
High Level of Phenotypic Differentiation of Common Yew (Taxus baccata L.) Populations in the North-Western Part of the Balkan Peninsula

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
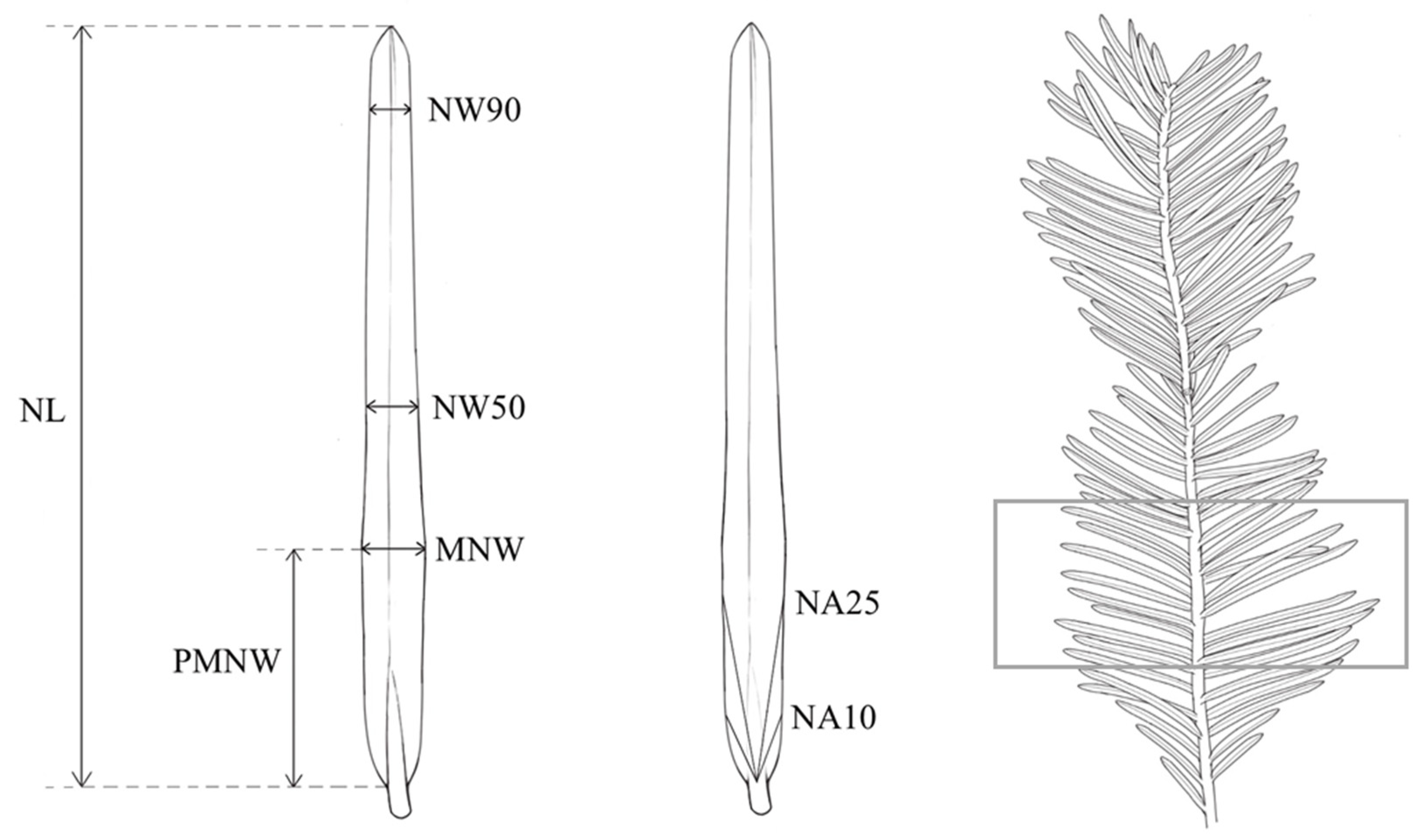
2.2. Morphometric Analysis
2.3. Statistical Analysis
3. Results
3.1. Needle Morphometric Traits and Population Variability
3.2. Population Structure
3.3. Isolation by Distance (IBD) and Environment (IBE)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Farjon, A.; Filer, D. An Atlas of the World’s Conifers; Brill Academic Publication: Leiden, The Netherlands; Boston, MA, USA, 2013; p. 525. [Google Scholar]
- Gonzalez-Martinez, S.C.; Dubreuil, M.; Ribva, M.; Vendramin, G.G.; Sebastiani, F.; Mayol, M. Spatial genetic structure of Taxus baccata L. in the western Mediterranean Basin: Past and present limits to gene movement over a broad geographic scale. Mol. Phylogenet. Evol. 2010, 55, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.A.; Polwart, A. Biological flora of the British Isles: Taxus baccata L. J. Ecol. 2003, 91, 489–524. [Google Scholar] [CrossRef]
- Benham, S.E.; Houston Durrant, T.; Caudullo, G.; de Rigo, D. Taxus baccata in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; p. e015921. [Google Scholar]
- Pietzarka, U. Zur ökologischen Strategie der Eibe (Taxus baccata L.) Wachstums- und Verjüngungsdynamik. Ph.D. Thesis, Technisches Universität Dresden, Dresden, Germany, 2005. [Google Scholar]
- Hageneder, F. Yew; Reaktion Books: London, UK, 2013; p. 224. [Google Scholar]
- Barwise, Y.; Kumar, P. Designing vegetation barriers for urban air pollution abatement: A practical review for appropriate plant species selection. Clim. Atmos. Sci. 2020, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Hageneder, F. Appendix III: Important occurrences of European yew (Taxus baccata L.). In Yew: A History, 2nd ed.; the History Press Ltd.: Stroud, UK, 2011; p. 10. [Google Scholar]
