
Genome-Wide Characterization of HSP90 Gene Family in Malus sieversii and Their Potential Roles in Response to Valsa mali Infection

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Pathogen Infection
2.2. RNA Isolation and Real Time Quantitative PCR Analysis
2.3. RNA-Seq Data Analysis
2.4. Genome Wide Identification of the MsHSP90 Gens and Gene Sequence Analysis
2.5. Prediction of Transcription Factor Binding Sites of MsHSP90 Genes Promoter
3. Results
3.1. Analysis of Plant Pathogen Interaction Pathway under V. amli Infection
3.2. Identification of HSP90 Gene Family in M. sieversii
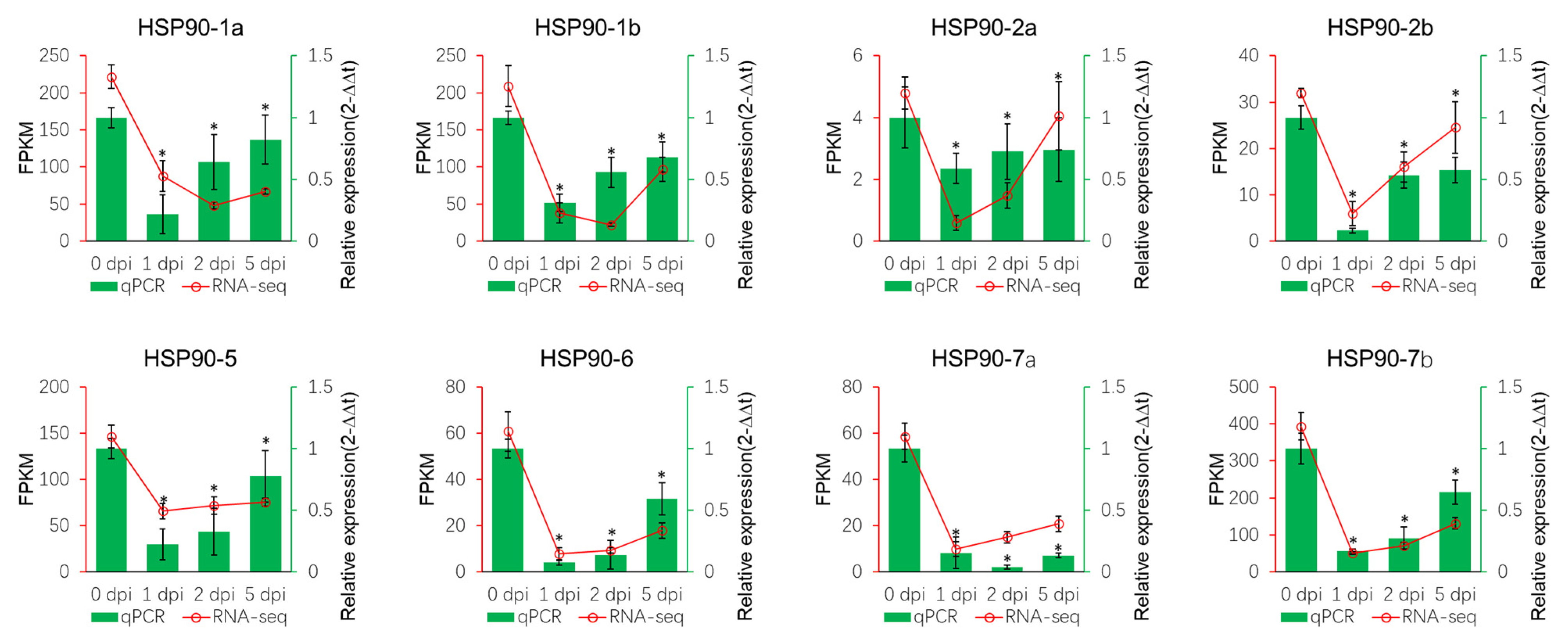
3.3. Expression of MsHSP90 Genes in Response to V.mali
3.4. Prediction of Potential Cis-Elements on HSP90s
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.; Gloor, G. The HSP90 family of proteins in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 238–246. [Google Scholar] [CrossRef]
- Ye, S.; Yu, S.; Shu, L.; Wu, J.; Wu, A.; Luo, L. Expression profile analysis of 9 heat shock protein genes throughout the life cycle and under abiotic stress in rice. Chin. Sci. Bull. 2012, 57, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xue, C.; Xue, D.; Zhao, J.; Gai, J.; Guo, N.; Xing, H. Overexpression of GmHSP90s, a heat shock protein 90 (HSP90) gene family cloning from soybean, decrease damage of abiotic stresses in Arabidopsis thaliana. PLoS ONE 2013, 8, e69810. [Google Scholar] [CrossRef]
- Zhang, K.; He, S.; Sui, Y.; Gao, Q.; Jia, S.; Lu, X.; Jia, L. Genome-Wide Characterization of HSP90 Gene Family in Cucumber and Their Potential Roles in Response to Abiotic and Biotic Stresses. Front. Genet. 2021, 12, 95. [Google Scholar] [CrossRef]
- Zhang, J.; Li, J.; Liu, B.; Zhang, L.; Chen, J.; Lu, M. Genome-wide analysis of the Populus HSP90 gene family reveals differential expression patterns, localization, and heat stress responses. BMC Genom. 2013, 14, 532. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Pan, F.; Yang, C.; Jia, H.; Jiang, H.; He, F.; Li, N.; Lu, X.; Zhang, H. Genome-wide identification and expression analysis of HSP90 gene family in Nicotiana tabacum. BMC Genet. 2019, 20, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.-S.; Li, Z.-Y.; Chen, Y.; Chen, M.; Li, L.-C.; Ma, Y.-Z. Heat Shock Protein 90 in Plants: Molecular Mechanisms and Roles in Stress Responses. Int. J. Mol. Sci. 2012, 13, 15706–15723. [Google Scholar] [CrossRef]
- Corigliano, M.G.; Clemente, M. The Role of Plant 90-kDa Heat Shock Proteins in Plant Immunity. In Heat Shock Proteins and Plants; Asea, A.A.A., Kaur, P., Calderwood, S.K., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 149–167. ISBN 978-3-319-46340-7. [Google Scholar]
- Zai, W.S.; Miao, L.X.; Xiong, Z.L.; Zhang, H.L.; Ma, Y.R.; Li, Y.L.; Chen, Y.B.; Ye, S.G. Comprehensive identification and expression analysis of HSP90s gene family in Solanum lycopersicum. Genet. Mol. Res. 2015, 14, 7811–7820. [Google Scholar] [CrossRef]
