
Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Plots
2.2. Field Methods
2.3. Laboratory Methods
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garbelotto, M.; Gonthier, P. Biology, epidemiology, and control of Heterobasidion species worldwide. Annu. Rev. Phytopathol. 2013, 51, 39–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaitnieks, T.; Arhipova, N.; Donis, J.; Stendlid, J.; Vasaitis, R. Butt rot incidence and related losses in Latvian Picea abies (L.) Kasrt. stands. In Proceedings of the 12th International Conference on Root and Butt Rots, 12–19 August 2007; The University of California: Berkley, CA, USA, 2008; pp. 177–179. [Google Scholar]
- Korhonen, K.; Stenlid, J. Biology of Heterobasidion annosum. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 43–70. [Google Scholar]
- Stenlid, J. Regional differentiation in Heterobasidion annosum. In Proceedings of the Root and Butt Rots: Proceedings of the International Conference on Root and Butt Rots (IUFRO) Working Party S2.06.01, Wik, Sweden and Haikko, Finland, 9–16 August 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1994; pp. 243–248. [Google Scholar]
- Gaitnieks, T.; Bruna, L.; Zaluma, A.; Burnevica, N.; Klavina, D.; Legzdina, L.; Jansons, J.; Piri, T. Development of Heterobasidion spp. fruit bodies on decayed Piecea abies. For. Ecol. Manag. 2021, 482, 118835. [Google Scholar] [CrossRef]
- Schütt, P.; Schuck, H.J.; Lautenschlager, K.; Prestle, W.; Stimm, B. Fomes annosus sporocarps—Their abundance on decayed logs left in the forest. Eur. J. For. Pathol. 1979, 9, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.M.; Heinonen, J.; Korhonen, K. Occurrence of Heterobasidion basidiocarps on cull pieces of Norway spruce left on cutting areas and in mature spruce stands. For. Pathol. 2007, 37, 374–386. [Google Scholar] [CrossRef]
- Möykkynen, T.; Von Weissenberg, K.; Pappinen, A. Estimation of dispersal gradients of S- and P-type basidiospores of Heterobasidion annosum. Eur. J. For. Pathol. 1997, 27, 291–300. [Google Scholar] [CrossRef]
- Gonthier, P.; Garbelotto, M.M.; Nicolotti, G. Seasonal patterns of spore deposition of Heterobasidion species in four forests of the western Alps. Phytopathology 2005, 95, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Penttilä, R.; Siitonen, J.; Kuusinen, M. Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol. Conserv. 2004, 117, 271–283. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Hanski, I. Fungivory: Fungi, Insects and Ecology. In Insect Fungus Interactions, Proceedings of the 14th Symposium of the Royal Entomological Society of London in Collabaration with the British Mycological Society, London, UK, 16–17 September 1989; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989; pp. 25–61. [Google Scholar]
- Hammond, P.; Lawrence, J.F. Appendix. Mycophagy in insects: A summary. In Insect Fungus Interactions, Proceedings of the 14th Symposium of the Royal Entomological Society of London in Collabaration with the British Mycological Society, London, UK, 16–17 September 1989; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989; pp. 275–324. [Google Scholar]
- Hågvar, S.; Steen, R. Succession of beetles (genus Cis) and oribatid mites (genus Carabodes) in dead sporocarps of the red-banded polypore fungus Fomitopsis pinicola. Scand. J. For. Res. 2013, 28, 436–444. [Google Scholar] [CrossRef]
- Jonsell, M.; Nordlander, G. Insects in polypore fungi as indicator species: A comparison between forest sites differing in amounts and continuity of dead wood. For. Ecol. Manag. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Komonen, A.; Jonsell, M.; Okland, B.; Sverdrup-Thygeson, A.; Thunes, K. Insect assemblage associated with the polypore Fomitopsis pinicola: A comparison across Fennoscandia. Entomol. Fenn. 2004, 15, 102–112. [Google Scholar] [CrossRef]
