
Comparative Efficacy Trials with Two Different Bacillus thuringiensis Serovar kurstaki Strains against Gypsy Moth in Mediterranean Cork Oak Forests




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Products
2.2. Study Area and Experimental Design
2.3. Evaluation of Btk Treatments
2.4. Statistical Analysis
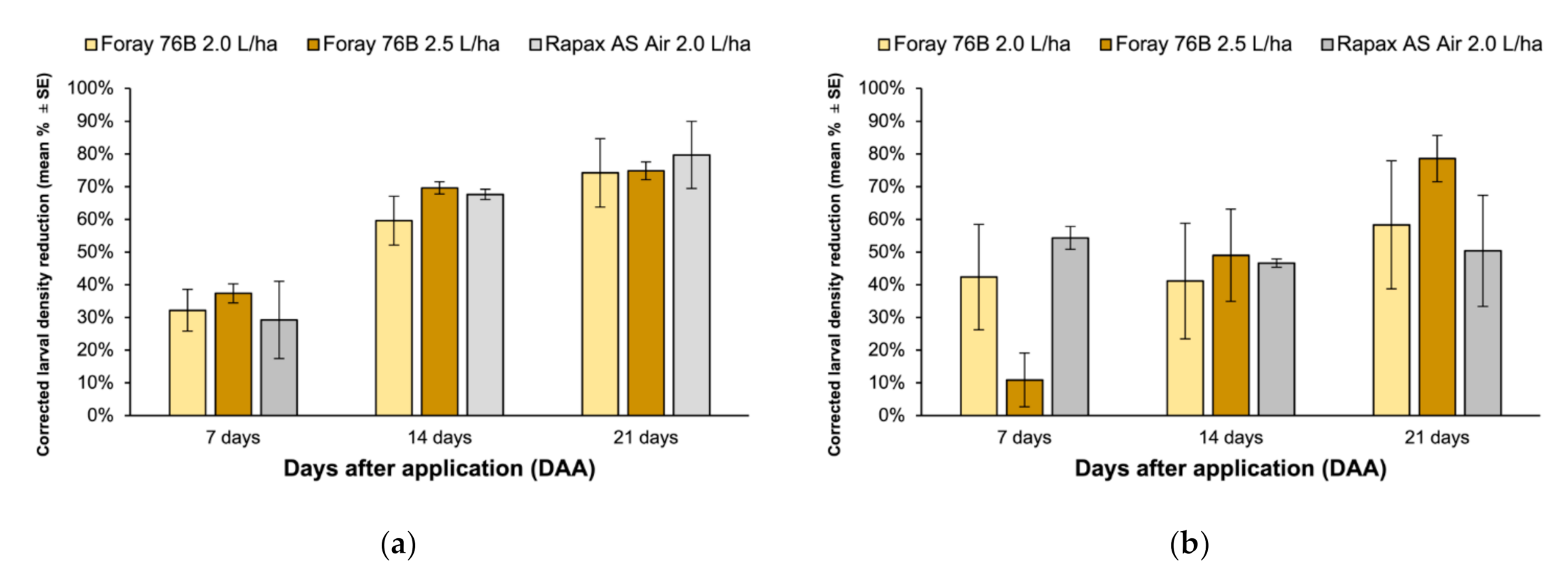
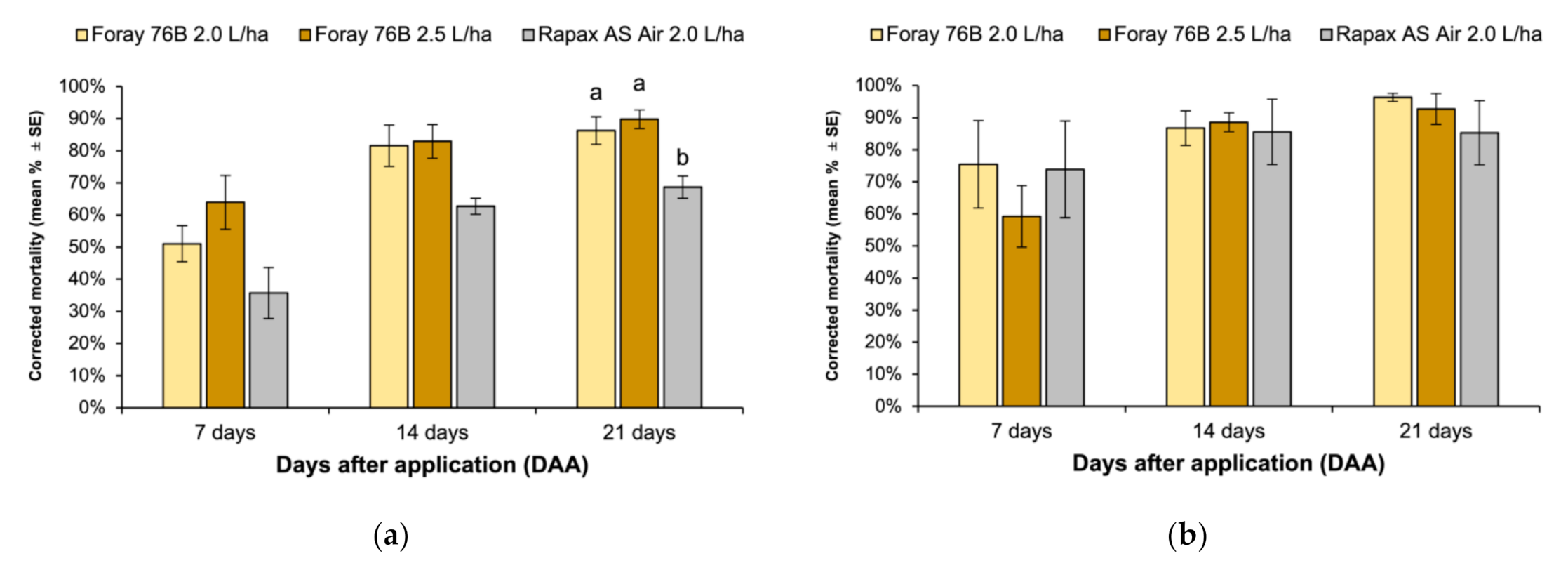
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aronson, J.; Pereira, J.S.; Pausas, J.G. Cork Oak Woodlands on the Edge: Ecology, Adaptative Management, and Restoration; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Verdinelli, M.; Yakhlef, S.E.B.; Cossu, C.S.; Pilia, O.; Mannu, R. Variability of ant community composition in cork oak woodlands across the Mediterranean region: Implication for forest management. IForest 2017, 10, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Mannu, R.; Pilia, O.; Fadda, M.L.; Verdinelli, M. Variability of beetle assemblages in Mediterranean cork oak woodlands: Does the higher taxa approach reliably characterize a specific response to grazing? Biodivers. Conserv. 2018, 27, 3599–3619. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Liebhold, A.M. Population dynamics of gypsy moth in North America. Annu. Rev. Entomol. 1990, 35, 571–596. [Google Scholar] [CrossRef]
- Luciano, P.; Prota, R. Insect pests in Sardinian cork-oak forests. IOBC/WPRS Bull 1995, 18, 1–7. [Google Scholar]
- Davidson, C.B.; Gottschalk, K.W.; Johnson, J.E. Tree mortality following defoliation by the European gypsy moth (Lymantria dispar L.) in the United States: A review. For. Sci. 1999, 45, 74–84. [Google Scholar] [CrossRef]
- Barbosa, P.; Martinat, P.; Waldvogel, M. Development, fecundity and survival of the herbivore Lymantria dispar and the number of plant species in its diet. Ecol. Entomol. 1986, 11, 1–6. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Gottschalk, K.W.; Muzika, R.M.; Montgomery, M.E.; Young, R.; O’Day, K.; Kelley, B. Suitability of North American Tree Species to the Gypsy Moth: A Summary of Field and Laboratory Tests; General Technical Report NE-211; USDA Forest Service: Radnor, PA, USA, 1995. [Google Scholar]
- Floris, I.; Cocco, A.; Buffa, F.; Mannu, R.; Satta, A. Insect pests of Eucalyptus plantations in Sardinia (Italy). Redia 2018, 101, 61–71. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Gottschalk, K.W.; Mason, D.A.; Bush, R.R. Forest susceptibility to the gypsy moth. J. For. 1997, 95, 20–24. [Google Scholar]
