Twenty-Fifth Symposium on Biotechnology for Fuels and Chemicals Held, Breckenridge, CO, USA, 4–7 May 2003; Springer: Berlin/Heidelberg, Germany, 2004; pp. 1127–1138. [Google Scholar]
- Berlin, A.; Maximenko, V.; Bura, R.; Kang, K.-Y.; Gilkes, N.; Saddler, J. A rapid microassay to evaluate enzymatic hydrolysis of lignocellulosic substrates. Biotechnol. Bioeng. 2006, 93, 880–886. [Google Scholar] [CrossRef]
- Ri-Ming, H.; Shan-An, H.; Shi-Jie, T.; Shou-Peng, W. Geographical distribution of Liriodendron chinense in china and its significance. J. Plant Resour. Environ. 1995, 1, 1–6. [Google Scholar]
- Fu, L.; Jin, J. Red List of Endangered Plants in China; Science Press: Beijing, China, 1992. [Google Scholar]
- Guan, Y.; Li, S.-G.; Fan, X.-F.; Su, Z.-H. Application of Somatic Embryogenesis in Woody Plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Wei, W.; Zhou, W.; McGuigan, L.D.; Ji, F.-Y.; Li, X.; Xing, Y.; Zhang, Q.; Fang, K.-F.; Cao, Q.-Q.; et al. Establishment of a somatic embryo regeneration system and expression analysis of somatic embryogenesis-related genes in Chinese chestnut (Castanea mollissima Blume). Plant Cell Tissue Organ Cult. 2017, 130, 601–616. [Google Scholar] [CrossRef] [Green Version]
- Khilwani, B.; Kaur, A.; Ranjan, R.; Kumar, A. Direct somatic embryogenesis and encapsulation of somatic embryos for in vitro conservation of Bacopa monnieri (L.) Wettst. Plant Cell Tissue Organ Cult. 2016, 127, 433–442. [Google Scholar] [CrossRef]
- Guillou, C.; Fillodeau, A.; Brulard, E.; Breton, D.; Maraschin, S.D.F.; Verdier, D.; Simon, M.; Ducos, J.-P. Indirect somatic embryogenesis of Theobroma cacao L. in liquid medium and improvement of embryo-to-plantlet conversion rate. Vitr. Cell. Dev. Biol. Anim. 2018, 54, 377–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, S.M.; Ghosh, B. High-frequency somatic embryogenesis and artificial seeds for mass production of true-to-type plants in Ledebouria revoluta: An important cardioprotective plant. Plant Cell Tissue Organ Cult. 2016, 127, 71–83. [Google Scholar] [CrossRef]
- Merkle, S.A.; Dean, J.F. Forest tree biotechnology. Curr. Opin. Biotechnol. 2000, 11, 298–302. [Google Scholar] [CrossRef]
- Chiancone, B.; Germanà, M.A. Micropropagation of Citrus spp. by Organogenesis and Somatic Embryogenesis. In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 99–118. [Google Scholar]
- Ozudogru, E.A.; Lambardi, M. Cryotechniques for the long-term conservation of embryogenic cultures from woody plants. In Vitro Embryogenesis in Higher Plants; Springer: Berlin/Heidelberg, Germany, 2016; pp. 537–550. [Google Scholar]
- Rugh, C.L.; Senecoff, J.F.; Meagher, R.B.; Merkle, S.A. Development of transgenic yellow poplar for mercury phytoremediation. Nat. Biotechnol. 1998, 16, 925–928. [Google Scholar] [CrossRef]
- Vidal, N.; Mallón, R.; Valladares, S.; Meijomín, A.; Vieitez, A. Regeneration of transgenic plants by agrobacterium-mediated transformation of somatic embryos of juvenile and mature Quercus robur. Plant Cell Rep. 2010, 29, 1411–1422. [Google Scholar] [CrossRef]
- Cabrera, J.C.; Wégria, G.; Onderwater, R.C.A.; González, G.; Nápoles, M.C.; Falcón-Rodríguez, A.B.; Costales, D.; Rogers, H.J.; Diosdado, E.; González, S.; et al. Practical use of oligosaccharins in agriculture. Acta Hortic. 2013, 1009, 195–212. [Google Scholar] [CrossRef]
- Kashyap, P.L.; Xiang, X.; Heiden, P. Chitosan nanoparticle-based delivery systems for sustainable agriculture. Int. J. Biol. Macromol. 2015, 77, 36–51. [Google Scholar] [CrossRef]
- Cadano, J.R.; Jose, M.; Lubi, A.G.; Maling, J.N.; Moraga, J.S.; Shi, Q.Y.; Vegafria, H.M.; VinceCruz-Abeledo, C.C. A comparative study on the raw chitin and chitosan yields of common bio-waste from philippine seafood. Environ. Sci. Pollut. Res. 2021, 28, 11954–11961. [Google Scholar] [CrossRef]