- Prasad, B.D.; Goel, S.; Krishna, P. In silico identification of carboxylate clamp type tetratricopeptide repeat proteins in Arabidopsis and rice as putative co-chaperones of HSP90/HSP70. PLoS ONE 2010, 5, e12761. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.C.; Grover, A. Constitutive over-expression of rice ClpD1 protein enhances tolerance to salt and desiccation stresses in transgenic Arabidopsis plants. Plant Sci. 2016, 250, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Vítámvás, P.; Prášil, I.T.; Kosová, K.; Planchon, S.; Renaut, J. Analysis of proteome and frost tolerance in chromosome 5A and 5B reciprocal substitution lines between two winter wheats during long-term cold acclimation. Proteomics 2012, 12, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Schöffl, F.; Prändl, R.; Reindl, A. Regulation of the Heat-Shock Response. Plant Physiol. 1998, 117, 1135–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasinos, C.; Krampis, K.; Samakovli, D.; Hatzopoulos, P. Tight regulation of expression of two Arabidopsis cytosolic HSP90 genes during embryo development. J. Exp. Bot. 2005, 56, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, C.; Eggers-Schumacher, G.; Wunderlich, M.; Schöffl, F. Two different heat shock transcription factors regulate immediate early expression of stress genes in Arabidopsis. Mol. Genet. Genom. 2004, 271, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Guo, H.; Yuan, E.; Ge, F. Elevated CO2 increases R gene-dependent resistance of Medicago truncatula against the pea aphid by up-regulating a heat shock gene. New Phytol. 2018, 217, 1696–1711. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Ohnishi, K.; Hikichi, Y.; Kiba, A. Molecular chaperons and co-chaperons, HSP90, RAR1, and SGT1 negatively regulate bacterial wilt disease caused by Ralstonia solanacearum in Nicotiana benthamiana. Plant Signal. Behav. 2015, 10, e970410. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yang, Y.; Wang, C.; Liu, M.; Li, H.; Fu, Y.; Wang, Y.; Nie, Y.; Liu, X.; Ji, W. Large-scale transcriptome comparison reveals distinct gene activations in wheat responding to stripe rust and powdery mildew. BMC Genom. 2014, 15, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, N.; Bai, Y.; Sun, H.; Wang, N.; Ma, Y.; Li, M.; Wang, X.; Jiao, C.; Legall, N.; Mao, L.; et al. Genome re-sequencing reveals the history of apple and supports a two-stage model for fruit enlargement. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.-R.; Norelli, J.; Burchard, E.; Abdelfattah, A.; Levin, E.; González-Candelas, L.; Droby, S.; Wisniewski, M. Transcriptomic Response of Resistant (PI613981–Malus sieversii) and Susceptible (“Royal Gala”) Genotypes of Apple to Blue Mold (Penicillium expansum) Infection. Front. Plant Sci. 2017, 8, 1981. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Bozorov, T.A.; Ma, R.; Ma, J.; Zhang, Y.; Yang, H.; Li, L.; Zhang, D. Characterization and pathogenicity of six Cytospora strains causing stem canker of wild apple in the Tianshan Forest, China. For. Pathol. 2020, 50, e12587. [Google Scholar] [CrossRef]