- Tiberi, R.; Branco, M.; Bracalini, M.; Croci, F.; Panzavolta, T. Cork oak pests: A review of insect damage and management. Ann. For. Sci. 2016, 73, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Cambini, A. Valutazione Dei Danni Causati Dagli Insetti Defogliatori Alla Quercia Da Sughero. In Proceedings of Atti del 1o Convegno Regionale del Sughero; Stazione Sperimentale del sughero: Tempio Pausania, Italy, 1971; pp. 327–339. [Google Scholar]
- Cambini, A. Effetti Delle Defogliazioni Singole e Duplici Sull’accrescimento Della Quercia da Sughero Quercus Suber L. Memoria 42; Stazione Sperimentale del Sughero: Tempio Pausania, Italy, 1975. [Google Scholar]
- Smitley, D.R.; Davis, T.W. Aerial application of Bacillus thuringiensis for suppression of gypsy moth (Lepidoptera: Lymantriidae) in Populus–Quercus forests. J. Econ. Entomol 1993, 86, 1178–1184. [Google Scholar] [CrossRef]
- Webb, R.E.; Peiffer, R.; Fuester, R.W.; Thorpe, K.W.; Calabrese, L.; McLaughlin, J.M. An evaluation of the residual activity of traditional, safe, and biological insecticides against the gypsy moth. J. Arboric. 1998, 24, 286–293. [Google Scholar]
- Luciano, P.; Lentini, A. Ten years of microbiological control program against lepidopterous defoliators in Sardinian cork oak forests. IOBC/WPRS Bull 2012, 76, 175–178. [Google Scholar]
- Ruiu, L.; Mannu, R.; Falchi, G.; Braggio, A.; Luciano, P. Evaluation of different Bacillus thuringiensis sv kurstaki formulations against Lymantria dispar and Malacosoma neustria larvae infesting Quercus suber trees. Redia 2013, 96, 27–31. [Google Scholar]
- Ruiu, L.; Mannu, R.; Olivieri, M.; Lentini, A. Gypsy Moth Management with LdMNPV Baculovirus in Cork Oak Forest. Forests 2021, 12, 495. [Google Scholar] [CrossRef]
- Lentini, A.; Mannu, R.; Cocco, A.; Ruiu, P.A.; Cerboneschi, A.; Luciano, P. Long-term monitoring and microbiological control programs against lepidopteran defoliators in Sardinian cork oak forests (Italy). Ann. Silvic. Res. 2020, 45, 21–30. [Google Scholar] [CrossRef]
- Mannu, R.; Cocco, A.; Luciano, P.; Lentini, A. Influence of Bacillus thuringiensis application timing on population dynamics of gypsy moth in Mediterranean cork oak forests. Pest. Manag. Sci. 2020, 76, 1103–1111. [Google Scholar] [CrossRef]
- Reardon, R.C.; Podgwaite, J.D. Summary of efficacy evaluations using aerially applied Gypchek® against gypsy moth in the USA. J. Environ. Sci. Health 1994, 29, 739–756. [Google Scholar] [CrossRef]
