
Stem Growth of Horse Chestnut (Aesculus hippocastanum L.) under a Warming Climate—Tree Age Matters



Abstract
:1. Introduction
2. Methods
2.1. Study Site and Experimental Design
2.2. Meteorological and Soil Water Variables
2.3. Stem Radius Variation
2.4. Growth Reaction Assessment
2.5. Statistical Analysis
3. Results
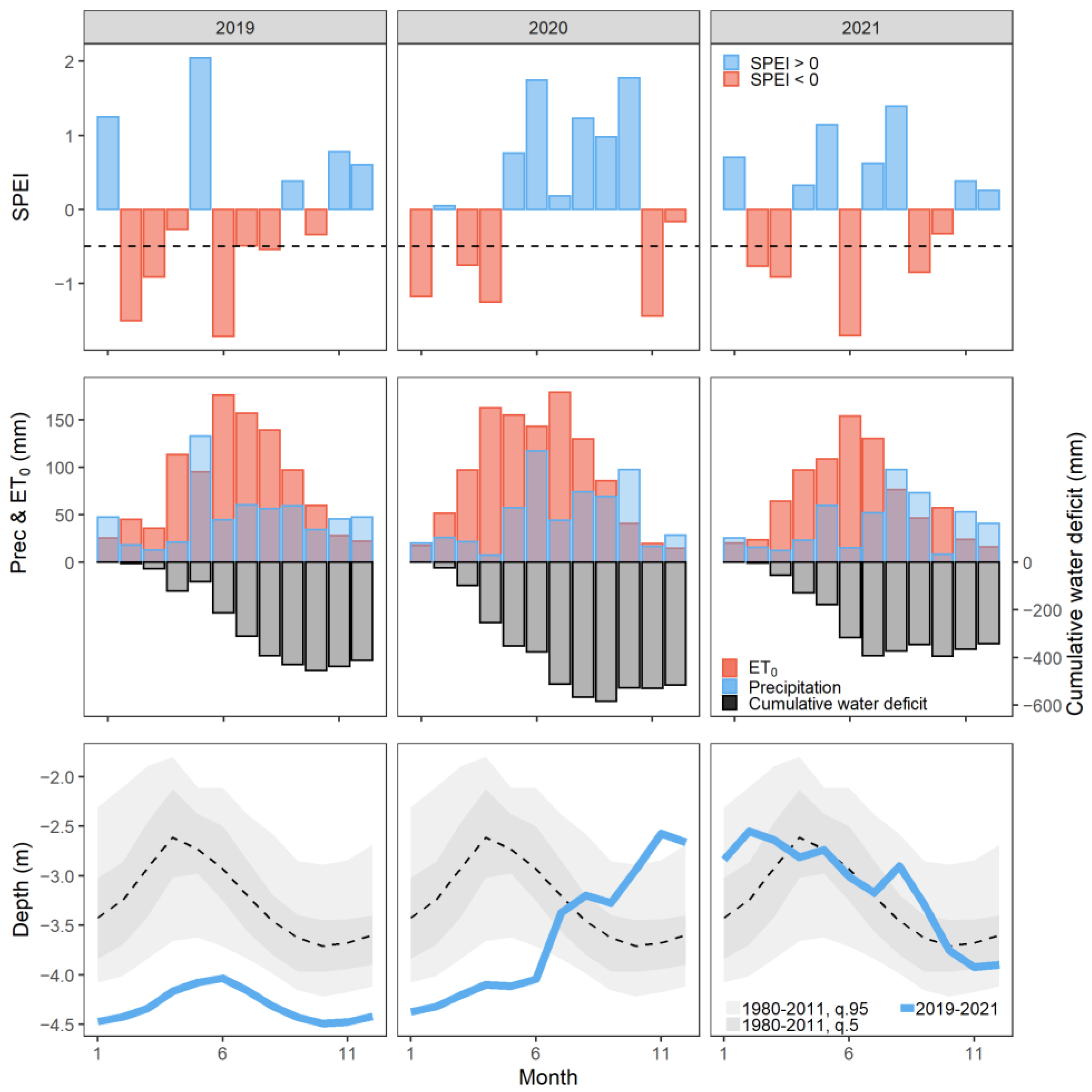
3.1. Meteorological Conditions
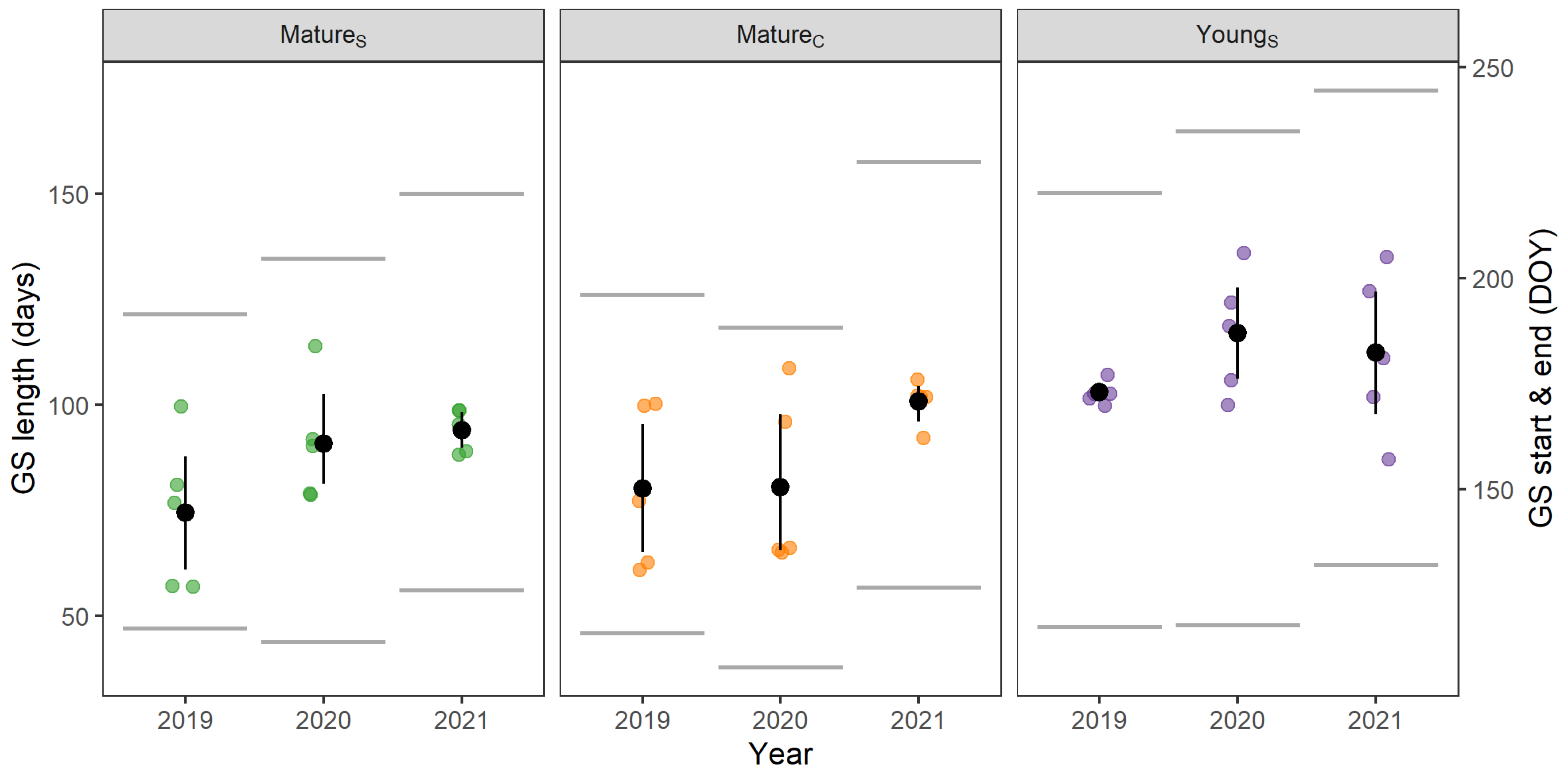
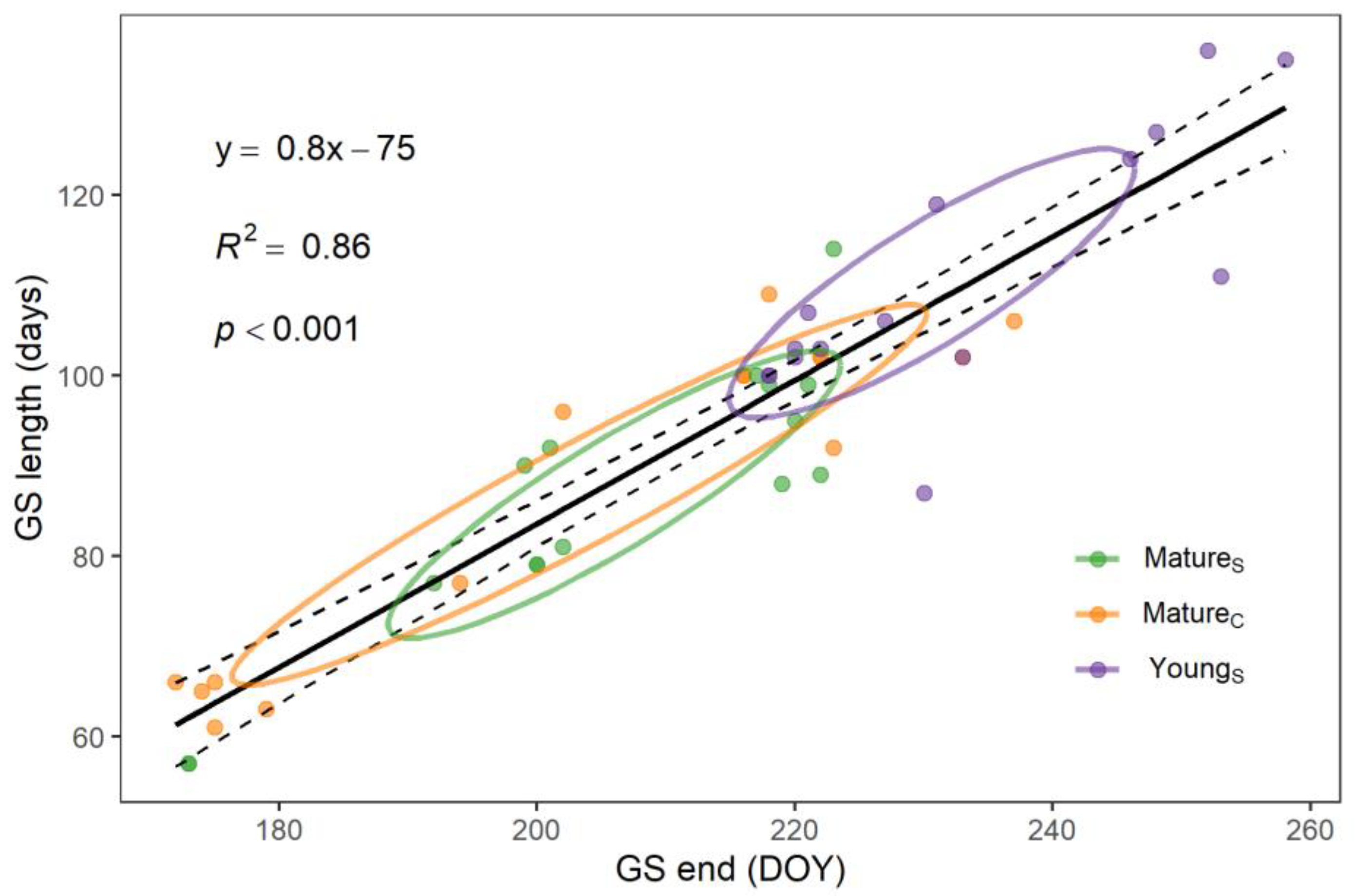
3.2. Phenology
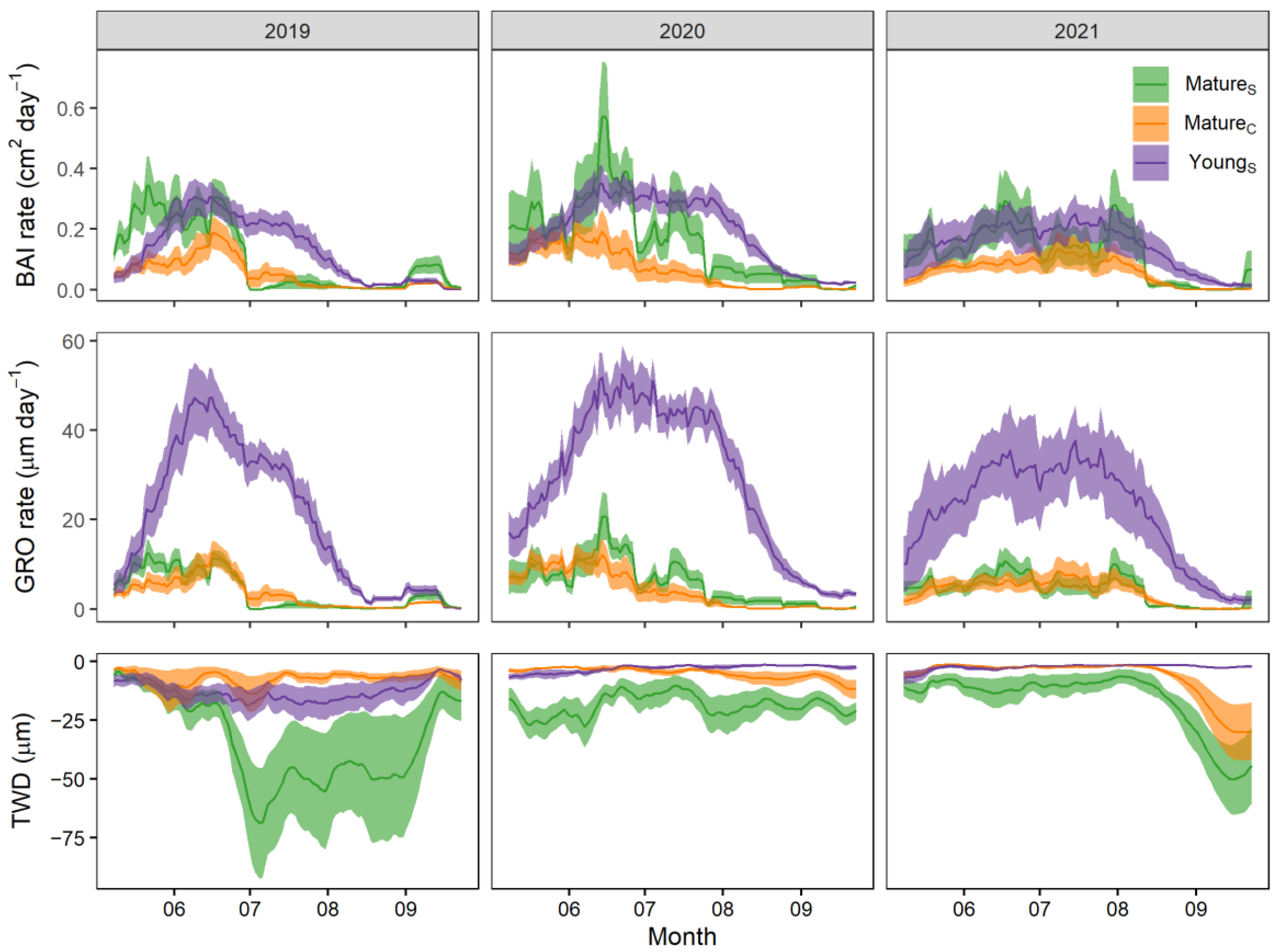
3.3. Secondary Growth Dynamics
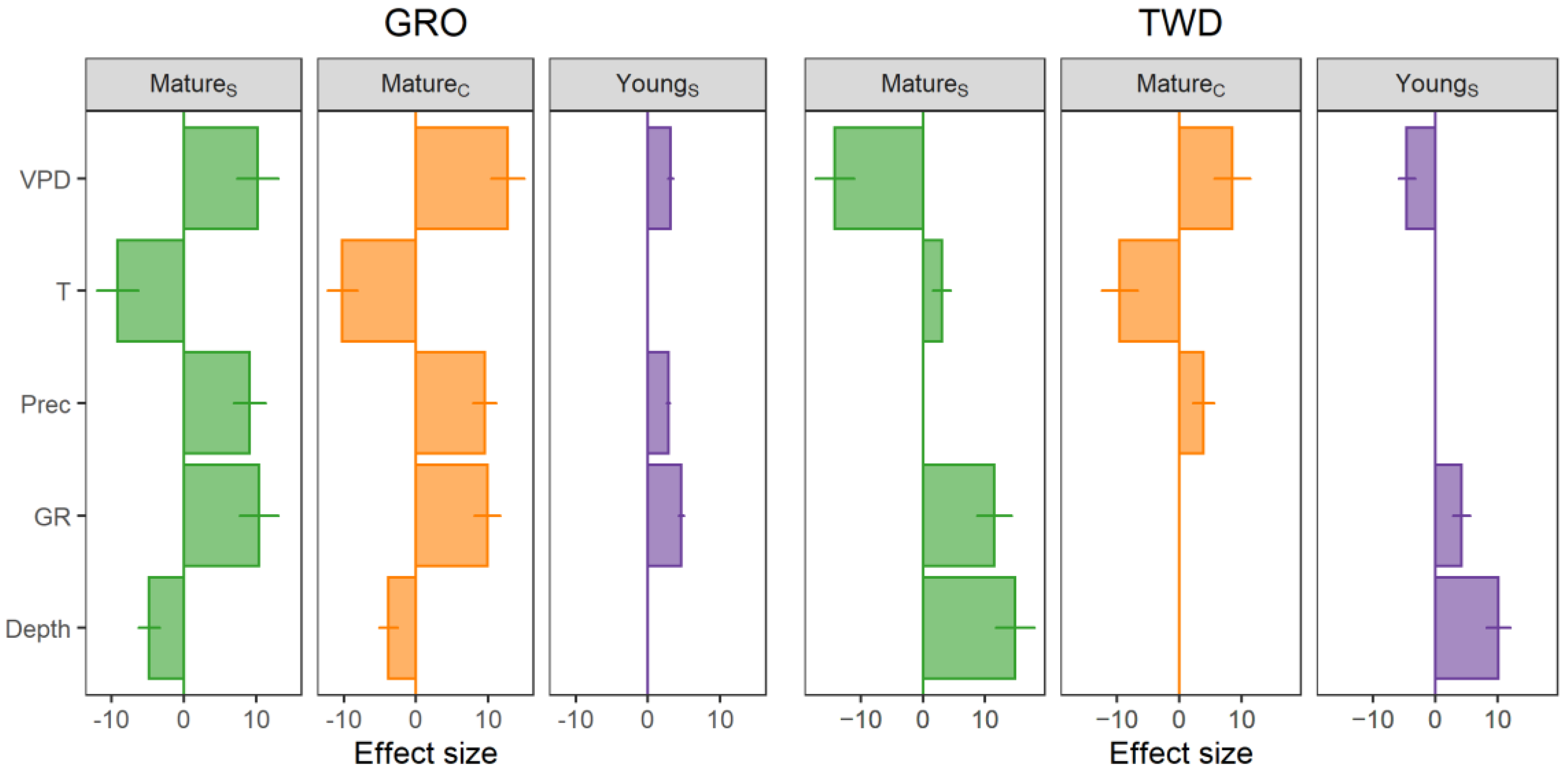
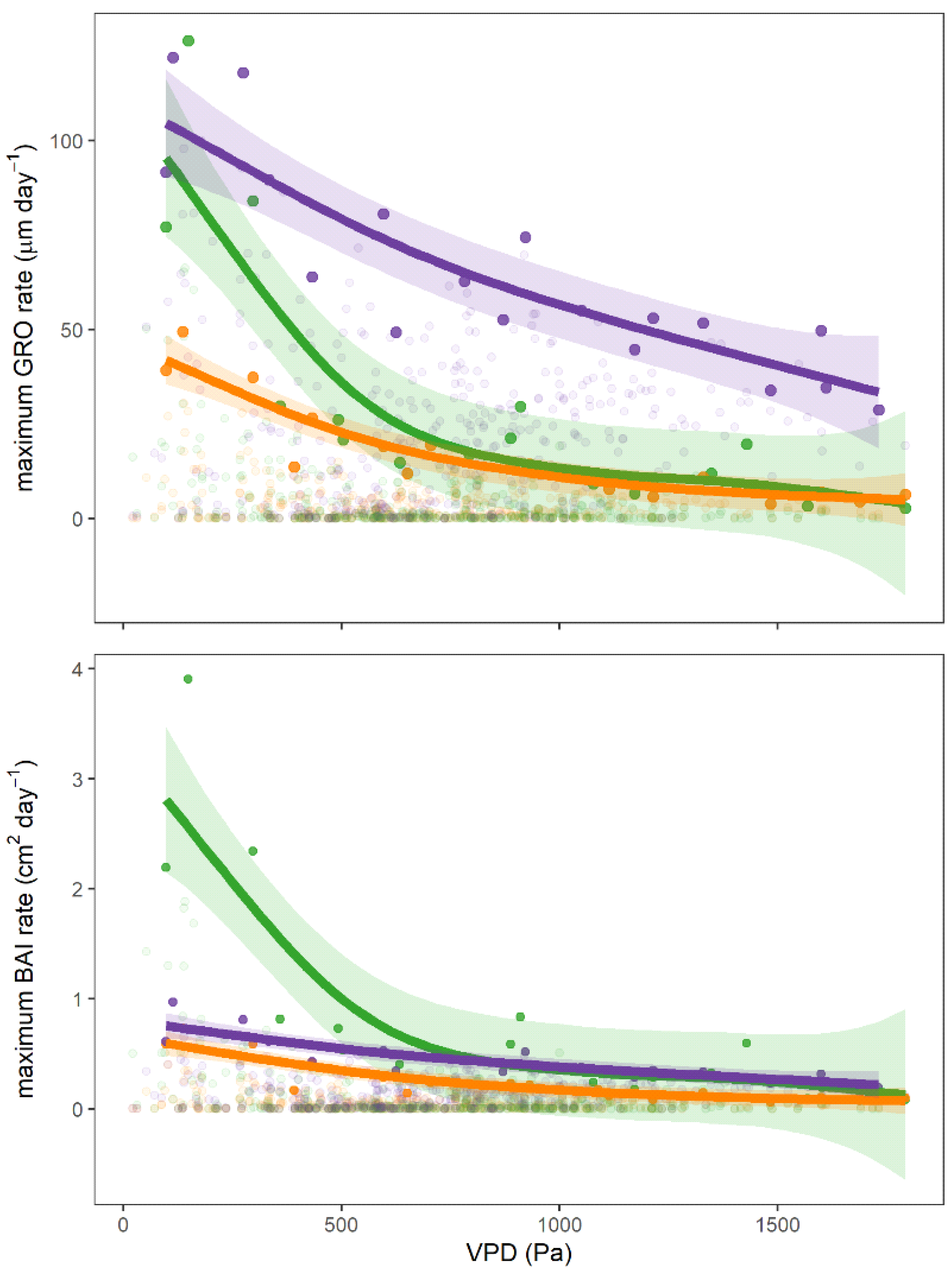
3.4. Environmental Factors and Tree Growth
4. Discussion
4.1. Meteorological Conditions
4.2. Phenology
4.3. Secondary Growth and Water Stress
4.4. Environmental Factors and Secondary Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lack, H.W. The Discovery and Rediscovery of the Horse Chestnut. Arnoldia 2002, 61, 15–19. [Google Scholar] [CrossRef]
- Mcbride, J.R. The World’s Urban Forests; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 9783319521077. [Google Scholar]
- Profous, G.V.; Rowntree, R.A. The Structure And Management of the Urban Forest in Prague, Czechoslovakia. II History of Prague Greenspace Management. Arboric. J. 1993, 17, 117–130. [Google Scholar] [CrossRef]
- Braude, R. Value of Horse Chestnuts as Food for Pigs. Nature 1943, 152, 571–572. [Google Scholar] [CrossRef]
- Klimo, E.; Kulhavý, J.; Prax, A.; Menšík, L.; Hadaš, P.; Mauer, O. Functioning of South Moravian Floodplain Forests (Czech Republic) in Forest Environment Subject to Natural and Anthropogenic Change. Int. J. For. Res. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Šenfeldr, M.; Horák, P.; Kvasnica, J.; Šrámek, M.; Hornová, H.; Maděra, P. Species-Specific Effects of Groundwater Level Alteration on Climate Sensitivity of Floodplain Trees. Forests 2021, 12, 1178. [Google Scholar] [CrossRef]
