
No Long-Term Decrease in Caterpillar Availability for Invertivorous Birds in Deciduous Forests in Hungary

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Estimating the Relative Abundances of the Selected Species
- The trap was set in or near a broadleaved (mainly oak) forest stand.
- Has been working for at least 20 years.
- Has been operated continuously, with a max. one year “time gap”.
- The last year in the time series had to be 2019.
- The species was univoltine.
- Its caterpillars fed on the foliage of woody plants (trees and shrubs).
- Its caterpillars developed in spring or early summer (from April to June) overlapping with the breeding season of invertivorous birds.
- The caterpillars were neither densely hairy, nor contained poisonous chemicals, likely to be regularly consumed by invertivorous birds.
- The species did not overwinter as adult.
- The species was regularly caught in most years and places.
2.2. Estimating the Caterpillar Body Mass
2.3. Calculation of the Estimated Total Caterpillar Biomass Index
2.4. Statistical Analysis
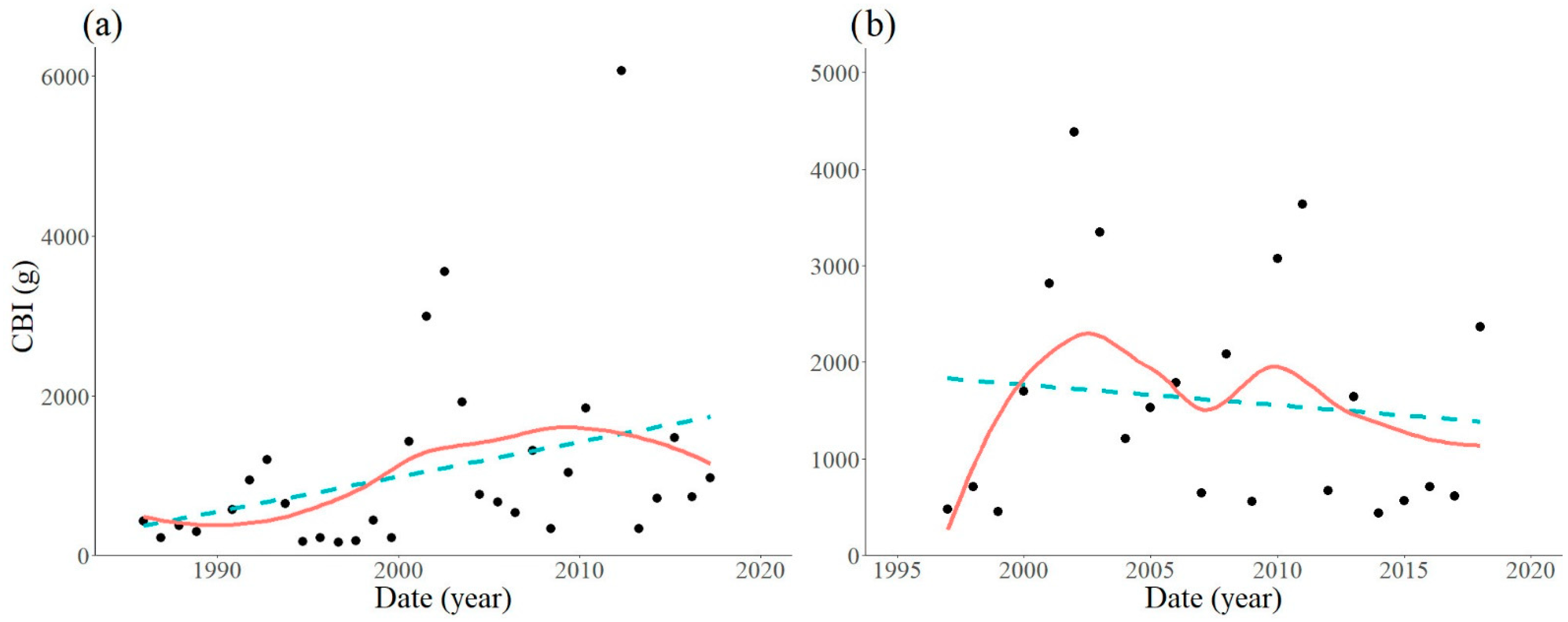
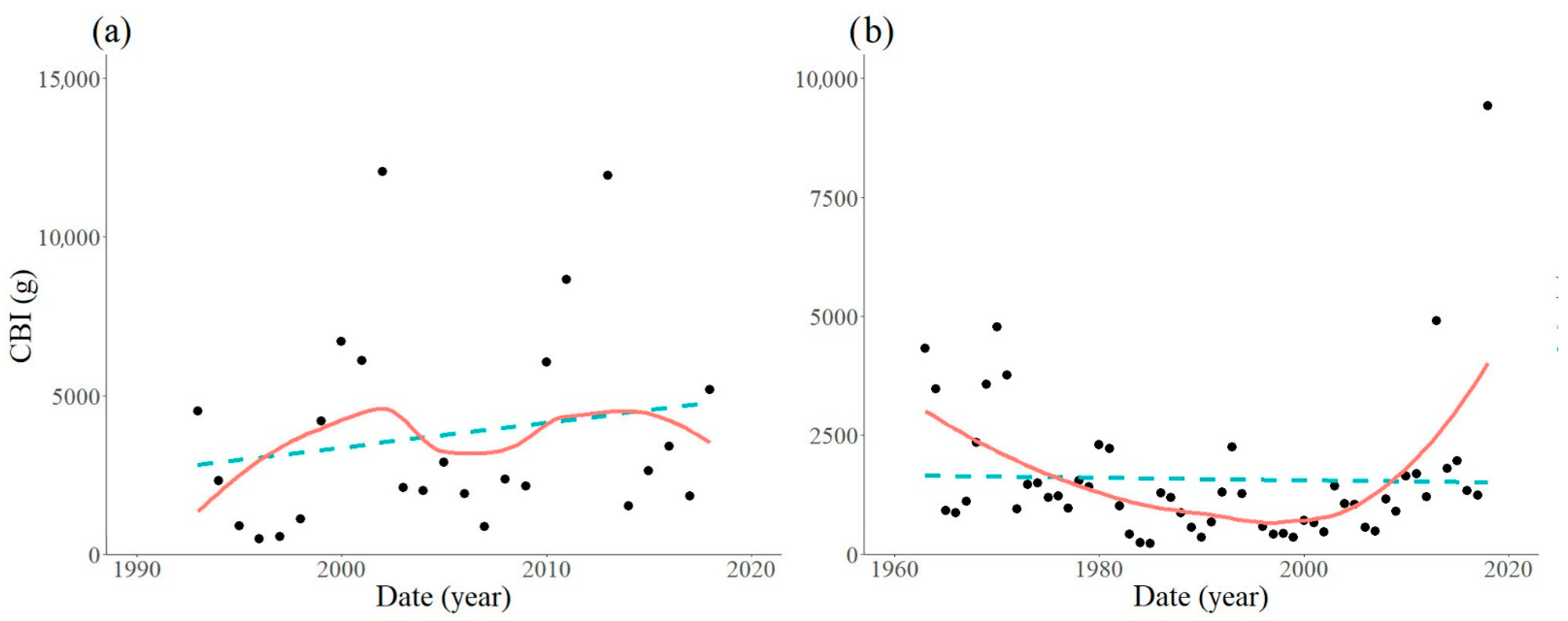
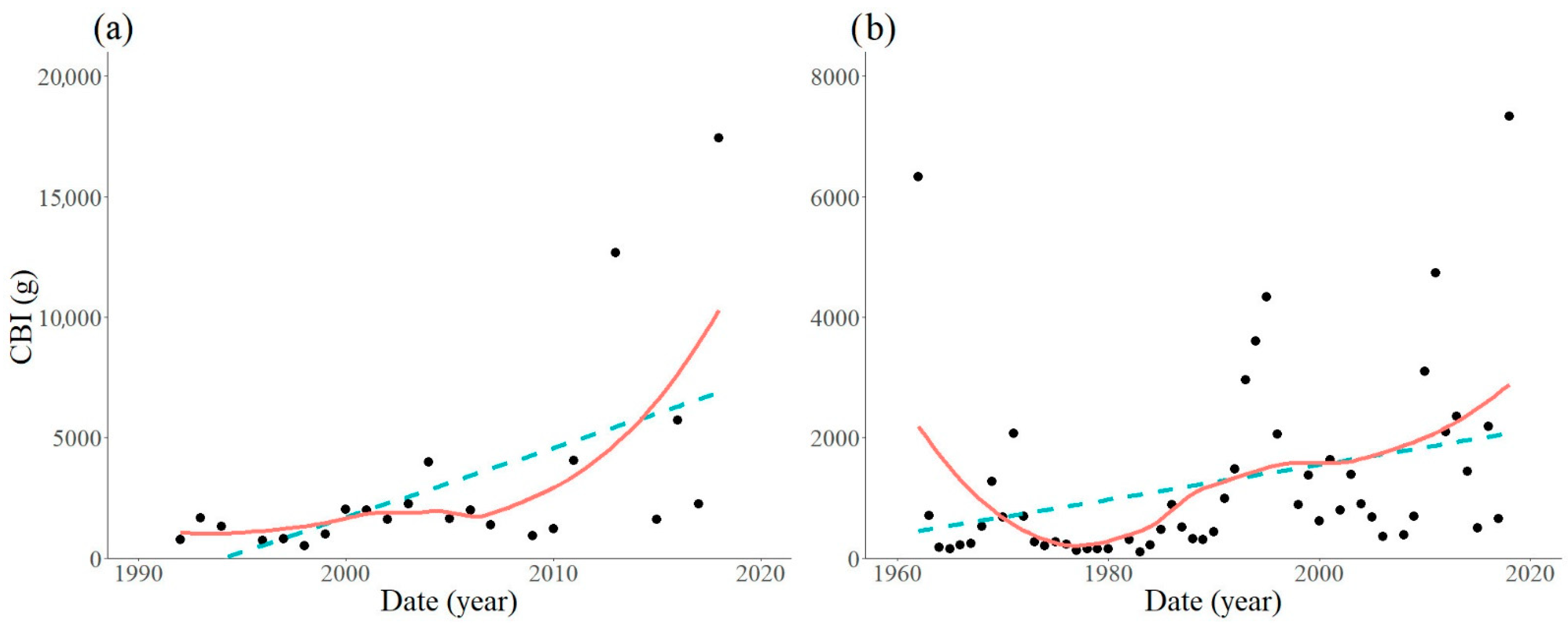
3. Results
3.1. Locations with Non-Significant Trends
3.2. Locations with Significant Trends
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, C.A.; Zeegers, T.; Klink, R.; Vermeulen, R.; Wielink, P.; Spijkers, H.; Deijk, J.; Steenis, W.; Jongejans, E. Declining abundance of beetles, moths and caddisflies in the Netherlands. Insect Conserv. Divers. 2020, 13, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Valtonen, A.; Hirka, A.; Szőcs, L.; Ayres, M.P.; Roininen, H.; Csóka, G. Long-term species loss and homogenization of moth communities in Central Europe. J. Anim. Ecol. 2017, 86, 730–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Leather, S.R. “Ecological Armageddon”—More evidence for the drastic decline in insect numbers. Ann. Appl. Biol. 2018, 172, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Szontagh, P. Outbreak and damage of the lackey moth (Malacosoma neustria L.) on or oak forests. Erdészeti Kut. 1962, 58, 125–142. (In Hungarian) [Google Scholar]