- Glare, T.R.; O’Callaghan, M. Bacillus thuringiensis. In Biology, Ecology and Safety; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Maczuga, S.A.; Mierzejewski, K.J. Droplet size and density effects of Bacillus thuringiensis kurstaki on gypsy moth (Lepidoptera: Lymantriidae) larvae. J. Econ. Entomol. 1995, 88, 1376–1379. [Google Scholar] [CrossRef]
- Lentini, A.; Luciano, P. Bacillus thuringiensis in the management of gypsy moth (Lymantria dispar L.) in Sardinian cork-oak forests. IOBC/WPRS Bull 1995, 18, 104–109. [Google Scholar]
- Lentini, A.; Cocco, A.; Luciano, P. Effects of Bacillus thuringiensis kurstaki treatments on gypsy moth population dynamics. IOBC/WPRS Bull 2012, 76, 171–174. [Google Scholar]
- Ruiu, L.; Lentini, A.; Coinu, M.; Loi, A.; Serra, G.; Luciano, P. Comparative applications of Bacillus thuringiensis formulations against Lymantria dispar in Sardinian forests. IOBC/WPRS Bull 2012, 76, 185–190. [Google Scholar]
- Boucias, D.G.; Pendland, J.C. Principles of Insect Pathology; Kluwer Academic Publisher: Boston, MA, USA, 1998. [Google Scholar]
- Ellar, D.J.; Knowles, B.H.; Carroll, J.; Horsnell, J.; Haider, M.Z.; Ahmad, W.; Nicholls, C.N.; Armstrong, G.; Hodgman, T.C. Genetic and biochemical studies of the mechanism of action of Bacillus thuringiensis entomocidal-endotoxins. In Bacterial Toxins: Zentralblatt für Bakteriologie; International Medical Microbiology, Supplement 19; Rappuoli, R., Alouf, J., Freer, J., Fehrenbach, F., Wadstrom, T., Witholt, B., Eds.; Gustav Fischer: Stuttgart, Germany, 1990; pp. 409–506. [Google Scholar]
- Crickmore, N. Beyond the spore–past and future developments of Bacillus thuringiensis as a biopesticide. J. Appl. Microbiol. 2006, 101, 616–619. [Google Scholar] [CrossRef]
- Van Frankenhuyzen, K.; Fast, P.G. Susceptibility of Three Coniferophagous Choristoneura Species (Lepidoptera: Tortricidae) to Bacillus thuringiensis var. kurstaki. J. Econ. Entomol. 1989, 82, 193–196. [Google Scholar] [CrossRef]
- Boulton, T.J.; Otvos, I.S.; Ring, R.A. Monitoring Nontarget Lepidoptera on Ribes cereum to Investigate Side Effects of an Operational Application of Bacillus thuringiensis subsp. kurstaki. Environ. Entomol. 2002, 31, 903–913. [Google Scholar] [CrossRef] [Green Version]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Satinder, K.B.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Recent advances in downstream processing and formulations of Bacillus thuringiensis based biopesticides. Process Biochem. 2006, 41, 323–342. [Google Scholar] [CrossRef]
