
Investigation of the Architecture and Age of Superlative Adansonia grandidieri from the Andombiry Forest, Madagascar

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
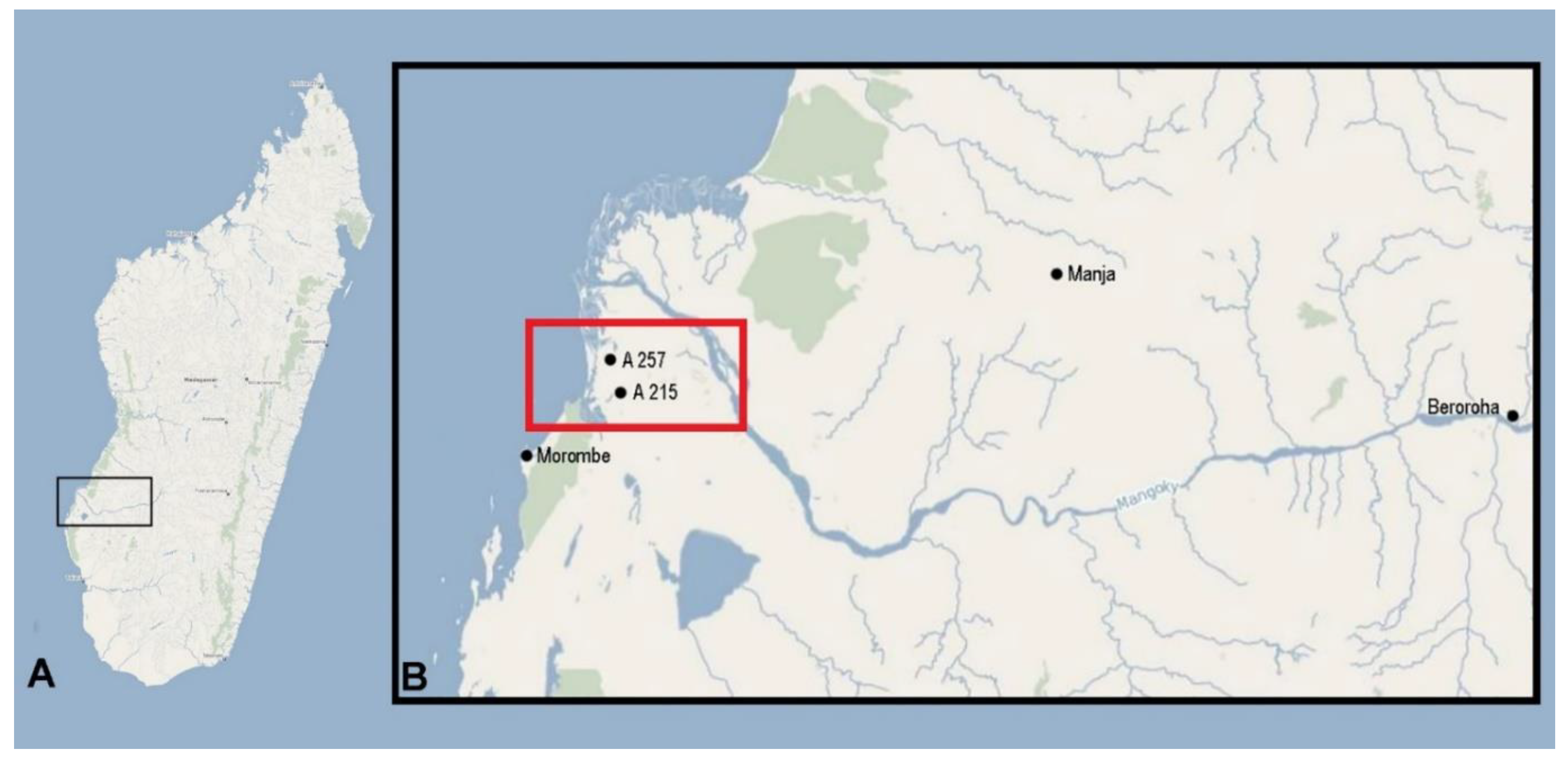
2.1. The Two Baobabs and Their Areas
2.2. Investigation of the Two Baobabs
2.2.1. Wood Samples
2.2.2. Sample Preparation
2.2.3. AMS Investigation
2.2.4. Calibration
3. Results
3.1. Radiocarbon Dates and Calibrated Ages
3.2. Sample Ages
4. Discussion
4.1. Architecture of the Two Baobabs
4.2. Age of the Two Baobabs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wickens, G.E. The Baobab—Africa’s Upside-Down Tree. Kew Bull. 1982, 37, 172–209. [Google Scholar] [CrossRef]
- Baum, D.A. A Systematic Revision of Adansonia (Bombacaceae). Ann. Mo. Bot. Gard. 1995, 82, 440. [Google Scholar] [CrossRef]
- Baum, D.A.; Small, R.L.; Wendel, J.F. Biogeography and Floral Evolution of Baobabs Adansonia, Bombacaceae as Inferred from Multiple Data Sets. Syst. Biol. 1998, 47, 181–207. [Google Scholar] [CrossRef] [PubMed]
- Wickens, G.E.; Lowe, P. The Baobabs: Pachycauls of Africa, Madagascar and Australia; Springer: Dordrecht, The Netherlands, 2008; 498 p, ISBN 978-1-4020-6431-9. [Google Scholar]
- Frsjd, P.; Bell, K.L.; Bhagwandin, A.; Grinan, E.; Jillani, N.; Meyer, J.; Wabuyele, E.; Vickers, C.E. Morphology, ploidy and molecular phylogenetics reveal a new diploid species from Africa in the baobab genus Adansonia (Malvaceae: Bombacoideae). Taxon 2012, 61, 1240–1250. [Google Scholar] [CrossRef]
- Cron, G.V.; Karimi, N.; Glennon, K.L.; Udeh, C.A.; Witkowski, E.T.; Venter, S.M.; Assogbadjo, A.E.; Mayne, D.H.; Baum, D.A. One African baobab species or two? Synonymy of Adansonia kilima and A. digitata. Taxon 2016, 65, 1462. [Google Scholar] [CrossRef] [Green Version]
- Petignat, A.; Jasper, L. Baobabs of the World: The Upside-Down Trees of Madagascar, Africa and Australia; Struik Publishers: Cape Town, South Africa, 2016; 112 p, ISBN 9781775843702. [Google Scholar]
- Cornu, C.; Danthu, P. Guide D’identification Illustré des Baobabs de Madagascar; CIRAD: Montpellier, France, 2015; 30 p. [Google Scholar]
- Patrut, A.; Von Reden, K.F.; Lowy, D.; Alberts, A.H.; Pohlman, J.W.; Wittmann, R.; Gerlach, D.S.; Xu, L.; Mitchell, C.S. Radiocarbon dating of a very large African baobab. Tree Physiol. 2007, 27, 1569–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrut, A.; Von Reden, K.F.; Van Pelt, R.; Mayne, D.H.; Lowy, D.; Margineanu, D. Age determination of large live trees with inner cavities: Radiocarbon dating of Platland tree, a giant African baobab. Ann. For. Sci. 2011, 68, 993–1003. [Google Scholar] [CrossRef]