- Fadlaoui, S.; Asri, O.E.; Mohammed, L.; Sihame, A.; Melhaoui, M. Isolation and characterization of chitin from shells of the freshwater crab Potamon algeriense. Prog. Chem. Appl. Chitin Its Deriv. 2019, 14, 23–35. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Trouvelot, S.; Héloir, M.-C.; Poinssot, B.; Gauthier, A.; Paris, F. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albersheim, P.; Darvill, A.G.; McNeil, M.; Valent, B.S.; Sharp, J.K.; Nothnagel, E.A.; Davis, K.R.; Yamazaki, N.; Gollin, D.J.; York, W.S.; et al. Oligosaccharins: Naturally occurring carbohydrates with biological regulatory functions. In Structure and Function of Plant Genomes; Springer: Berlin/Heidelberg, Germany, 1983; pp. 293–312. [Google Scholar]
- Enríquez-Guevara, E.A.; Aispuro-Hernández, E.; Vargas-Arispuro, I.; Martínez-Téllez, M.Á. Cell wall oligosaccharine derivatives: Biological activity and participation in the response of plant defense. Rev. Mex. Fitopatol. 2010, 28, 144–155. [Google Scholar]
- López-Guerrero, A.G.; Rodríguez-Hernández, A.M.; Mounzer, O.; Zenteno-Savín, T.; Rivera-Cabrera, F.; Izquierdo-Oviedo, H.; Soriano-Melgar, L.d.A.A. Effect of oligosaccharins on the vase life of lisianthus (Eustoma grandiflorum raf.) cv. ‘Mariachi blue’. J. Hortic. Sci. Biotechnol. 2019, 95, 316–324. [Google Scholar] [CrossRef]
- Cté, F.; Hahn, M.G. Oligosaccharins: Structures and signal transduction. Plant Mol. Biol. 1994, 26, 1379–1411. [Google Scholar]
- Meng, Q.-Y.; Wang, H.; Cui, Z.-B.; Yu, W.-G.; Lu, X.-Z. Chitosan oligosaccharides attenuate amyloid formation of hiapp and protect pancreatic β-cells from cytotoxicity. Molecules 2020, 25, 1314. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Zheng, J.; Jiao, S.; Cheng, G.; Feng, C.; Du, Y.; Liu, H. A review on the preparation of chitosan oligosaccharides and application to human health, animal husbandry and agricultural production. Carbohydr. Polym. 2019, 220, 60–70. [Google Scholar] [CrossRef]
- Salachna, P.; Grzeszczuk, M.; Soból, M. Effects of chitooligosaccharide coating combined with selected ionic polymers on the stimulation of Ornithogalum saundersiae growth. Molecules 2017, 22, 1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, G.; Azad, M.; Kalam, A.; Lin, Y.; Kim, S.W.; Tian, Y.; Liu, G.; Wang, H. Biological Effects and Applications of Chitosan and Chito-Oligosaccharides. Front. Physiol. 2019, 10, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Necha, L.L.B.; Bautista-Baños, S. Chapter 8–prospects for the use of chitosan and other alternatives in ornamental conservation. Chitosan Preserv. Agric. Commod. 2016, 221–249. [Google Scholar]
- Dumville, J.C.; Fry, S.C. Solubilisation of tomato fruit pectins by ascorbate: A possible non-enzymic mechanism of fruit softening. Planta 2003, 217, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, Y.; Zhang, R.; Wang, W.; Zhao, X.; Du, Y.; Yin, H. Effects of chitosan oligosaccharides on the yield components and production quality of different wheat cultivars (Triticum aestivum L.) in northwest China. Field Crop. Res. 2015, 172, 11–20. [Google Scholar] [CrossRef]
- He, Y.; Santosh, B.; Wang, W.; Jia, X.; Lu, H.; Yin, H. Pre-harvest treatment of chitosan oligosaccharides improved strawberry fruit quality. Int. J. Mol. Sci. 2018, 19, 2194. [Google Scholar] [CrossRef] [Green Version]
- Lodhi, G.; Kim, Y.S.; Hwang, J.W.; Kim, S.K.; Jeon, Y.J.; Je, J.Y.; Ahn, C.B.; Moon, S.H.; Jeon, B.T.; Park, P.J. Chitooligosaccharide and its derivatives: Preparation and biological applications. BioMed Res. Int. 2014, 2014, 654913. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Castro, J.; Vera, J.; Moenne, A. Seaweed oligosaccharides stimulate plant growth by enhancing carbon and nitrogen assimilation, basal metabolism, and cell division. J. Plant Growth Regul. 2013, 32, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Malerba, M.; Cerana, R. Chitosan Effects on Plant Systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trends Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Reactive Oxygen and Nitrogen Species in Defense/Stress Responses Activated by Chitosan in Sycamore Cultured Cells. Int. J. Mol. Sci. 2015, 16, 3019–3034. [Google Scholar] [CrossRef] [Green Version]