- Thunes, K.H.; Midtgaard, F.; Gjerde, I. Diversity of coleoptera of bracket fungus Fomitopsis pinicola in a Norweigian spruce forest. Biodivers. Conserv. 2000, 9, 833–852. [Google Scholar] [CrossRef]
- Benick, L. Pilzkäfer und Käferpilze. Ökologische und statistische Untersuchungen. Acta Zool. Fenn. 1952, 70, 1–250. [Google Scholar]
- Okland, B. Insect fauna compared between six polypore species in a southern Norwegian spruce forest. Fauna Nor. Ser. B 1995, 42, 21–26. [Google Scholar]
- Graves, R.C.; Graves, A.C.F. The Insects and Other Inhabitants of Shelf Fungi in the Southern Blue Ridge Region of Western North Carolina. I. Introduction, Description of the Habitat, and the Host Fungi. Ann. Entomol. Soc. Am. 1966, 59, 381–390. [Google Scholar] [CrossRef]
- Takahashi, K.H.; Tuno, N.; Kagaya, T. Abundance of mycophagous arthropods present on different species of fungi in relation to resource abundance at different spatial scales. Eur. J. Entomol. 2005, 102, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Schigel, D.S.; Niemelä, T.; Kinnunen, J. Polypores of western Finnish Lapland and seasonal dynamics of polypore beetles. Karstenia 2006, 46, 37–64. [Google Scholar] [CrossRef] [Green Version]
- Schigel, D.S. Polypore–beetle associations in Finland. Fenn. Ann. Zool. Fenn. 2011, 48, 319–348. [Google Scholar] [CrossRef]
- Reibnitz, J. Verbreitung und Lebensräume der Baumschwammfresser Südwestdeutschlands (Coleoptera: Cisidae). Mitt. Entomol. Ver. Stuttg. 1999, 34, 1–76. [Google Scholar]
- Stenlid, J. Population structure of Heterobasidion annosum as determined by somatic incompatibility, sexual incompatibility, and isoenzyme patterns. Can. J. Bot. 1985, 63, 2268–2273. [Google Scholar] [CrossRef]
- Piri, T.; Korhonen, K.; Sairanen, A. Occurrence of Heterobasidion annosum in Pure and Mixed Spruce Stands in Southern Finland. Scand. J. For. Res. 1990, 5, 113–125. [Google Scholar] [CrossRef]
- Kadlec, Z.; Stary, P.; Zumr, V. Field evidence for the large pine weevil, Hylobius abietis as a vector of Heterobasidion annosum. Eur. J. For. Pathol. 1992, 22, 316–318. [Google Scholar] [CrossRef]
- Drenkhan, T.; Sibul, I.; Kasanen, R.; Vainio, E.J. Viruses of Heterobasidion parviporum persist within their fungal host during passage through the alimentary tract of Hylobius abietis. For. Pathol. 2013, 43, 317–323. [Google Scholar] [CrossRef]
- Bušs, K. Forest ecosystem classification in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 1997, 51, 204–218. [Google Scholar]
- Crossley, D.A.; Blair, J.M. A high-efficiency, “low-technology” Tullgren-type extractor for soil microarthropods. Agric. Ecosyst. Environ. 1991, 34, 187–192. [Google Scholar] [CrossRef]
- Lindquist, E.E. Remarkable convergence between two taxa of ascid mites (Acari: Mesostigmata) adapted to living in pore tubes of bracket fungi in North America, with description of Mycolaelaps new genus. Can. J. Zool. 1995, 73, 104–128. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, T.; Bolger, T. Fungal fruiting bodies and the structure of fungus-micro-arthropod assemblages. Biol. Environ. 1997, 97, 249–262. [Google Scholar]
- Dunger, W.; Fiedler, H.J. Methoden der Bodenbiologie; Gustav Fischer Verlag: Stuttgart, Germany, 1997. [Google Scholar]
- Unwin, D.M. A key to the families of British Beetles. Field Stud. Counc. 1988, 166, 149–197. [Google Scholar]
- Lompe, A. Käfer Europas. Familir Ciidae. Available online: http://www.coleonet.de/coleo/texte/ciidae.htm (accessed on 13 May 2017).
- Colwell, R.K.; Chang, X.M.; Jing, C. Interpolating, extrapolating and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2015. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. “Package ‘Vegan’ ” Community Ecology Package, Version 2.5–7; 2020. Available online: http://CRAN.R-project.org/package=vegan (accessed on 25 January 2021).