- Patrut, A.; von Reden, K.F.; Mayne, D.H.; Lowy, D.A.; Patrut, R.T. AMS radiocarbon investigation of the African baobab: Searching for the oldest tree. Nucl. Instr. Meth. B 2013, 294, 622–626. [Google Scholar] [CrossRef]
- Patrut, A.; Woodborne, S.; Von Reden, K.F.; Hall, G.; Hofmeyr, M.; Lowy, D.A.; Patrut, R. African Baobabs with False Inner Cavities: The Radiocarbon Investigation of the Lebombo Eco Trail Baobab. PLoS ONE 2015, 10, e0117193. [Google Scholar] [CrossRef]
- Patrut, A.; Woodborne, S.; von Reden, K.F.; Hall, G.; Patrut, R.T.; Rakosy, L.; Danthu, P.; Pock-Tsy, J.-M.L.; Lowy, D.A.; Margineanu, D. The Growth Stop Phenomenon of Baobabs (Adansonia spp.) Identified by Radiocarbon Dating. Radiocarbon 2017, 59, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Patrut, A.; Woodborne, S.; Patrut, R.; Rakosy, L.; Lowy, D.A.; Hall, G.; Von Reden, K.F. The demise of the largest and oldest African baobabs. Nat. Plants 2018, 4, 423–426. [Google Scholar] [CrossRef]
- Patrut, A.; Woodborne, S.; Patrut, R.T.; Hall, G.; Rakosy, L.; Winterbach, C.; Von Reden, K.F. Age, Growth and Death of a National Icon: The Historic Chapman Baobab of Botswana. Forests 2019, 10, 983. [Google Scholar] [CrossRef] [Green Version]
- Patrut, A.; Garg, A.; Woodborne, S.; Patrut, R.T.; Rakosy, L.; Ratiu, A.; Lowy, D.A. Radiocarbon dating of two old African baobabs from India. PLoS ONE 2020, 15, e0227352. [Google Scholar] [CrossRef] [Green Version]
- Patrut, A.; Von Reden, K.F.; Danthu, P.; Pock-Tsy, J.-M.L.; Patrut, R.; Lowy, D.A. Searching for the Oldest Baobab of Madagascar: Radiocarbon Investigation of Large Adansonia rubrostipa Trees. PLoS ONE 2015, 10, e0121170. [Google Scholar] [CrossRef] [Green Version]
- Patrut, A.; von Reden, K.F.; Danthu, P.; Pock-Tsy, J.M.L.; Rakosy, L.; Patrut, R.T.; Lowy, D.A.; Margineanu, D. AMS radi-ocarbon dating of very large Grandidier’s baobabs (Adansonia grandidieri). Nucl. Instr. Meth. B 2015, 361, 591–598. [Google Scholar] [CrossRef]
- Patrut, A.; Patrut, R.; Danthu, P.; Pock-Tsy, J.-M.L.; Rákosy, L.; Lowy, D.A.; Von Reden, K.F. AMS Radiocarbon Dating of Large Za Baobabs (Adansonia za) of Madagascar. PLoS ONE 2016, 11, e0146977. [Google Scholar] [CrossRef] [Green Version]
- Patrut, A.; Patrut, R.T.; Pock-Tsy, J.-M.L.; Woodborne, S.; Rakosy, L.; Ratiu, I.-A.; Bodis, J.; Danthu, P. Radiocarbon dating of a very large grandidier baobab, the giant of Bevoay. Studia UBB Chem. 2020, 65, 151–158. [Google Scholar] [CrossRef]
- Patrut, R.; Patrut, A.; Pock-Tsy, J.-M.L.; Woodborne, S.; Rakosy, L.; Danthu, P.; Ratiu, A.; Bodis, J.; Von Reden, K.; Biodiversité, A.D.F.E.; et al. Radiocarbon investigation of a superlative grandidier baobab, the big reniala of Isosa. Studia UBB Chem. 2019, 64, 131–139. [Google Scholar] [CrossRef]