- Mayol, M.; Riba, M.; Gonzalez-Martinez, S.C.; Bagnoli, F.; de Beaulieu, J.; Breganzo, E.; Burgarella, C.; Dubreuil, M.; Kraj-merova, D.; Paule, L.; et al. Adapting through glacial cycles: Insights from a long-lived tree (Taxus baccata). New Phytol. 2015, 208, 973–986. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.; Cusidó, R.M.; Hossein Mirjalili, M.; Moyano, E.; Palazón, J.; Bonfill, M. Production of the anticancer drug taxol in Taxus baccata suspension cultures: A review. Proc. Biochem. 2011, 46, 23–34. [Google Scholar] [CrossRef]
- Stefanović, M.; Ristić, M.; Popović, Z.; Matić, R.; Nikolić, B.; Vidaković, V.; Oratov-Petković, D.; Bojović, S. Chemical composition and interpopulation variability of essential oils of Taxus baccata L. from Serbia. Chem. Biodivers. 2016, 13, 943–953. [Google Scholar] [CrossRef]
- Iszkuło, G. Success and failure of endangered tree species: Low temperatures and low light availability affect survival and growth of European yew (Taxus baccata L.) seedlings. Pol. J. Ecol. 2010, 58, 259–271. [Google Scholar]
- Iszkuło, G.; Didukh, Y.; Giertych, J.; Jasinska, A.K.; Sobierajska, K.; Szmyt, J. Weak competitive ability may explain decline of Taxus baccata. Ann. For. Sci. 2012, 69, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Rivers, M.; Beech, E.; Bazos, I.; Bogunić, F.; Buira, A.; Caković, D.; Carapeto, A.; Carta, A.; Cornier, B.; Fenu, G.; et al. European Red List of Trees, 1st ed.; IUCN: Cambridge, UK; Brussels, Belgium, 2019; p. 72. [Google Scholar]
- Vukelić, J. Šumska Vegetacija Hrvatske, 1st ed.; University of Zagreb, Faculty of Forestry, Institute for Nature Protection: Zagreb, Croatia, 2012; pp. 191–193. [Google Scholar]
- Ramírez-Valiente, J.; Sánchez-Gómez, D.; Aranda, I.; Vallardes, F. Phenotypic plasticity and local adaptation in leaf ecophysiological traits of 13 contrasting cork oak populations under different water availabilities. Tree Phys. 2010, 30, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Regent Instruments Inc. WinFolia TM; Version PRO 2005b; Regent Instruments Inc.: Quebec, QC, Canada, 2001.