- Szontagh, P. The role of light traps in the prognosis of forest pests. Növényvédelem 1975, 11, 54–57. (In Hungarian) [Google Scholar]
- Tallós, P. The role of light traps in forest protection. Az Erdő 1966, 15, 134–136. (In Hungarian) [Google Scholar]
- Hirka, A.; Szabóky, C.; Szőcs, L.; Csóka, G. 50 years of the Forestry Light Trap Network. Növényvédelem 2011, 47, 474–479. (In Hungarian) [Google Scholar]
- Csóka, G.; Hirka, A.; Szőcs, L.; Móricz, N.; Rasztovits, E.; Podor, Z. Weather-dependent fluctuations in the abundance of the oak processionary moth, Thaumetopoea processionea (Lepidoptera: Notodontidae). Eur. J. Entomol. 2018, 115, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Schowalter, T.D. Insects as Regulators of Ecosystem Processes. In Insect Ecology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 511–537. [Google Scholar]
- Schowalter, T.D.; Noriega, J.A.; Tscharntke, T. Insect effects on ecosystem services—Introduction. Basic Appl. Ecol. 2018, 26, 1–7. [Google Scholar] [CrossRef]
- Dangles, O.; Casas, J. Ecosystem services provided by insects for achieving sustainable development goals. Ecosyst. Serv. 2019, 35, 109–115. [Google Scholar] [CrossRef]
- Losey, J.E.; Vaughan, M. The Economic Value of Ecological Services Provided by Insects. Bioscience 2006, 56, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Şekercioğlu, Ç.H. Ecological significance of bird populations. In Handbook of the Birds of the World—Volume 11; del Hoyo, J., Elliott, A., Christie, D.A., BirdLife International, Eds.; Lynx Edicions: Cambridge, UK; Barcelona, Spain, 2006; pp. 15–51. [Google Scholar]
- Morse, D.H. The Insectivorous Bird as an Adaptive Strategy. Annu. Rev. Ecol. Syst. 2017, 2, 177–200. [Google Scholar] [CrossRef]
- Nyffeler, M.; Şekercioğlu, Ç.H.; Whelan, C.J. Insectivorous birds consume an estimated 400–500 million tons of prey annually. Sci. Nat. 2018, 105, 47. [Google Scholar] [CrossRef] [Green Version]