- Paterska, M.; Bandurska, H.; Wysłouch, J.; Molińska-Glura, M.; Moliński, K. Chemical composition of horse-chestnut (Aesculus) leaves and their susceptibility to chestnut leaf miner Cameraria ohridella Deschka & Dimić. Acta Physiol. Plant. 2017, 39, 105. [Google Scholar] [CrossRef] [Green Version]
- Straw, N.A.; Bellett-Travers, M. Impact and management of the horse chestnut leaf-miner (Cameraria ohridella). Arboric. J. 2004, 28, 67–83. [Google Scholar] [CrossRef]
- Percival, G.C.; Barrow, I.; Noviss, K.; Keary, I.; Pennington, P. The impact of horse chestnut leaf miner (Cameraria ohridella Deschka and Dimic; HCLM) on vitality, growth and reproduction of Aesculus hippocastanum L. Urban For. Urban Green. 2011, 10, 11–17. [Google Scholar] [CrossRef]
- Nardini, A.; Raimondo, F.; Scimone, M.; Salleo, S. Impact of the leaf miner Cameraria ohridella on whole-plant photosynthetic productivity of Aesculus hippocastanum: Insights from a model. Trees 2004, 18, 714–721. [Google Scholar] [CrossRef]
- Granier, A.; Reichstein, M.; Bréda, N.; Janssens, I.; Falge, E.; Ciais, P.; Grünwald, T.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; et al. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric. For. Meteorol. 2007, 143, 123–145. [Google Scholar] [CrossRef]
- Ruehr, N.K.; Grote, R.; Mayr, S.; Arneth, A. Beyond the extreme: Recovery of carbon and water relations in woody plants following heat and drought stress. Tree Physiol. 2019, 39, 1285–1299. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.B.; Castro, V.; Fickle, J.C.; Jacobsen, A.L. Embolism resistance of different aged stems of a California oak species (Quercus douglasii): Optical and microCT methods differ from the benchtop-dehydration standard. Tree Physiol. 