- Hirano, S.; Yamamoto, T.; Hayashi, M.; Nishida, T.; Inui, H. Chitinase activity in [several plants] seeds coated with chitosan derivatives. Agric. Biol. Chem. 2014, 54, 2719–2720. [Google Scholar]
- Choi, C.; Nam, J.-P.; Nah, J.-W. Application of chitosan and chitosan derivatives as biomaterials. J. Ind. Eng. Chem. 2016, 33, 1–10. [Google Scholar] [CrossRef]
- Guo, W.; Ye, Z.; Wang, G.; Zhao, X.; Yuan, J.; Du, Y. Measurement of oligochitosan–tobacco cell interaction by fluorometric method using europium complexes as fluorescence probes. Talanta 2009, 78, 977–982. [Google Scholar] [CrossRef]
- Guo, W.; Yin, H.; Ye, Z.; Zhao, X.; Yuan, J.; Du, Y. A comparison study on the interactions of two oligosaccharides with tobacco cells by time-resolved fluorometric method. Carbohydr. Polym. 2012, 90, 491–495. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.; Zhao, X.; Wang, W.; Du, Y.; He, A.; Sun, K. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr. Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef]
- Wan, C.; Wang, M.; Yang, D.; Han, X.; Che, C.; Ding, S.; Xiao, Y.; Qin, Z. Synthesis and biological activity of 2′, 3′-iso-aryl-abscisic acid analogs. Molecules 2017, 22, 2229. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Du, Y.; Dong, Z. Chitin oligosaccharide and chitosan oligosaccharide: Two similar but different plant elicitors. Front. Plant Sci. 2016, 7, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, W.-J.; Park, R.-D. Bioproduction of Chitooligosaccharides: Present and Perspectives. Mar. Drugs 2014, 12, 5328–5356. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Yamaguchi, H.; Yuasa, T.; Iwaya-Inoue, M.; Arima, S.; Zheng, S.-H. Hydrogen peroxide spraying alleviates drought stress in soybean plants. J. Plant Physiol. 2011, 168, 1562–1567. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jiang, X.; Chen, X.S. Inhibitory effects of chitosan on superoxide anion radicals and lipid free radicals. Chin. Sci. Bull. 2002, 47, 887–889. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Medium No. | Components of Culture Medium |
|---|---|
| 1 | 3/4 MS + VC5 mg L−1 + Sucrose30 g L−1 + ABA 2 mg L−1 |
| 2 | 3/4 MS + VC5 mg L−1 + Sucrose30 g L−1 + COS 0.01 mg L−1 |
| 3 | 3/4 MS + VC5 mg L−1 + Sucrose30 g L−1 + COS 0.05 mg L−1 |
| 4 | 3/4 MS + VC5 mg L−1 + Sucrose30 g L−1 + COS 0. 1 mg L−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Zhang, J.; Zhou, M.; Chen, T.; Shah, L.; Rehman, S.u.; Hayat, S.; Shi, J.; Chen, J. Chitosan Oligosaccharides Stimulate the Efficacy of Somatic Embryogenesis in Different Genotypes of the Liriodendron Hybrid. Forests 2021, 12, 557. https://doi.org/10.3390/f12050557
Ali A, Zhang J, Zhou M, Chen T, Shah L, Rehman Su, Hayat S, Shi J, Chen J. Chitosan Oligosaccharides Stimulate the Efficacy of Somatic Embryogenesis in Different Genotypes of the Liriodendron Hybrid. Forests. 2021; 12(5):557. https://doi.org/10.3390/f12050557
Chicago/Turabian StyleAli, Asif, Jiaji Zhang, Minmin Zhou, Tingting Chen, Liaqat Shah, Shams ur Rehman, Sikandar Hayat, Jisen Shi, and Jinhui Chen. 2021. "Chitosan Oligosaccharides Stimulate the Efficacy of Somatic Embryogenesis in Different Genotypes of the Liriodendron Hybrid" Forests 12, no. 5: 557. https://doi.org/10.3390/f12050557
APA StyleAli, A., Zhang, J., Zhou, M., Chen, T., Shah, L., Rehman, S. u., Hayat, S., Shi, J., & Chen, J. (2021). Chitosan Oligosaccharides Stimulate the Efficacy of Somatic Embryogenesis in Different Genotypes of the Liriodendron Hybrid. Forests, 12(5), 557. https://doi.org/10.3390/f12050557