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W.H. Freeman and Co.: New York, NY, USA, 2012; p. 937. [Google Scholar]
- StatSoft, Inc. STATISTICA (Data Analysis Software System); version 13; StatSoft, Inc.: Tulsa, OK, USA, 2018.
- McGarigal, K.; Cushman, S.; Stafford, S. Multivariate Statistics for Wildlife and Ecology Research; Springer: New York, NY, USA, 2000; p. 283. [Google Scholar]
- Douaihy, B.; Sobierajska, K.; Jasinska, A.K.; Boratynska, K.; Ok, T.; Romo, A.; Machon, N.; Didukh, Y.; Dagher-Kharrat, M.B.; Boratynski, A. Morphological versus molecular markers to describe variability in Juniperus excelsa subsp. excelsa (Cupressaceae). AoB Plants 2012, 2012, pls13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boratynski, A.; Jasinska, A.K.; Marcysiak, K.; Mazur, M.; Romo, A.M.; Boratynska, K.; Sobierajska, K.; Iszkuło, G. Morphological differentiation supports the genetic pattern of the geographic structure of Juniperus thurifera (Cupressaceae). Plant Syst. Evol. 2013, 299, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Sobierajska, K.; Boratynska, K.; Jasinska, A.; Dering, M.; Ok, T.; Douaihy, B.; Dagher-Kharrat, M.B.; Romo, A.; Boratynski, A. Effect of the Aegean Sea barrier between Europe and Asia on differentiation in Juniperus drupacea (Cupressaceae). Bot. J. Linn. Soc. 2016, 180, 365–385. [Google Scholar] [CrossRef] [Green Version]
- Poljak, I.; Idžojtić, M.; Šapić, I.; Korijan, P.; Vukelić, J. Diversity and structure of Croatian continental and Alpine-Dinaric populations of grey alder (Alnus incana /L./ Moench subsp. incana); Isolation by distance and environment explains phenotypic divergence. Šumar. List 2018, 142, 19–31. [Google Scholar] [CrossRef]
- Koutecký, P. MorphoTools: A set of R functions for morphometric analysis. Plant Syst. Evol. 2015, 301, 1115–1121. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. Available online: http://www.R-project.org/ (accessed on 25 October 2021).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Smouse, P.E.; Long, J.C.; Sokal, R. Multiple regression and correlation extensions of the Mantel test of matrix correspondence. Syst. Zool. 1986, 35, 627–632. [Google Scholar] [CrossRef]
- Manly, B.F.J. Randomization, Bootstrap and Monte Carlo Methods in Biology, 3rd ed.; Chapman & Hall/CRC, Taylor & Francis Group: Boca Raton, FL, USA, 2007; p. 480. [Google Scholar]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Version 2.2; Applied Biostatistics Inc.: New York, NY, USA, 2005; p. 44. [Google Scholar]
- Dempsey, D.; O’Flaherty, C.; Hook, I. Taxus Species—Morphological and chemical variations in needles and seeds. In Proceedings of the 4th International Conifer Conference, Wye, UK, 30 September 2003; Mill, R.R., Ed.; ISHS: Leuven, Belgium, 2003. [Google Scholar]
- Zarek, M. Variability of morphological features of needles, shoots and seeds of Taxus baccata L. in nature reservations of Southern Poland. EJPAU 2007, 10, 18. [Google Scholar]
- Iszkuło, G.; Jasinska, A.K.; Giertych, M.J.; Boratynski, A. Do secondary sexual dimorphism and female intolerance to drought influence the sex ratio and extinction risk of Taxus baccata? Plant Ecol. 2009, 200, 229–240. [Google Scholar] [CrossRef]