2019, 40, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Brutovská, E.; Sámelová, A.; Dušička, J.; Mičieta, K. Ageing of trees: Application of general ageing theories. Ageing Res. Rev. 2013, 12, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J. Plant Physiology: A big issue for trees. Nature 2005, 437, 965–966. [Google Scholar] [CrossRef] [PubMed]
- Czech Hydrometeorological Institute Public Data. Available online: https://www.chmi.cz/ (accessed on 1 July 2022).
- Gatsuk, L.E.; Smirnova, O.V.; Vorontzova, L.I.; Zhukova, L.B.Z.A. Age States of Plants of Various Growth Forms: A Review. J. Ecol. 1980, 68, 675. [Google Scholar] [CrossRef]
- Penman, H.L. Evaporation: An Introductory Survey. Neth. J. Agric. Sci. 1956, 4, 9–29. [Google Scholar] [CrossRef]
- Penman, H.L. Natural Evaporation from Open Water, Bare Soil and Grass. Crop Sci. 1948, 193, 120–145. [Google Scholar]
- Beguería, S.; Latorre, L.; Reig, F.; Vicente-Serrano, S.M. Global SPEI Database. Available online: https://spei.csic.es/ (accessed on 30 August 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Beguería, S.; Vicente-Serrano, M.S.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Lee, S.-H.; Yoo, S.-H.; Kim, T. Analysis of Drought Intensity and Trends Using the Modified SPEI in South Korea from 1981 to 2010. Water 2018, 10, 327. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Zweifel, R. Radial stem variations—A source of tree physiological information not fully exploited yet. Plant Cell Environ. 2015, 39, 231–232. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, L.; Zweifel, R.; Kahmen, A. Daily stem diameter variations can predict the canopy water status of mature temperate trees. Tree Physiol. 2018, 38, 941–952. [Google Scholar] [CrossRef]
- Kyselý, J. Trends in heavy precipitation in the Czech Republic over 1961-2005. Int. J. Clim. 2008, 29, 1745–1758. [Google Scholar] [CrossRef]