- Bozorov, T.A.; Luo, Z.; Li, X.; Zhang, D. Agrilus mali Matsumara (Coleoptera: Buprestidae), a new invasive pest of wild apple in western China: DNA barcoding and life cycle. Ecol. Evol. 2019, 9, 1160–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soosaar, J.L.M.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, C.; You, Y.; Liang, W.; Wang, N.; MA, F.; Li, C. Validation of reference genes for qRT-PCR analysis in peel and flesh of six apple cultivars (Malus domestica) at diverse stages of fruit development. Sci. Hortic. 2019, 244, 165–171. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Wen, X.; Zhang, Y.; Ding, Y.; Zhang, Y.; Gao, B.; Zhang, D. PacBio full-length transcriptome of wild apple (Malus sieversii) provides insights into canker disease dynamic response. BMC Genom. 2021, 22, 1–19. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2019, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Hu, G.; Han, B. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice. Plant Sci. 2009, 176, 583–590. [Google Scholar] [CrossRef]
- Daccord, N.; Celton, J.-M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Weichen, N.; Liu, S.; Cai, B.; Xing, H.; Wang, S. Transcriptomics Analysis of Apple Leaves in Response to Alternaria alternata Apple Pathotype Infection. Front. Plant Sci. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genom. 2007, 8, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Burch-Smith, T.; Schiff, M.; Feng, S.; Dinesh-Kumar, S.P. Molecular Chaperone HSP90 Associates with Resistance Protein N and Its Signaling Proteins SGT1 and Rar1 to Modulate an Innate Immune Response in Plants. J. Biol. Chem. 2004, 279, 2101–2108. [Google Scholar] [CrossRef]
- Takahashi, A.; Casais, C.; Ichimura, K.; Shirasu, K. HSP90 interacts with RAR1 and SGT1 and is essential for RPS2-mediated disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 11777–11782. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.; Zhao, J.; Du, Y.; Zhao, X.; Han, M.; Liu, Y. HSP90 Interacts With Tm-22 and Is Essential for Tm-22-Mediated Resistance to Tobacco mosaic virus. Front. Plant Sci. 2018, 9, 411. [Google Scholar] [CrossRef] [Green Version]
- Shirasu, K. The HSP90-SGT1 Chaperone Complex for NLR Immune Sensors. Annu. Rev. Plant Biol. 2009, 60, 139–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, C.; Betsuyaku, S.; Peart, J.; Takahashi, A.; Noël, L.; Sadanandom, A.; Casais, C.; Parker, J.; Shirasu, K. Role of SGT1 in resistance protein accumulation in plant immunity. EMBO J. 2006, 25, 2007–2016. [Google Scholar] [CrossRef] [Green Version]
- Austin, M.J.; Muskett, P.; Kahn, K.; Feys, B.J.; Jones, J.D.G.; Parker, J.E. Regulatory Role of SGT1 in Early R Gene-Mediated Plant Defenses. Science 2002, 295, 2077–2080. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Length of CDS (bp) | Chromosomes | Genomic Coordinates | Strand | Number of Amino Acids in Protein |
|---|---|---|---|---|---|
| MdHSP90-1a | 2112 | 7 | NC_041795.1: 34410217-34414101 | + | 703 |
| MdHSP90-1b | 2112 | 1 | NC_041789.1: 30385625-30388837 | + | 703 |
| MdHSP90-2a | 2100 | 11 | NC_041799.1: 3210146-3213261 | − | 699 |