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Makarova, O.L. Gamasid mites (Parasitiformes, Mesostigmata) dwelling in bracket fungi at the pechoro-ilychskii reserve (Republic of Komi). Zool. Zhurnal 2004, 83, 1335–1340. [Google Scholar]
- Hågvar, S.; Amundsen, T.; Økland, B. Mites of the genus Carabodes (Acari, Oribatida) in Norwegian coniferous forests: Occurrence in different soils, vegetation types and polypore hosts. Scand. J. For. Res. 2014, 29, 629–638. [Google Scholar] [CrossRef]
- Negruckii, S.F. Kornevaya Gubka, 2nd ed.; Agropromizdat: Moscow, Russian, 1986. [Google Scholar]
- Vasiliauskas, A.P. Root Fungus and Resistance of Coniferous Forests Ecosystems; Mokslas Publishers: Vilnius, Lithuania, 1989. [Google Scholar]
- Graves, R.C. Ecological observation on the insects and other inhabinants of woody shelf fungi (Basidiomycetes: Polyporaceae) in the Chichago area. Ann. Entomol. Soc. Amer. 1960, 53, 61–78. [Google Scholar] [CrossRef]
- Buxton, P.A. British Diptera associated with fungi. 111: Plies of all families reared from about 150 species of fungi. Entomol. Mon. 1960, 96, 61–94. [Google Scholar]
- Põldmaa, K.; Jürgenstein, S.; Bahram, M.; Teder, T.; Kurina, O. Host diversity and trophic status as determinants of species richness and community composition of fungus gnats. Basic Appl. Ecol. 2015, 16, 46–53. [Google Scholar] [CrossRef]
- Põldmaa, K.; Kaasik, A.; Tammaru, T.; Kurina, O.; Jürgenstein, S.; Teder, T. Polyphagy on unpredictable resources does not exclude host specialization: Insects feeding on mushrooms. Ecology 2016, 97, 2824–2833. [Google Scholar] [CrossRef]
- Brandtberg, P.; Johansson, M.; Seeger, P. Effects of season and urea treatment on infection of stumps of Picea abies by Heterobasidion annosum in stands on former arable land. Scand. J. For. Res. 1996, 11, 261–268. [Google Scholar] [CrossRef]