- Schirone, B.; Caetano Ferreira, R.; Vessella, F.; Schirone, A.; Piredda, R.; Cosimosi Simeone, M. Taxus baccata in the Azores: A relict form at risk of imminent exticinction. Biodivers. Conserv. 2010, 19, 1547–1565. [Google Scholar] [CrossRef]
- Vessella, F.; Simeone, M.C.; Fernandes, F.M.; Schirone, A.; Pires Gomes, M.; Schirone, B. Morphological and molecular data from Madeira support the persistence of an ancient lineage of Taxus baccata L. in Macaronesia and call for immediate conservation actions. Caryologia 2013, 66, 162–177. [Google Scholar] [CrossRef]
- Hamidouche-Si, M.C.; Bouhamed, A.; Vessella, F.; Krouchi, F.; Derridj, A. Geographic distribution and morphological variation of Taxus baccata in Algeria. Der Eibenfreund 2014, 20, 39–53. [Google Scholar]
- Stefanović, M. Varijabilnost terpenskog sastava i morfoloških karakteristika četina Taxus baccata L. u Srbiji. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 2015. [Google Scholar]
- Stefanović, M.; Nikolić, B.; Matić, R.; Popović, Z.; Vidaković, V.; Bojović, S. Exploration of sexual dimorphism of Taxus baccata L. needles in natural populations. Trees 2017, 31, 1697–1710. [Google Scholar] [CrossRef]
- Iszkuło, G.; Boratynsky, A. Analysis of the relationship between photosynthetic photon flux density and natural Taxus baccata seedlings occurrence. Acta Oecol. 2006, 29, 78–84. [Google Scholar] [CrossRef]
- Iszkuło, G.; Lewandowski, A.; Jasinska, A.K.; Dering, M. Light limitation of growth in 10-year-old seedling of Taxus baccata L. (European yew). Pol. J. Ecol. 2007, 55, 827–831. [Google Scholar]
- Robakowski, P.; Wyka, T.; Samardakiewicz, S.; Kierzkowski, D. Growth, photosynthesis, and needle structure of silver fir (Abies alba Mill.) seedlings under different canopies. For. Ecol. Manag. 2004, 201, 211–227. [Google Scholar] [CrossRef]
- Cescatti, A.; Zorer, R. Structural acclimation and radiation regime of silver fir (Abies alba Mill.) shoots along a light gradient. Plant Cell Environ. 2003, 26, 429–442. [Google Scholar] [CrossRef]
- Grassi, G.; Bagnaresi, U. Foliar morphological and physiological plasticity in Picea abies and Abies alba saplings along a natural light gradient. Tree Physiol. 2000, 21, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü. Acclimation to low irradiance in Picea abies: Influences of past and present light climate on foliage structure and function. Tree Physiol. 1996, 17, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Petritan, A.M.; von Lupke, B.; Petritan, I.C. Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur. J. For. Res. 2009, 128, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Adamidis, G.C.; Varsamis, G.; Tsiripidis, I.; Dimitrakopoulos, P.G.; Papageorgoiu, A.C. Patterns of leaf morphological traits of beech (Fagus sylvatica L.) along an altitudinal gradient. Forests 2021, 12, 14. [Google Scholar] [CrossRef]