- Dubois, N.R. New laboratory and field developments in Bacillus thuringiensis and host susceptibility. In Proceedings of the U.S. Department of Agriculture Interagency Gypsy Moth Research Forum, Annapolis, MD, USA, 19–22 January 2003. [Google Scholar]
- Cocco, A.; Cossu, A.Q.; Erre, P.; Nieddu, G.; Luciano, P. Spatial analysis of gypsy moth populations in Sardinia using geostatistical and climate models. Agr. For. Entomol. 2010, 12, 417–426. [Google Scholar] [CrossRef]
- Mannu, R.; Gilioli, G.; Luciano, P. Occupancy of the territory by Lymantria dispar (L.) (Lepidoptera Erebidae) egg masses as a predictive index of damage. Redia 2017, 100, 157–165. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Pünterner, W. Manual for field trials in plant protection. In Agricultural Division; Ciba-Geigy, Ltd.: Basle, Switzerland, 1981. [Google Scholar]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Therneau, T.M. Coxme: Mixed Effects Cox Models. R Package Version 2.2-5. Available online: https://CRAN.R-project.org/package=coxme (accessed on 24 April 2021).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Alalouni, U.; Schädler, M.; Brandl, R. Natural enemies and environmental factors affecting the population dynamics of the gypsy moth. J. Appl. Entomol. 2013, 137, 721–738. [Google Scholar] [CrossRef]
- Eichhorn, O. Experimental studies upon the parasitoid complex of the gypsy moth (Lymantria dispar L.) (Lep. Lymantriidae) in lower host populations in eastern Austria. J. Appl. Entomol. 1996, 120, 205–212. [Google Scholar] [CrossRef]
- Hoch, G.; Zubrik, M.; Novotny, J.; Schopf, A. The natural enemy complex of the gypsy moth, Lymantria dispar (Lep. Lymantriidae) in different phases of its population dynamics in eastern Austria and Slovakia–A comparative study. J. Appl Entomol 2001, 125, 217–227. [Google Scholar] [CrossRef]
- Contarini, M.; Luciano, P.; Pilarska, D.; Pilarski, P.; Solter, L.; Huang, W.F.; Georgiev, G. Survey of pathogens and parasitoids in late instar Lymantria dispar larval populations in Sardinia, Italy. Bull. Insectol. 2013, 66, 51–58. [Google Scholar]
- Ladurner, E.; Benuzzi, M.; Franceschini, S. Bacillus thuringiensis sv kurstaki strain EG 2348: Effect of formulation on efficacy against tomato leaf miner (Tuta absoluta). IOBC/WPRS Bull 2011, 66, 39–42. [Google Scholar]