- Holmes, R.T.; Schultz, J.C.; Nothnagle, P. Bird Predation on Forest Insects: An Exclosure Experiment. Science 1979, 206, 462–463. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, J.J.; Anderson, G.Q.A.; Grice, P.V.; Vickery, J.A.; Watts, P.N.; Sutherland, W.J. Foraging habitat selection, diet and nestling condition in Yellow Wagtails Motacilla flava breeding on arable farmland. Bird Study 2009, 56, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Pagani-Núñez, E.; Renom, M.; Mateos-Gonzalez, F.; Cotín, J.; Senar, J.C. The diet of great tit nestlings: Comparing observation records and stable isotope analyses. Basic Appl. Ecol. 2017, 18, 57–66. [Google Scholar] [CrossRef]
- Tremblay, I.; Thomas, D.; Blondel, J.; Perret, P.; Lambrechts, M.M. The effect of habitat quality on foraging patterns, provisioning rate and nestling growth in Corsican Blue Tits Parus caeruleus. Ibis 2005, 147, 17–24. [Google Scholar] [CrossRef]
- Gibb, J.A.; Betts, M.M. Food and Food Supply of Nestling Tits (Paridae) in Breckland Pine. J. Anim. Ecol. 1963, 32, 489. [Google Scholar] [CrossRef]
- Török, J.; Tóth, L. Asymmetric competition between two tit species: A reciprocal removal experiment. J. Anim. Ecol. 1999, 68, 338–345. [Google Scholar] [CrossRef]
- Seress, G.; Sándor, K.; Evans, K.L.; Liker, A. Food availability limits avian reproduction in the city: An experimental study on great tits Parus major. J. Anim. Ecol. 2020, 89, 1570–1580. [Google Scholar] [CrossRef]
- Perrins, C.M. Tits and their caterpillar food supply. Ibis 2008, 133, 49–54. [Google Scholar] [CrossRef]
- Seress, G.; Hammer, T.; Bókony, V.; Vincze, E.; Preiszner, B.; Pipoly, I.; Sinkovics, C.; Evans, K.L.; Liker, A. Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird. Ecol. Appl. 2018, 28, 1143–1156. [Google Scholar] [CrossRef] [Green Version]
- Csóka, G. Lepkehernyók (Caterpillars); Agroinform: Budapest, Hungary, 1996; ISBN 2050000014741. [Google Scholar]
- Porter, J. Colour Identification Guide to Caterpillars of the British Isles. Macrolepidoptera; Viking: London, UK, 1997; ISBN 8788757951. [Google Scholar]
- Leskó, K.; Szentkirályi, F.; Kádár, F. Fluctuation patterns of the Gipsy moth (Lymantria dispar L.) population between 1963–1990 in Hungary. Erdészeti Kut. 1994, 84, 163–176. (In Hungarian) [Google Scholar]