- Brázdil, R.; Zahradníček, P.; Dobrovolný, P.; Štěpánek, P.; Trnka, M. Observed changes in precipitation during recent warming: The Czech Republic, 1961–2019. Int. J. Clim. 2021, 41, 3881–3902. [Google Scholar] [CrossRef]
- Zahradníček, P.; Brázdil, R.; Štěpánek, P.; Trnka, M. Reflections of global warming in trends of temperature characteristics in the Czech Republic, 1961–2019. Int. J. Clim. 2020, 41, 1211–1229. [Google Scholar] [CrossRef]
- Seiwa, K. Changes in Leaf Phenology Are Dependent on Tree Height in Acer Mono, a Deciduous Broad-Leaved Tree. Ann. Bot. 1999, 83, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Augspurger, C.K.; Bartlett, E.A. Differences in Leaf Phenology between Juvenile and Adult Trees in a Temperate Deciduous Forest. Tree Physiol. 2003, 23, 517–525. [Google Scholar] [CrossRef]
- Mediavilla, S.; Escudero, A. Ontogenetic changes in leaf phenology of two co-occurring Mediterranean oaks differing in leaf life span. Ecol. Res. 2009, 24, 1083–1090. [Google Scholar] [CrossRef]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B.K. Temperate and boreal forest tree phenology: From organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H.; Wünsche, J.N.; Norling, C.L.; Wiggins, H.N. Root-Zone Temperatures Affect Phenology of Bud Break, Flower Clus-terdevelopment, Shoot Extension Growth and Gas Exchange of ‘Braeburn’ (Malus Domestica) Apple Trees. Tree Physiol. 2005, 105–111. [Google Scholar]
- De Barba, D.; Rossi, S.; DesLauriers, A.; Morin, H. Effects of soil warming and nitrogen foliar applications on bud burst of black spruce. Trees 2016, 30, 87–97. [Google Scholar] [CrossRef]