- Gonthier, P.; Garbelotto, M.; Varese, G.C.; Nicolotti, G. Relative abundance and potential dispersal range of intersterility groups of Heterobasidion annosum in pure and mixed forests. Can. J. Bot. 2001, 79, 1057–1065. [Google Scholar] [CrossRef]
- Vollbrecht, G.; Johansson, U.; Eriksson, H.; Stendlid, J. Butt rot incidence, yield and growth pattern in a tree species experiment in southwestern Sweden. For. Ecol. Manag. 1995, 76.1, 87–93. [Google Scholar] [CrossRef]
- Greig, B.J.W. Field recognition and diagnosis of Heretobasidion annisum. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 35–92. [Google Scholar]
- Schigel, D.S. Collecting and rearing fungivorous Coleoptera. Rev. d’Ecol. 2008, 63, 15–20. [Google Scholar]
- Querner, P. Combining pitfall traps and soil samples to collect Collembola for site scale biodiversity assessments. Appl. Soil Ecol. 2010, 45, 293–297. [Google Scholar] [CrossRef]
- Thorn, S.; Müller, J.; Bässler, C.; Gminder, A.; Brandl, R.; Heibl, C. Host abundance, durability, basidiome form and phylogenetic isolation determine fungivore species richness. Biol. J. Linn. Soc. 2015, 114, 699–708. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
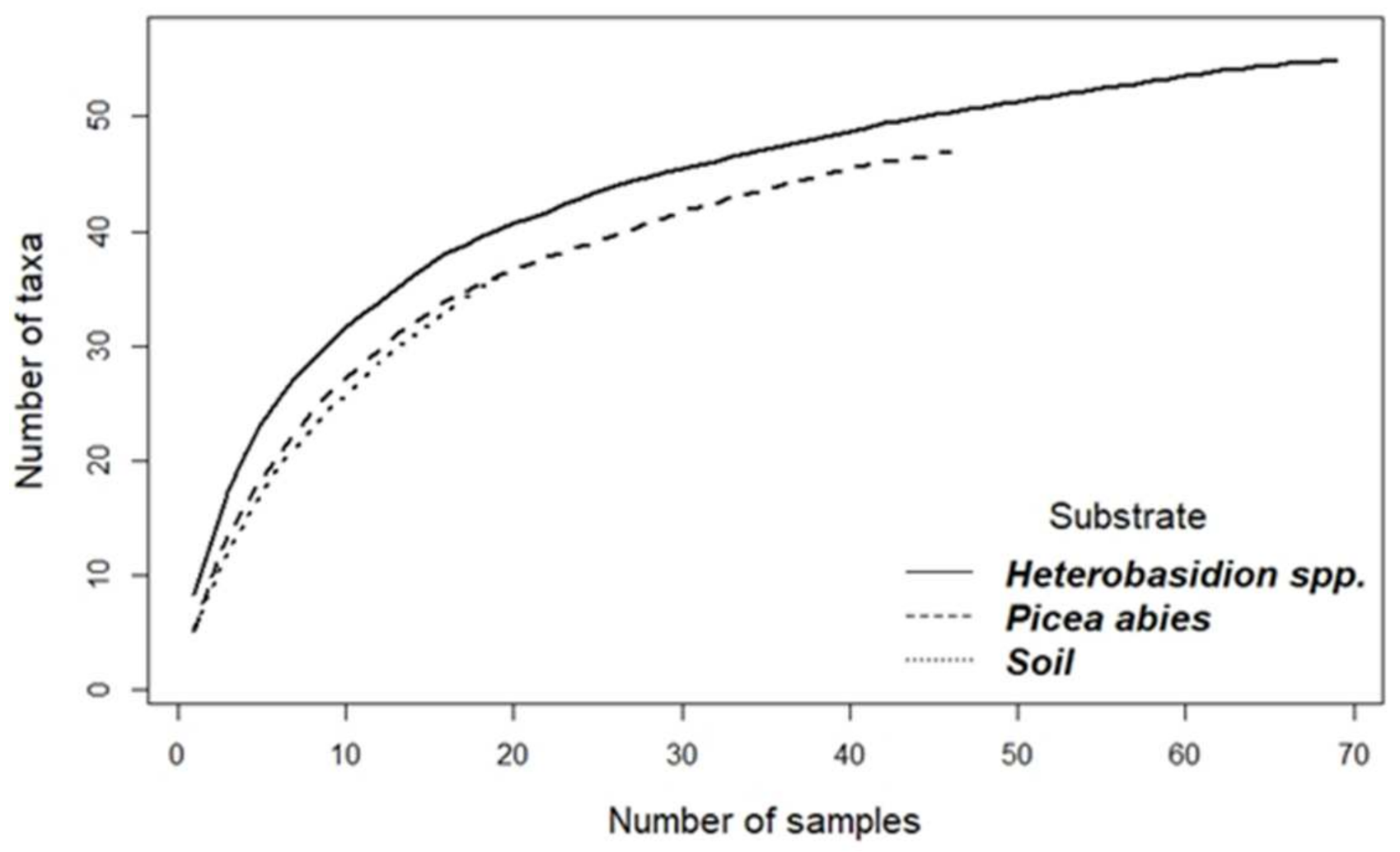{kind=link}
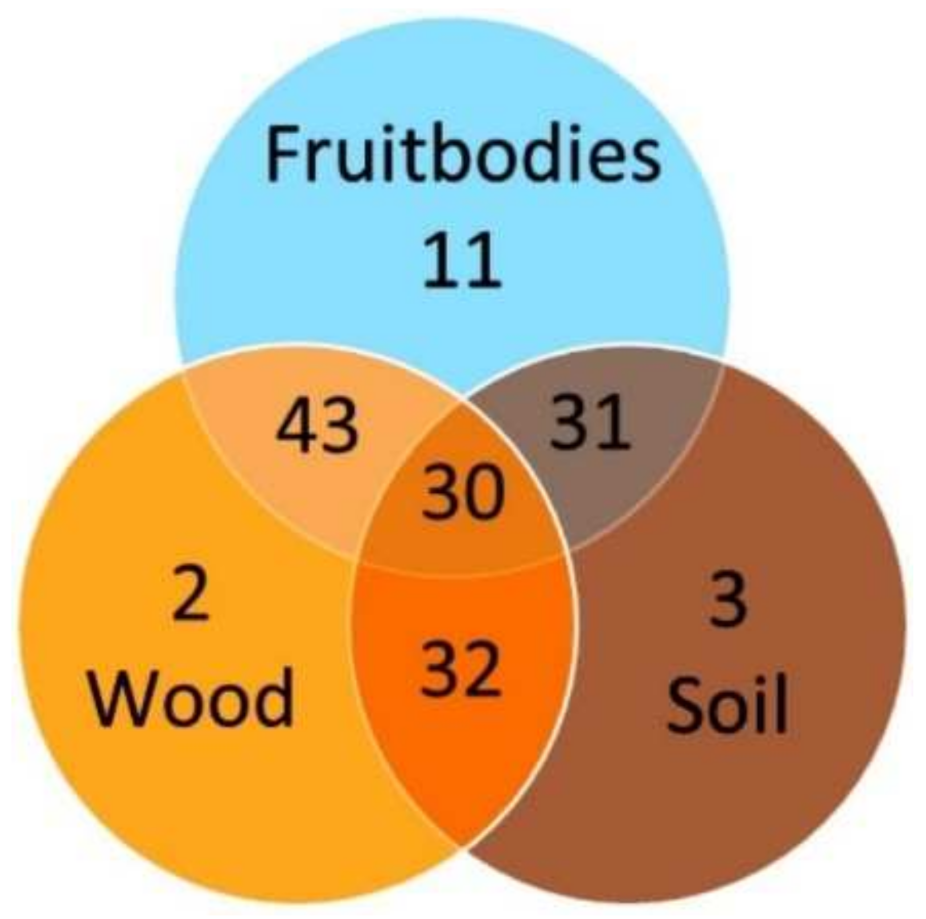{kind=link}
{kind=link}
| Invertebrate Taxa | Heterobasidion | Picea abies Wood | Soil | |||
|---|---|---|---|---|---|---|
| Fruitbodies | ||||||
| Subclass/Family | Adults | Larvae | Adults | Larvae | Adults | Larvae |
| Acari | (4111) | (1277) | (634) | |||
| Hypochthoniidae | 0.6 | - | 2.6 | - | 1.4 | - |
| Pthiracaridae | 0.9 | - | 4.8 | - | 2.2 | - |
| Euphthiracaridae | 0.2 | - | 5.9 | - | 0.3 | - |
| Crotonioidae | 0.7 | - | 0.1 | - | 1.4 | - |
| Nanhermanniidae | 0.1 | - | 0.3 | - | 0.4 | - |
| Damaeidae | 0.2 | - | 0.1 | - | 0.7 | - |
| Eremaeidae | 0.1 | - | 0.1 | - | - | - |