- Leskó, K.; Szentkirályi, F.; Kádár, F. Long term fluctuation pattern of the brown -tale moth (Euproctis chrysorrhoea L.) Hungarian population. Erdészeti Kut. 1995, 85, 169–185. (In Hungarian) [Google Scholar]
- Leskó, K.; Szentkirályi, F.; Kádár, F. Long term (1962-1996) fluctuation of the lackey moth population in Hungary. Erdészeti Kut. 1997, 86–87, 171–200. (In Hungarian) [Google Scholar]
- Leskó, K.; Szentkirályi, F.; Kádár, F. An analysis of fluctuation pattern of geometrid moths based on long term (1961–1997) light trap and damage data time series in Hungary. Erdészeti Kut. 1998, 88, 319–333. (In Hungarian) [Google Scholar]
- Leskó, K.; Szentkirályi, F.; Kádár, F. Characterisation of the long term (1962–1997) fluctuation of winter moth (Operophtera brumata L.) population based on Forestry Light Trap data. Erdészeti Kut. 1999, 89, 169–182. (In Hungarian) [Google Scholar]
- R Foundation for Statistical Computing. R Development Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Cleveland, W.S. Robust Locally Weighted Regression and Smoothing Scatterplots. J. Am. Stat. Assoc. 1979, 74, 829. [Google Scholar] [CrossRef]
- Macgregor, C.J.; Williams, J.H.; Bell, J.R.; Thomas, C.D. Moth biomass has fluctuated over 50 years in Britain but lacks a clear trend. Nat. Ecol. Evol. 2019, 3, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Abery, J.C.G. Estimating rates of butterfly decline from distribution maps: The effect of scale. Biol. Conserv. 1995, 73, 59–65. [Google Scholar] [CrossRef]
- Thomas, J.A. Comparative Losses of British Butterflies, Birds, and Plants and the Global Extinction Crisis. Science 2004, 303, 1879–1881. [Google Scholar] [CrossRef] [Green Version]
- Conrad, K.F.; Woiwod, I.P.; Parsons, M.; Fox, R.; Warren, M.S. Long-term population trends in widespread British moths. J. Insect Conserv. 2004, 8, 119–136. [Google Scholar] [CrossRef]
- Conrad, K.F.; Warren, M.S.; Fox, R.; Parsons, M.S.; Woiwod, I.P. Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biol. Conserv. 2006, 132, 279–291. [Google Scholar] [CrossRef]