- Loader, N.; Robertson, I.; Barker, A.; Switsur, V.; Waterhouse, J. An improved technique for the batch processing of small wholewood samples to α-cellulose. Chem. Geol. 1997, 136, 313–317. [Google Scholar] [CrossRef]
- Sofer, Z. Preparation of carbon dioxide for stable carbon isotope analysis of petroleum fractions. Anal. Chem. 1980, 52, 1389–1391. [Google Scholar] [CrossRef]
- Vogel, J.; Southon, J.; Nelson, D.; Brown, T. Performance of catalytically condensed carbon for use in accelerator mass spectrometry. Nucl. Instr. Meth. B 1984, 5, 289–293. [Google Scholar] [CrossRef]
- Mbele, V.L.; Mullins, S.M.; Winkler, S.R.; Woodborne, S. Acceptance Tests for AMS Radiocarbon Measurements at iThemba LABS, Gauteng, South Africa. Phys. Proc. 2017, 90, 10–16. [Google Scholar] [CrossRef]
- Ramsey, C.B. Bayesian Analysis of Radiocarbon Dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef] [Green Version]
- Hogg, A.G.; Heaton, T.J.; Hua, Q.; Palmer, J.G.; Turney, C.S.; Southon, J.; Bayliss, A.; Blackwell, P.G.; Boswijk, G.; Ramsey, C.B.; et al. SHCal20 Southern Hemisphere Calibration, 0–55,000 Years cal BP. Radiocarbon 2020, 62, 759–778. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Higuchi, N.; Schimel, J.P. Ancient trees in Amazonia. Nature 1998, 391, 135–136. [Google Scholar] [CrossRef]
- Worbes, M.; Junk, W.J. How Old are Tropical Trees? the Persistence of a Myth. IAWA J. 1999, 20, 255–260. [Google Scholar] [CrossRef]
- Williamson, G.; Van Eldik, T.; Delamônica, P.; Laurance, W.F. How many millenarians in Amazonia? Sizing the ages of large trees. Trends Plant Sci. 1999, 4, 387. [Google Scholar] [CrossRef]
- Fichtler, E.; Clark, D.A.; Worbes, M. Age and Long-term Growth of Trees in an Old-growth Tropical Rain Forest, Based on Analyses of Tree Rings and 14C1. Biotropica 2003, 35, 306–317. [Google Scholar] [CrossRef]
- Kinahan, J. Human Responses to Climatic Variation in the Namib Desert during the Last 1000 Years. Afr. Archaeol. Rev. 2016, 33, 183–203. [Google Scholar] [CrossRef]
- Ehrlich, Y.; Regev, L.; Kerem, Z.; Boaretto, E. Radiocarbon Dating of an Olive Tree Cross-Section: New Insights on Growth Patterns and Implications for Age Estimation of Olive Trees. Front. Plant Sci. 2017, 8, 1918. [Google Scholar] [CrossRef]
- Chen, J.; Shen, H.; Sasa, K.; Lan, H.; Matsunaka, T.; Matsumura, M.; Takahashi, T.; Hosoya, S.; He, M.; He, Y.; et al. Radiocarbon dating of Chinese Ancient Tea Trees. Radiocarbon 2019, 61, 1741–1748. [Google Scholar] [CrossRef]
- Garg, A.; Patrut, R.T.; Patrut, A.; Woodborne, S.; Rakosy, L. Radiocarbon dating and status of the oldest extant Ceylon iron wood (Manilkara hexandra) in the riverine Ramsar site of India. Curr. Sci. 2021, 120, 562–566. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample/ Segment Code | Depth 1 (Height 2) (m) | Radiocarbon Date (Error) (14C yr BP) | Cal CE Range 1σ (Confidence Interval) | Assigned Year (Error) (cal CE) | Sample/Segment Age (Error) (cal CE) | Accession Number |