| MdHSP90-2b | 2100 | 3 | NC_041791.1: 2915642-2918677 | − | 699 |
| MdHSP90-5 | 2385 | 16 | NC_041804.1: 3808810-3813939 | − | 794 |
| MdHSP90-6a | 2421 | 17 | NC_041805.1: 9778825-9785023 | − | 806 |
| MdHSP90-6b | 2430 | 9 | NC_041797.1: 9428843-9434534 | − | 809 |
| MdHSP90-7 | 2457 | 8 | NC_041796.1: 866442-871008 | + | 818 |
| MsHSP90-1a | 2490 | 9 | Chr9:35058827-35061327 | + | 829 |
| MsHSP90-1b | 1512 | 13 | Chr13:32200739-32203239 | + | 503 |
| MsHSP90-2a | 2100 | 5 | Chr5:3353408-3350908 | − | 699 |
| MsHSP90-2b | 2085 | 8 | Chr8:2754830-2752330 | − | 694 |
| MsHSP90-5 | 2385 | 6 | Chr6:3862971-3860471 | − | 794 |
| MsHSP90-6 | 2415 | 10 | Chr10:27515958-27518458 | + | 804 |
| MsHSP90-7a | 2457 | 1 | Chr1:622597-625097 | + | 818 |
| MsHSP90-7b | 2457 | 15 | Chr15:32199616-32197116 | − | 818 |
| Gene Name | Start | End | p-Value | q-Value | Matched Sequence |
|---|---|---|---|---|---|
| MsHSP90-1a | 40 | 49 | 7.01 × 10−5 | 0.672 | --GTTCACGAAC-- |
| MsHSP90-1b | 2139 | 2148 | 1.19 × 10−5 | 0.232 | --GGAACGTTCC-- |
| 2176 | 2185 | 2.63 × 10−4 | 0.509 | --AGAACATTCT-- | |
| 2435 | 2444 | 2.63 × 10−4 | 0.509 | --AGAAAATTCG-- | |
| 2457 | 2466 | 2.63 × 10−4 | 0.509 | --CGAATTTTCT-- | |
| MsHSP90-2a | 194 | 203 | 1.76 × 10−4 | 0.509 | --CGAAAATTCC-- |
| 1754 | 1763 | 1.76 × 10−4 | 0.509 | --AGAACGTTCA-- | |
| 445 | 454 | 2.63 × 10−4 | 0.509 | --AGAAATTTCG-- | |
| 1447 | 1456 | 2.89 × 10−4 | 0.672 | --TTTCATGAAA-- | |
| MsHSP90-2b | 117 | 126 | 1.76 × 10−4 | 0.509 | --GGAATTTTCG-- |
| MsHSP90-5 | 1301 | 1310 | 2.89 × 10−4 | 0.509 | --TGAATTTTCT-- |
| 2244 | 2253 | 1.76 × 10−4 | 0.672 | --GTTCCAGAAA-- | |
| 2290 | 2299 | 1.76 × 10−4 | 0.672 | --TTTCCAGAAG-- | |
| MsHSP90-7a | 1540 | 1549 | 2.63 × 10−4 | 0.509 | --TGAACTTTCA-- |
| MsHSP90-7b | 1182 | 1191 | 1.76 × 10−4 | 0.509 | --GGAAATTTCG-- |
| 2372 | 2381 | 1.76 × 10−4 | 0.672 | --TTTCTGGAAC-- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haxim, Y.; Si, Y.; Liu, X.; Wen, X.; Kahar, G.; Ding, Y.; Li, X.; Zhang, D. Genome-Wide Characterization of HSP90 Gene Family in Malus sieversii and Their Potential Roles in Response to Valsa mali Infection. Forests 2021, 12, 1232. https://doi.org/10.3390/f12091232
Haxim Y, Si Y, Liu X, Wen X, Kahar G, Ding Y, Li X, Zhang D. Genome-Wide Characterization of HSP90 Gene Family in Malus sieversii and Their Potential Roles in Response to Valsa mali Infection. Forests. 2021; 12(9):1232. https://doi.org/10.3390/f12091232
Chicago/Turabian StyleHaxim, Yakupjan, Yu Si, Xiaojie Liu, Xuejing Wen, Gulnaz Kahar, Yu Ding, Xiaoshuang Li, and Daoyuan Zhang. 2021. "Genome-Wide Characterization of HSP90 Gene Family in Malus sieversii and Their Potential Roles in Response to Valsa mali Infection" Forests 12, no. 9: 1232. https://doi.org/10.3390/f12091232
APA StyleHaxim, Y., Si, Y., Liu, X., Wen, X., Kahar, G., Ding, Y., Li, X., & Zhang, D. (2021). Genome-Wide Characterization of HSP90 Gene Family in Malus sieversii and Their Potential Roles in Response to Valsa mali Infection. Forests, 12(9), 1232. https://doi.org/10.3390/f12091232