- Jactel, H.; Petit, J.; Desprez-Loustau, M.-L.; Delzon, S.; Piou, D.; Battisti, A.; Koricheva, J. Drought effects on damage by forest insects and pathogens: A meta-analysis. Glob. Chang. Biol. 2012, 18, 267–276. [Google Scholar] [CrossRef]
- Klapwijk, M.J.; Walter, J.A.; Hirka, A.; Csóka, G.; Björkman, C.; Liebhold, A.M. Transient synchrony among populations of five foliage-feeding Lepidoptera. J. Anim. Ecol. 2018, 87, 1058–1068. [Google Scholar] [CrossRef] [Green Version]
- Wainhouse, D.; Inward, D.J.G. The influence of climate change on forest insect pests in Britain. FCRN 2016, 021, 1–10. [Google Scholar]
- Csóka, G. Increased insect damage in Hungarian forests under drought impact. Biologia 1997, 52, 159–162. [Google Scholar]
- Zúbrik, M.; Hajek, A.; Pilarska, D.; Špilda, I.; Georgiev, G.; Hrašovec, B.; Hirka, A.; Goertz, D.; Hoch, G.; Barta, M.; et al. The potential for Entomophaga maimaiga to regulate gypsy moth Lymantria dispar (L.) (Lepidoptera: Erebidae) in Europe. J. Appl. Entomol. 2016, 140, 565–579. [Google Scholar] [CrossRef]
- McManus, M.; Csóka, G. History and Impact of Gypsy Moth in North America and Comparison to the Recent Outbreaks in Europe. Acta Silv. Lignaria Hung. 2007, 3, 47–64. [Google Scholar]
- Hlásny, T.; Trombik, J.; Holuša, J.; Lukášová, K.; Grendár, M.; Turčáni, M.; Zúbrik, M.; Tabaković-Tošić, M.; Hirka, A.; Buksha, I.; et al. Multi-decade patterns of gypsy moth fluctuations in the Carpathian Mountains and options for outbreak forecasting. J. Pest Sci. 2016, 89, 413–425. [Google Scholar] [CrossRef]
- Hajek, A.E.; Butler, L.; Wheeler, M.M. Laboratory Bioassays Testing the Host Range of the Gypsy Moth Fungal Pathogen Entomophaga maimaiga. Biol. Control 1995, 5, 530–544. [Google Scholar] [CrossRef]
- Hajek, A.E.; Butler, L.; Walsh, S.R.A.; Silver, J.C.; Hain, F.P.; Hastings, F.L.; Odell, T.M.; Smitley, D.R. Host Range of the Gypsy Moth (Lepidoptera: Lymantriidae) Pathogen Entomophaga maimaiga (Zygomycetes: Entomophthorales) in the Field versus Laboratory. Environ. Entomol. 1996, 25, 709–721. [Google Scholar] [CrossRef]