| Tenuialidae | 0.04 | - | 0.1 | - | 0.3 | - |
| Xenillidae | 0.02 | - | 0.2 | - | - | - |
| Carabodidae | 65.9 | - | 12.4 | - | 11.4 | - |
| Oppiidae | 7.3 | - | 39.2 | - | 32.7 | - |
| Scheloribatidae | - | - | - | - | 0.6 | - |
| Ceratozetidae | 0.8 | - | 5.3 | - | 34.9 | - |
| Galumnidae | 0.04 | - | 0.1 | - | 0.6 | - |
| Other 1 | 7.1 | - | 10.3 | - | 4.3 | - |
| Collembola | (309) | (156) | (24) | |||
| Hypogastruridae | 0.2 | - | 0.2 | - | 0.1 | - |
| Neanuridae | 4.2 | - | 6.4 | - | 2.3 | - |
| Onychiuridae | 0.8 | - | 1.5 | - | 0.4 | - |
| Entomobryidae | 0.1 | - | 1.2 | - | 0.3 | - |
| Isotomidae | 0.7 | - | 0.1 | - | 0.3 | - |
| Tomoceridae | 0.2 | - | 0.4 | - | - | - |
| Sminthuridae | 0.02 | - | 0.1 | - | - | - |
| Coleoptera | (170) | (62) | (12) | (29) | (7) | (3) |
| Anobiidae | 0.02 | - | - | - | - | - |
| Cantharidae | - | 17.8 | - | 16.7 | - | 40.0 |
| Carabidae | 0.1 | - | - | - | - | - |
| Ciidae | 3.0 | 11.9 | - | 1.9 | - | - |
| Cryptophagidae | - | - | - | - | 0.1 | - |
| Elateridae | 0.02 | 23.8 | - | 33.3 | - | - |
| Pselaphidae | - | - | 0.1 | - | 0.1 | - |
| Ptilidae | 0.1 | - | 0.5 | - | - | - |
| Silphidae | 0.1 | - | 0.1 | - | - | - |
| Staphylinidae | 0.2 | 7.9 | 0.1 | 1.9 | 0.7 | 20.0 |
| Diplopoda | (126) | (50) | (1) | |||
| Blaniulidae | 2.3 | - | 2.9 | - | 0.1 | - |
| Julidae | 0.02 | - | - | - | - | - |
| Polydesmidae | 0.3 | - | 0.3 | - | - | - |
| Diptera | (50) | (39) | (10) | (22) | (3) | (1) |
| Bibionidae | 0.02 | 0.9 | - | - | - | - |
| Brachycera | 0.1 | 15.8 | - | 7.4 | - | 20.0 |
| Cecidomyiidae | 0.04 | 2.9 | - | - | - | - |
| Ceratopogonidae | 0.1 | - | - | 3.7 | - | - |
| Chiromidae | 0.4 | 14.8 | 0.3 | 25.9 | - | - |
| Drosophilidae | - | - | 0.1 | - | - | - |
| Limoniidae | - | - | - | 3.7 | - | - |
| Psychodidae | 0.02 | - | - | - | - | - |
| Rhagionidae | - | - | - | - | 0.1 | - |