- Boutsios, S.; Vidalis, A.; Adamidis, G.C.; Hatziskakis, S.; Varsamis, G.; Tsiripidis, I.; Karanikola, P.; Papageorgious, A.C. Diversity in shade and light leaf morphology in beech populations of South Rodopi mountains. In Proceedings of the National Academy of Sciences, India Section B: Biological Sciences; Ghosh, A., Ed.; Springer: Berlin, Germany, 2021; Volume 91, pp. 53–61. [Google Scholar] [CrossRef]
- Mitchell, A.K. Acclimation of Pacific yew (Taxus brevifolia) foliage to sun and shade. Tree Physiol. 1997, 18, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Petit, R.J.; Aguinagalde, I.; de Beaulieu, J.-L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial refugia: Hotspots but not melting pots of genetic diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar] [CrossRef]
- van Strien, M.J.; Holderegger, R.; Van Heck, H.J. Isolation-by-distance in landscapes: Considerations for landscape genetics. Heredity 2015, 11, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Mazur, M.; Boratynska, K.; Marcysiak, K.; Didukh, Y.; Romo, A.; Kosinski, P.; Boratynski, A. Low level of inter-populational differentiation in Juniperus excelsa M. Bieb. (Cupressaceae). Dendrobiology 2004, 52, 39–46. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Trait | Descriptive Parameters | Population | Region | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P01 | P02 | P03 | P04 | P05 | P06 | P07 | Dinaric | Continental | |||
| NA (mm2) | M | 0.29 | 0.32 | 0.39 | 0.46 | 0.41 | 0.45 | 0.51 | 0.33 | 0.46 | 0.40 |
| CV | 25.14 | 31.44 | 30.52 | 24.84 | 28.62 | 22.84 | 21.65 | 32.62 | 25.52 | 31.95 | |
| NL (mm) | M | 15.62 | 17.79 | 20.49 | 23.41 | 22.77 | 24.39 | 24.16 | 17.97 | 23.68 | 21.23 |
| CV | 18.16 | 22.56 | 21.89 | 15.45 | 21.32 | 17.32 | 18.29 | 24.15 | 18.35 | 24.41 | |
| MNW (mm) | M | 2.16 | 2.09 | 2.25 | 2.61 | 2.23 | 2.29 | 2.66 | 2.16 | 2.45 | 2.33 |
| CV | 10.27 | 13.69 | 12.03 | 22.22 | 17.93 | 16.72 | 17.13 | 12.42 | 20.41 | 18.90 | |
| PMNW (mm) | M | 5.45 | 6.06 | 6.10 | 8.41 | 7.06 | 7.60 | 7.26 | 5.87 | 7.58 | 6.85 |
| CV | 30.99 | 38.32 | 43.51 | 32.31 | 40.39 | 40.05 | 40.75 | 38.76 | 38.75 | 40.96 | |
| NW50 (mm) | M | 2.06 | 1.96 | 2.09 | 2.44 | 2.05 | 2.09 | 2.45 | 2.04 | 2.26 | 2.16 |
| CV | 10.48 | 13.50 | 12.41 | 22.73 | 17.73 | 16.78 | 18.11 | 12.44 | 21.01 | 18.93 | |
| NW90 (mm) | M | 1.30 | 1.30 | 1.41 | 1.57 | 1.39 | 1.37 | 1.62 | 1.33 | 1.49 | 1.42 |
| CV | 15.84 | 17.42 | 12.38 | 17.09 | 13.88 | 12.20 | 10.58 | 15.70 | 15.56 | 16.54 | |
| NA10 (°) | M | 25.76 | 24.00 | 23.53 | 19.72 | 19.81 | 19.12 | 21.94 | 24.43 | 20.15 | 21.98 |
| CV | 13.65 | 15.40 | 15.51 | 12.06 | 14.32 | 15.06 | 11.85 | 15.33 | 14.32 | 17.77 | |
| NA25 (°) | M | 13.84 | 11.96 | 11.30 | 10.72 | 9.95 | 9.35 | 11.02 | 12.37 | 10.26 | 11.16 |
| CV | 17.14 | 19.86 | 19.69 | 20.99 | 22.54 | 20.20 | 16.80 | 20.70 | 21.12 | 22.98 | |
| NL/MNW | M | 7.26 | 8.53 | 9.13 | 9.23 | 10.41 | 10.86 | 9.18 | 8.31 | 9.92 | 9.23 |
| CV | 17.18 | 17.68 | 18.67 | 20.21 | 22.77 | 20.61 | 17.17 | 20.32 | 21.81 | 23.07 | |
| Trait | Variance Component | df | % Variation | F | p |
|---|---|---|---|---|---|
| NA | Between regions | 1 | 34.81 | 12.43 | <0.05 |