- Suzuki, M.; Yoda, K.; Suzuki, H. Phenological Comparison of the Onset of Vessel Formation between Ring-Porous and Diffuse-Porous Deciduous Trees in a Japanese Temperate Forest. IAWA J. 1996, 17, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Michelot, A.; Bréda, N.; Damesin, C.; Dufrêne, E. Differing growth responses to climatic variations and soil water deficits of Fagus sylvatica, Quercus petraea and Pinus sylvestris in a temperate forest. For. Ecol. Manag. 2012, 265, 161–171. [Google Scholar] [CrossRef]
- Urban, J.; Dvořák, M. Sap flow-based quantitative indication of progression of Dutch elm disease after inoculation with Ophiostoma novoulmi. Trees 2014, 28, 1599–1605. [Google Scholar] [CrossRef]
- Sanz-Pérez, V.; Castro-Diez, P. Summer water stress and shade alter bud size and budburst date in three mediterranean Quercus species. Trees 2010, 24, 89–97. [Google Scholar] [CrossRef]
- Spieß, N.; Oufir, M.; Matušíková, I.; Stierschneider, M.; Kopecky, D.; Homolka, A.; Burg, K.; Fluch, S.; Hausman, J.-F.; Wilhelm, E. Ecophysiological and transcriptomic responses of oak (Quercus robur) to long-term drought exposure and rewatering. Environ. Exp. Bot. 2012, 77, 117–126. [Google Scholar] [CrossRef]
- Kuster, T.M.; Dobbertin, M.; Günthardt-Goerg, M.S.; Schaub, M.; Arend, M. A Phenological Timetable of Oak Growth under Experimental Drought and Air Warming. PLoS ONE 2014, 9, e89724. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of Threshold Temperatures for Xylogenesis in Conifers at High Altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Gričar, J. Xylem and Phloem Formation in Sessile Oak from Slovenia in 2007. Wood Res. 2010, 55, 15–22. [Google Scholar]
- Giagli, K.; Gričar, J.; Vavrčík, H.; Menšík, L.; Gryc, V. The effects of drought on wood formation in fagus sylvatica during two contrasting years. IAWA J. 2016, 37, 332–348. [Google Scholar] [CrossRef]
- Defila, C.; Clot, B. Phytophenological trends in Switzerland. Int. J. Biometeorol. 2001, 45, 203–207. [Google Scholar] [CrossRef]