- Zúbrik, M.; Špilda, I.; Pilarska, D.; Hajek, A.E.; Takov, D.; Nikolov, C.; Kunca, A.; Pajtík, J.; Lukášová, K.; Holusa, J. Distribution of the entomopathogenic fungus Entomophaga maimaiga (Entomophthorales: Entomophthoraceae) at the northern edge of its range in Europe. Ann. Appl. Biol. 2018, 173, 35–41. [Google Scholar] [CrossRef]
- Pilarska, D.; McManus, M.; Hajek, A.E.; Herard, F.; Vega, F.E.; Pilarski, P.; Markova, G. Introduction of the entomopathogenic fungus Entomophaga maimaiga Hum., Shim. and Sop. (Zygomycetes: Entomophthorales) to a Lymantria dispar (L.) (Lepidoptera: Lymantriidae) population in Bulgaria. Anzeiger Schadlingskd. 2000, 73, 125–126. [Google Scholar] [CrossRef]
- Csóka, G.; Hirka, A.; Szőcs, L.; Hajek, A.E. First record of the pathogenic fungus Entomophaga maimaiga Humber, Shimazu and Soper, 1988 (Entomophtorales: Entomphtoraceae) in hungarian Gipsy moth (Lymantria dispar) populations. Növényvédelem 2014, 50, 257–262. (In Hungarian) [Google Scholar]
- Hrašovec, B.; Pernek, M.; Lukić, I.; Milotić, M.; Diminić, D.; Franjević, M.; A., H.; Linde, A.; Pilarska, D. First record of the pathogenic fungus Entomophaga maimaiga Humber, Shimazu, and Soper (Entomophthorales: Entomophthoraceae) within an outbreak populations of Lymantria dispar (Lepidoptera: Erebidae) in Croatia. Period. Biol. 2013, 115, 379–383. [Google Scholar]
- Zúbrik, M.; Barta, M.; Pilarska, D.; Goertz, D.; Úradník, M.; Galko, J.; Vakula, J.; Gubka, A.; Rell, S.; Kunca, A. First record of Entomophaga maimaiga (Entomophthorales: Entomophthoraceae) in Slovakia. Biocontrol Sci. Technol. 2014, 24, 710–714. [Google Scholar] [CrossRef]
- Georgiev, G.; Mirchev, P.; Rossnev, B.; Petkov, P.; Georgieva, M.; Pilarska, D.; Golemansky, V.; Pilarski, P.; Hubenov, Z. Potential of Entomophaga maimaiga Humber, Shimazu and Soper (Entomophthorales) for suppressing Lymantria dispar (Linnaeus) outbreaks in Bulgaria. Comptes Rendus L’Academie Bulg. Sci. 2013, 66, 1025–1032. [Google Scholar] [CrossRef]
- Manderino, R.; Crist, T.O.; Haynes, K.J. Lepidoptera-specific insecticide used to suppress gypsy moth outbreaks may benefit non-target forest Lepidoptera. Agric. For. Entomol. 2014, 16, 359–368. [Google Scholar] [CrossRef]