| Among populations within regions | 5 | 8.23 | 4.72 | <0.01 | |
| Within populations | 98 | 32.35 | 40.44 | <0.01 | |
| Error | 24.61 | ||||
| NL | Between regions | 1 | 44.22 | 20.74 | <0.01 |
| Among populations within regions | 5 | 5.69 | 3.85 | <0.01 | |
| Within populations | 98 | 29.23 | 43.04 | <0.01 | |
| Error | 20.86 | ||||
| MNW | Between regions | 1 | 14.46 | 4.28 | 0.09 |
| Among populations within regions | 5 | 13.43 | 8.91 | <0.01 | |
| Within populations | 98 | 23.85 | 15.82 | <0.01 | |
| Error | 48.26 | ||||
| PMNW | Between regions | 1 | 16.11 | 18.97 | <0.01 |
| Among populations within regions | 5 | 2.14 | 3.31 | <0.01 | |
| Within populations | 98 | 11.60 | 5.96 | <0.01 | |
| Error | 70.15 | ||||
| NW50 | Between regions | 1 | 8.43 | 2.73 | 0.16 |
| Among populations within regions | 5 | 15.00 | 9.65 | <0.01 | |
| Within populations | 98 | 24.26 | 14.92 | <0.01 | |
| Error | 52.31 | ||||
| NW90 | Between regions | 1 | 14.32 | 3.69 | 0.11 |
| Among populations within regions | 5 | 16.31 | 9.36 | <0.01 | |
| Within populations | 98 | 27.90 | 21.18 | <0.01 | |
| Error | 41.47 | ||||
| NA10 | Between regions | 1 | 43.88 | 21.33 | <0.01 |
| Among populations within regions | 5 | 5.41 | 3.72 | <0.01 | |
| Within populations | 98 | 29.09 | 41.35 | <0.01 | |
| Error | 21.62 | ||||
| NA25 | Between regions | 1 | 24.94 | 7.36 | <0.05 |
| Among populations within regions | 5 | 11.36 | 6.42 | <0.01 | |
| Within populations | 98 | 30.30 | 28.22 | <0.01 | |
| Error | 33.40 | ||||
| NL/MNW | Between regions | 1 | 20.60 | 7.11 | <0.05 |
| Among populations within regions | 5 | 12.85 | 6.24 | <0.01 | |
| Within populations | 98 | 35.78 | 35.89 | <0.01 | |
| Error | 30.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tumpa, K.; Liber, Z.; Šatović, Z.; Medak, J.; Idžojtić, M.; Vidaković, A.; Vukelić, J.; Šapić, I.; Nikl, P.; Poljak, I. High Level of Phenotypic Differentiation of Common Yew (Taxus baccata L.) Populations in the North-Western Part of the Balkan Peninsula. Forests 2022, 13, 78. https://doi.org/10.3390/f13010078
Tumpa K, Liber Z, Šatović Z, Medak J, Idžojtić M, Vidaković A, Vukelić J, Šapić I, Nikl P, Poljak I. High Level of Phenotypic Differentiation of Common Yew (Taxus baccata L.) Populations in the North-Western Part of the Balkan Peninsula. Forests. 2022; 13(1):78. https://doi.org/10.3390/f13010078
Chicago/Turabian StyleTumpa, Katarina, Zlatko Liber, Zlatko Šatović, Jasnica Medak, Marilena Idžojtić, Antonio Vidaković, Joso Vukelić, Irena Šapić, Patricija Nikl, and Igor Poljak. 2022. "High Level of Phenotypic Differentiation of Common Yew (Taxus baccata L.) Populations in the North-Western Part of the Balkan Peninsula" Forests 13, no. 1: 78. https://doi.org/10.3390/f13010078
APA StyleTumpa, K., Liber, Z., Šatović, Z., Medak, J., Idžojtić, M., Vidaković, A., Vukelić, J., Šapić, I., Nikl, P., & Poljak, I. (2022). High Level of Phenotypic Differentiation of Common Yew (Taxus baccata L.) Populations in the North-Western Part of the Balkan Peninsula. Forests, 13(1), 78. https://doi.org/10.3390/f13010078