- Boergens, E.; Güntner, A.; Dobslaw, H.; Dahle, C. Quantifying the Central European Droughts in 2018 and 2019 with grace follow-on. Geophys. Res. Lett. 2020, 47, e2020GL087285. [Google Scholar] [CrossRef]
- Hari, V.; Rakovec, O.; Markonis, Y.; Hanel, M.; Kumar, R. Increased future occurrences of the exceptional 2018–2019 Central European drought under global warming. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Mariën, B.; Papadimitriou, D.; Kotilainen, T.; Zuccarini, P.; Dox, I.; Verlinden, M.; Heinecke, T.; Mariën, J.; Willems, P.; Decoster, M.; et al. Timing leaf senescence: A generalized additive models for location, scale and shape approach. Agric. For. Meteorol. 2022, 315, 108823. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Brienen, R.J.W.; Gloor, E.; Phillips, O.L.; Prior, L.D. Detecting trends in tree growth: Not so simple. Trends Plant Sci. 2013, 18, 11–17. [Google Scholar] [CrossRef]
- Bosshard, H.H. Aspects of the Aging Process in Cambium and Xylem. Holzforschung 1965, 19, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Mäkinen, H.; Seo, J.-W.; Nöjd, P.; Schmitt, U.; Jalkanen, R. Seasonal dynamics of wood formation: A comparison between pinning, microcoring and dendrometer measurements. Forstwiss. Centralblatt 2008, 127, 235–245. [Google Scholar] [CrossRef]
- Loehle, C. Biomechanical constraints on tree architecture. Trees 2016, 30, 2061–2070. [Google Scholar] [CrossRef]
- Lehnebach, R.; Beyer, R.; Letort, V.; Heuret, P. The pipe model theory half a century on: A review. Ann. Bot. 2018, 121, 773–795. [Google Scholar] [CrossRef]
- Bartelink, H.H. Allometric relationships for biomass and leaf area of beech (Fagus sylvatica L.). Ann. Sci. For. 1997, 54, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]
- Lilles, E.B.; Astrup, R. Multiple resource limitation and ontogeny combined: A growth rate comparison of three co-occurring conifers. Can. J. For. Res. 2012, 42, 99–110. [Google Scholar] [CrossRef]