|---|---|---|---|---|---|---|
| A 257-1a | 0.10 (1.33) | 98 (±40) | 1708–1720 (6.7%) 1812–1836 (16.8%) 1852–1866 (7.5%) 1880–1926 (37.3%) | 1903 (±25) | 120 (±25) | IT-C-2229 |
| A 257-1b | 0.20 (1.33) | 207 (±24) | 1671–1688 (15.0%) 1730–1782 (43.9%) 1792–1802 (9.4%) | 1756 (±25) | 265 (±25) | IT-C-2693 |
| A 257-1c | 0.35 (1.33) | 361 (±23) | 1505–1518 (9.8%) 1524–1592 (50.6%) 1618–1628 (7.8%) | 1558 (±35) | 465 (±35) | IT-C-2699 |
| A 257-1d | 0.50 (1.33) | 477 (±39) | 1426–1484 (68.3%) | 1455 (±30) | 565 (±30) | IT-C-2215 |
| A 257-1e | 0.60 (1.33) | 625 (±22) | 1324–1346 (45.4%) 1391–1402 (22.8%) | 1335 (±10) | 685 (±10) | IT-C-2705 |
| A 257-1f | 0.75 (1.33) | 923 (±30) | 1152–1214 (68.3%) | 1183 (±20) | 860 (±20) | IT-C-1682 |
| A 215-2a | 0.20 (1.40) | 126 (±23) | 1710–1718 (6.0%) 1813–1836 (18.6%) 1855–1866 (6.5%) 1880–1925 (37.2%) | 1902 (±20) | 120 (±20) | IT-C-2701 |
| A 215-2b | 0.45 (1.40) | 220 (±27) | 1668–1680 (10.1%) 1734–1785 (49.4%) 1792–1802 (8.7%) | 1770 (±25) | 250 (±25) | IT-C-2704 |
| A 215-2c | 0.65 (1.40) | 269 (±30) | 1640–1672 (40.6%) 1743–1763 (6.4%) 1765–1772 (4.8%) 1780–1797 (16.4%) | 1656 (±15) | 365 (±15) | IT-C-2230 |
| A 215-2d | 0.78 (1.40) | 206 (±29) | 1669–1692 (16.7%) 1727–1784 (41.3%) 1794–1808 (10.3%) | 1755 (±30) | 265 (±30) | IT-C-2711 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patrut, A.; Patrut, R.T.; Leong Pock-Tsy, J.-M.; Danthu, P.; Woodborne, S.; Rakosy, L.; Ratiu, I.A. Investigation of the Architecture and Age of Superlative Adansonia grandidieri from the Andombiry Forest, Madagascar. Forests 2021, 12, 1258. https://doi.org/10.3390/f12091258
Patrut A, Patrut RT, Leong Pock-Tsy J-M, Danthu P, Woodborne S, Rakosy L, Ratiu IA. Investigation of the Architecture and Age of Superlative Adansonia grandidieri from the Andombiry Forest, Madagascar. Forests. 2021; 12(9):1258. https://doi.org/10.3390/f12091258
Chicago/Turabian StylePatrut, Adrian, Roxana Teodora Patrut, Jean-Michel Leong Pock-Tsy, Pascal Danthu, Stephan Woodborne, Laszlo Rakosy, and Ileana Andreea Ratiu. 2021. "Investigation of the Architecture and Age of Superlative Adansonia grandidieri from the Andombiry Forest, Madagascar" Forests 12, no. 9: 1258. https://doi.org/10.3390/f12091258
APA StylePatrut, A., Patrut, R. T., Leong Pock-Tsy, J. -M., Danthu, P., Woodborne, S., Rakosy, L., & Ratiu, I. A. (2021). Investigation of the Architecture and Age of Superlative Adansonia grandidieri from the Andombiry Forest, Madagascar. Forests, 12(9), 1258. https://doi.org/10.3390/f12091258