- Welti, E.A.R.; Joern, A.; Ellison, A.M.; Lightfoot, D.C.; Record, S.; Rodenhouse, N.; Stanley, E.H.; Kaspari, M. Studies of insect temporal trends must account for the complex sampling histories inherent to many long-term monitoring efforts. Nat. Ecol. Evol. 2021, 5, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Török, J. Food segregation in three hole-nesting bird species during the breeding season. Ardea 1986, 74, 129–136. [Google Scholar]
- Bereczki, K.; Ódor, P.; Csóka, G.; Mag, Z.; Báldi, A. Effects of forest heterogeneity on the efficiency of caterpillar control service provided by birds in temperate oak forests. For. Ecol. Manag. 2014, 327, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Kirstin, A.; Patocka, J. Birds as predators of Lepidoptera: Selected examples. Biologia 1997, 52, 319–326. [Google Scholar]
- Török, J. Resource partitioning among three woodpecker species Dendrocopos spp. during the breeding season. Ecography 1990, 13, 257–264. [Google Scholar] [CrossRef]
- Duna-Ipoly Nemzeti Park Igazgatóság. Forest State Evaluation in Upland Forests of Hungary—Rosalia a Duna-Ipoly Nemzeti Park Igazgatóság tanulmánykötetei 9; Standovár, T., Bán, M., Kézdi, P., Szerk, Eds.; Duna-Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2017. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
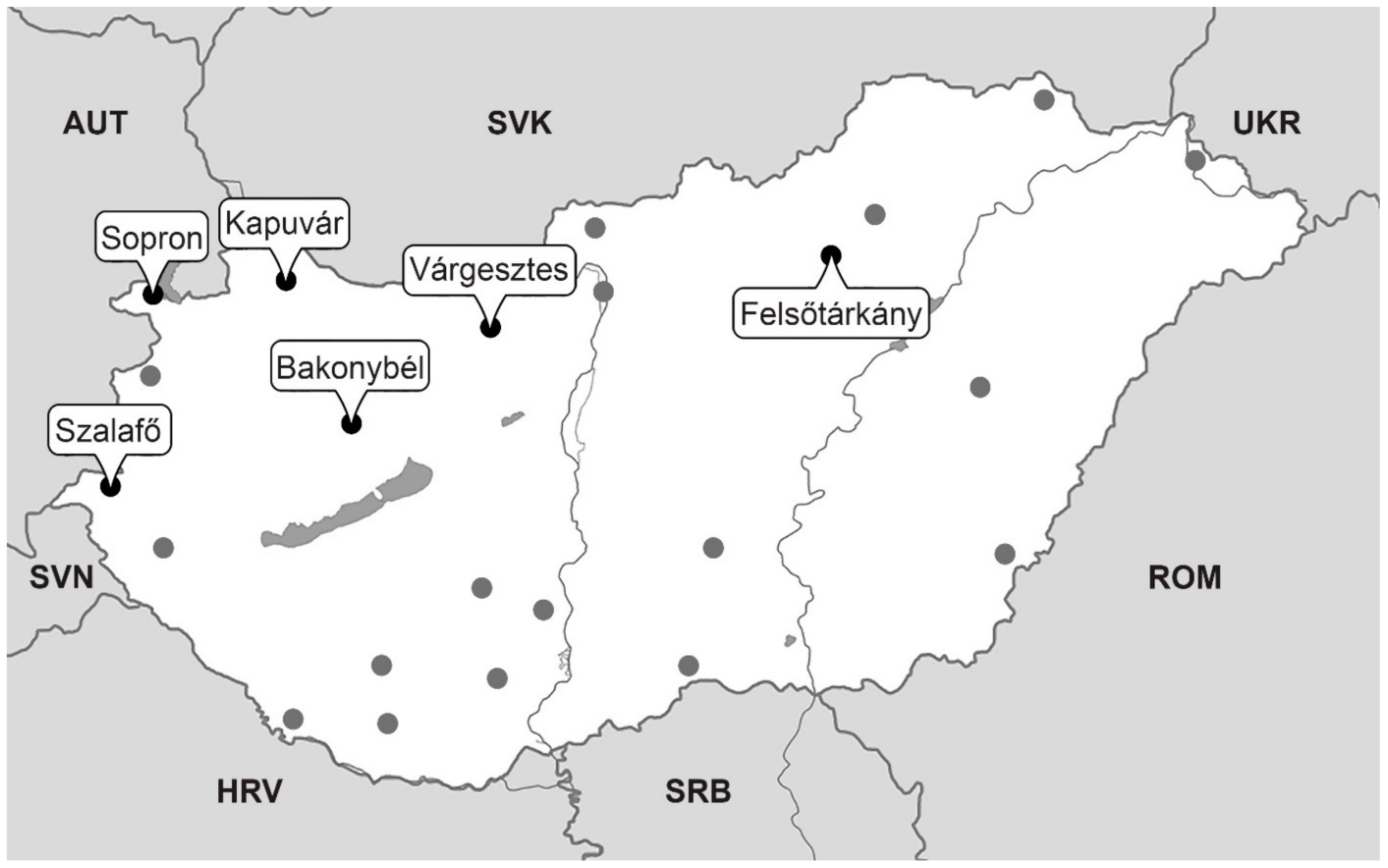
| Trap Location | Trap Coordinates | Altitude (m) | Forest Stands Around the Trap | Period | Length (Years) | Number of Individuals in Calculation |
|---|---|---|---|---|---|---|
| Felsőtárkány | 47°58′51″ N 20°26′03″ E | 238 | Oak (Quercus petraea (Matt.) Liebl.) and hornbeam (Carpinus betulus L.) dominated mixed forests. | 1962–2019 | 58 | 108,224 |
| Várgesztes | 47°28′18″ N 18°23′54″ E | 286 | Turkey oak (Quercus cerris DC.) and hornbeam (C. betulus) dominated mixed forests. | 1963–2019 | 57 | 138,589 |
| Bakonybél | 47°15′04″ N 17°45′41″ E | 390 | Oak (Quercus robur L.) and hornbeam (C. betulus) dominated mixed forests. | 1992–2019 | 28 | 59,051 |