- Jyske, T.; Hölttä, T. Comparison of phloem and xylem hydraulic architecture in Piceaabies stems. New Phytol. 2014, 205, 102–115. [Google Scholar] [CrossRef]
- Cardoso, S.; Quilhó, T.; Pereira, H. Influence of cambial age on the bark structure of Douglasfir. Wood Sci. Technol. 2018, 53, 191–210. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [Green Version]
- Merilo, E.; Yarmolinsky, D.; Jalakas, P.; Parik, H.; Tulva, I.; Rasulov, B.; Kilk, K.; Kollist, H. Stomatal VPD Response: There Is More to the Story Than ABA. Plant Physiol. 2017, 176, 851–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- Woodruff, D.R.; Meinzer, F.C. Water stress, shoot growth and storage of non-structural carbohydrates along a tree height gradient in a tall conifer. Plant Cell Environ. 2011, 34, 1920–1930. [Google Scholar] [CrossRef]
- Bačovský, V.; Vyhnánek, T.; Hanáček, P.; Mertelík, J.; Šafránková, I. Genetic diversity of chestnut tree in relation to susceptibility to leaf miner (Cameraria ohridella Deschka & Dimič). Trees 2017, 31, 753–763. [Google Scholar] [CrossRef]
- Klepper, E.L.; Gano, K.A.; Cadwell, L.L. Rooting Depth and Distributions of Deep-Rooted Plants in the 200 Area Control Zone of the Hanford Site; No. PNL-5247; Pacific Northwest Laboratory: Richland, WA, USA, 1985. [Google Scholar]
- Schenk, H.J.; Jackson, R.B. The Global Biogeography of Roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- Fan, Y.; Miguez-Macho, G.; Jobbágy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic regulation of plant rooting depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef] [Green Version]