| Sciaridae | 0.3 | 0.9 | 0.3 | - | 0.3 | - |
| Stratiomyidae | 0.02 | 2.9 | - | - | - | - |
| Haplotaxida | (41) | (18) | (8) | |||
| Lumbricidae | 0.4 | - | 0.6 | - | 0.7 | - |
| Enchytraeidae | 0.4 | - | 0.5 | - | 0.4 | - |
| Thysanoptera | 0.7 (35) | - | 0.6 (10) | - | 0.1 (1) | - |
| Hymenoptera | (15) | (7) | (8) | |||
| Parasitica | 0.3 | - | 0.4 | - | 1.2 | - |
| Formicidae | 0.02 | - | - | - | - | - |
| Aranea | 0.2 (10) | - | 0.5 (8) | - | 0.4 (3) | - |
| Gastropoda | (6) | (6) | (1) | |||
| Arionidae | 0.02 | - | - | - | - | - |
| Clausiliidae | 0.02 | - | - | - | 0.1 | - |
| Zonitiidae | 0.1 | - | 0.4 | - | - | - |
| Chilopoda | 0.1 (5) | - | 0.2 (3) | - | 0.1 (1) | - |
| Hemiptera | 0.04 (2) | - | 0.1 (1) | - | 0.3 (2) | - |
| Lepidoptera | (2) | (3) | (1) | |||
| Tineidae | 0.04 | - | - | 5.5 | - | 20.0 |
| Isopoda | (2) | (2) | ||||
| Trichoniscidae | 0.04 | - | 0.1 | - | - | - |
| Opiliones | 0.02 (1) | - | - | - | - | |
| Protura | 0.02 (1) | - | - | - | - | - |
| Pseudoscorpionida | - | - | 0.6 (10) | - | 0.3 (2) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legzdina, L.; Spungis, V.; Burnevica, N.; Gaitnieks, T.; Menkis, A. Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil. Forests 2021, 12, 1100. https://doi.org/10.3390/f12081100
Legzdina L, Spungis V, Burnevica N, Gaitnieks T, Menkis A. Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil. Forests. 2021; 12(8):1100. https://doi.org/10.3390/f12081100
Chicago/Turabian StyleLegzdina, Liva, Voldemars Spungis, Natalija Burnevica, Talis Gaitnieks, and Audrius Menkis. 2021. "Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil" Forests 12, no. 8: 1100. https://doi.org/10.3390/f12081100
APA StyleLegzdina, L., Spungis, V., Burnevica, N., Gaitnieks, T., & Menkis, A. (2021). Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil. Forests, 12(8), 1100. https://doi.org/10.3390/f12081100