| Kapuvár | 47°41′16″ N 17°00′30″ E | 120 | Oak (Q. robur and Q. cerris) dominated mixed broadleaved forests. | 1993–2019 | 27 | 83,179 |
| Sopron | 47°39′51″ N 16°33′14″.E | 375 | Oak (Q. petraea) and beech (Fagus sylvatica L.) dominated mixed broadleaved forests. | 1997–2019 | 23 | 41,036 |
| Szalafő | 46°51′19″ N 16°22′33″ E | 264 | Oak (Q. robur) and hornbeam (C. betulus) dominated mixed broadleaved forests. | 1986–2019 | 34 | 40,403 |
| Trap Location | Period Analysed | Equation | R2 | F-Statistic | DF | p-Value |
|---|---|---|---|---|---|---|
| Szalafő | 1986–2018 | Y = 42.6X − 84280.4 | 0.1102 | 3.591 | 29 | 0.0681 |
| Sopron | 1997–2018 | Y = −21.1X + 44024.5 | 0.0131 | 0.266 | 20 | 0.6117 |
| Kapuvár | 1993–2018 | Y = 78.1X − 152889.2 | 0.0342 | 0.814 | 23 | 0.3764 |
| Várgesztes | 1963–2018 | Y = −2.6X + 2565.4 | 0.0001 | 0.042 | 53 | 0.8398 |
| Várgesztes | 1997–2018 | Y = 175.4X − 350478.0 | 0.3257 | 9.659 | 20 | 0.0056 |
| Bakonybél | 1992–2018 | Y = 288.0X − 574272.9 | 0.3199 | 9.877 | 21 | 0.0049 |
| Felsőtárkány | 1962–2018 | Y = 29.0X − 56333.8 | 0.0994 | 5.738 | 52 | 0.0202 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eötvös, C.B.; Hirka, A.; Gimesi, L.; Lövei, G.L.; Gáspár, C.; Csóka, G. No Long-Term Decrease in Caterpillar Availability for Invertivorous Birds in Deciduous Forests in Hungary. Forests 2021, 12, 1070. https://doi.org/10.3390/f12081070
Eötvös CB, Hirka A, Gimesi L, Lövei GL, Gáspár C, Csóka G. No Long-Term Decrease in Caterpillar Availability for Invertivorous Birds in Deciduous Forests in Hungary. Forests. 2021; 12(8):1070. https://doi.org/10.3390/f12081070
Chicago/Turabian StyleEötvös, Csaba Béla, Anikó Hirka, László Gimesi, Gábor L. Lövei, Csaba Gáspár, and György Csóka. 2021. "No Long-Term Decrease in Caterpillar Availability for Invertivorous Birds in Deciduous Forests in Hungary" Forests 12, no. 8: 1070. https://doi.org/10.3390/f12081070
APA StyleEötvös, C. B., Hirka, A., Gimesi, L., Lövei, G. L., Gáspár, C., & Csóka, G. (2021). No Long-Term Decrease in Caterpillar Availability for Invertivorous Birds in Deciduous Forests in Hungary. Forests, 12(8), 1070. https://doi.org/10.3390/f12081070