- Kannenberg, S.A.; Schwalm, C.R.; Anderegg, W.R.L. Ghosts of the past: How drought legacy effects shape forest functioning and carbon cycling. Ecol. Lett. 2020, 23, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Salomón, R.L.; Peters, R.L.; Zweifel, R.; Sass-Klaassen, U.G.W.; Stegehuis, A.I.; Smiljanic, M.; Poyatos, R.; Babst, F.; Cienciala, E.; Fonti, P.; et al. The 2018 European heatwave led to stem dehydration but not to consistent growth reductions in forests. Nat. Commun. 2022, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; Lopez-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | T | Prec |
|---|---|---|
| (°C) | (mm) | |
| 2019 | 11.2 | 581 |
| 2020 | 10.3 | 580 |
| 2021 | 9.52 | 478 |
| 1981–2010 | 10.6 | 587 |
| MATUREC | MATURES | YOUNGS | Tree | Year | ||
|---|---|---|---|---|---|---|
| Estimate [95 CI] | % | % | ||||
| BAI | (cm2) | 5.9 [3.3–10.6] a | 13.9 [7.8–25] a,b | 17.2 [9.6–31] b | 73 | 21 |
| GRO | (μm) | 408.8 [252.2–662.7] a | 531.5 [158.2–1786] a | 2668 [794–8964] b | 67 | 26 |
| TWD | (μm) | 488.4 [296.3–804.9] a | 1914 [546.6–6705] b | 184.7 [52.7–647] c | <1 | 96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plichta, R.; Úradníček, L.; Gebauer, R. Stem Growth of Horse Chestnut (Aesculus hippocastanum L.) under a Warming Climate—Tree Age Matters. Forests 2022, 13, 1677. https://doi.org/10.3390/f13101677
Plichta R, Úradníček L, Gebauer R. Stem Growth of Horse Chestnut (Aesculus hippocastanum L.) under a Warming Climate—Tree Age Matters. Forests. 2022; 13(10):1677. https://doi.org/10.3390/f13101677
Chicago/Turabian StylePlichta, Roman, Luboš Úradníček, and Roman Gebauer. 2022. "Stem Growth of Horse Chestnut (Aesculus hippocastanum L.) under a Warming Climate—Tree Age Matters" Forests 13, no. 10: 1677. https://doi.org/10.3390/f13101677
APA StylePlichta, R., Úradníček, L., & Gebauer, R. (2022). Stem Growth of Horse Chestnut (Aesculus hippocastanum L.) under a Warming Climate—Tree Age Matters. Forests, 13(10), 1677. https://doi.org/10.3390/f13101677