- McGaughey, W.H.; Beeman, R.W. Resistance to Bacillus thuringiensis in Colonies of Indianmeal Moth and Almond Moth (Lepidoptera: Pyralidae). J. Econ. Entomol. 1988, 81, 28–33. [Google Scholar] [CrossRef]
- Ferré, J.; Van Rie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef]
- Janmaat, A.F.; Myers, J. Rapid evolution and the cost of resistance to Bacillus thuringiensis in greenhouse populations of cabbage loopers Trichoplusia ni. Proc. Biol. Sci. 2003, 270, 2263–2270. [Google Scholar] [CrossRef] [Green Version]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Ann. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | Treatments | First Instar | Second Instar | Third Instar | Fourth Instar |
|---|---|---|---|---|---|
| 2018 | Foray 2.0 L/ha | 242 | 521 | 121 | - |
| Foray 2.5 L/ha | 278 | 527 (−) | 218 (+) | - | |
| Rapax | 106 (+) | 120 | 8 (−) | - | |
| Control | 192 | 467 (+) | 65 (−) | - | |
| 2019 | Foray 2.0 L/ha | 11 (−) | 190 (−) | 163 (+) | 24 (+) |
| Foray 2.5 L/ha | 15 | 118 | 61 (+) | 2 | |
| Rapax | 42 | 308 | 46 (−) | 0 | |
| Control | 48 | 394 (+) | 26 (−) | 1 (−) |
| Year | Foray 2.0 L/ha | Foray 2.5 L/ha | Rapax | Control | F | p |
|---|---|---|---|---|---|---|
| 2018 | 294.7 ± 101.3 | 247.0 ± 19.2 | 78.0 ± 35.7 | 241.3 ± 65.9 | 2.20 | 0.17 |
| 2019 | 129.7 ± 61.6 | 65.3 ± 13.7 | 132 ± 12.2 | 156.3 ± 51.9 | 0.89 | 0.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivieri, M.; Mannu, R.; Ruiu, L.; Ruiu, P.A.; Lentini, A. Comparative Efficacy Trials with Two Different Bacillus thuringiensis Serovar kurstaki Strains against Gypsy Moth in Mediterranean Cork Oak Forests. Forests 2021, 12, 602. https://doi.org/10.3390/f12050602
Olivieri M, Mannu R, Ruiu L, Ruiu PA, Lentini A. Comparative Efficacy Trials with Two Different Bacillus thuringiensis Serovar kurstaki Strains against Gypsy Moth in Mediterranean Cork Oak Forests. Forests. 2021; 12(5):602. https://doi.org/10.3390/f12050602
Chicago/Turabian StyleOlivieri, Maurizio, Roberto Mannu, Luca Ruiu, Pino A. Ruiu, and Andrea Lentini. 2021. "Comparative Efficacy Trials with Two Different Bacillus thuringiensis Serovar kurstaki Strains against Gypsy Moth in Mediterranean Cork Oak Forests" Forests 12, no. 5: 602. https://doi.org/10.3390/f12050602
APA StyleOlivieri, M., Mannu, R., Ruiu, L., Ruiu, P. A., & Lentini, A. (2021). Comparative Efficacy Trials with Two Different Bacillus thuringiensis Serovar kurstaki Strains against Gypsy Moth in Mediterranean Cork Oak Forests. Forests, 12(5), 602. https://doi.org/10.3390/f12050602