



Distribution Pattern and Factors Influencing Spontaneous Plant Diversity in Different Wetland Habitats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
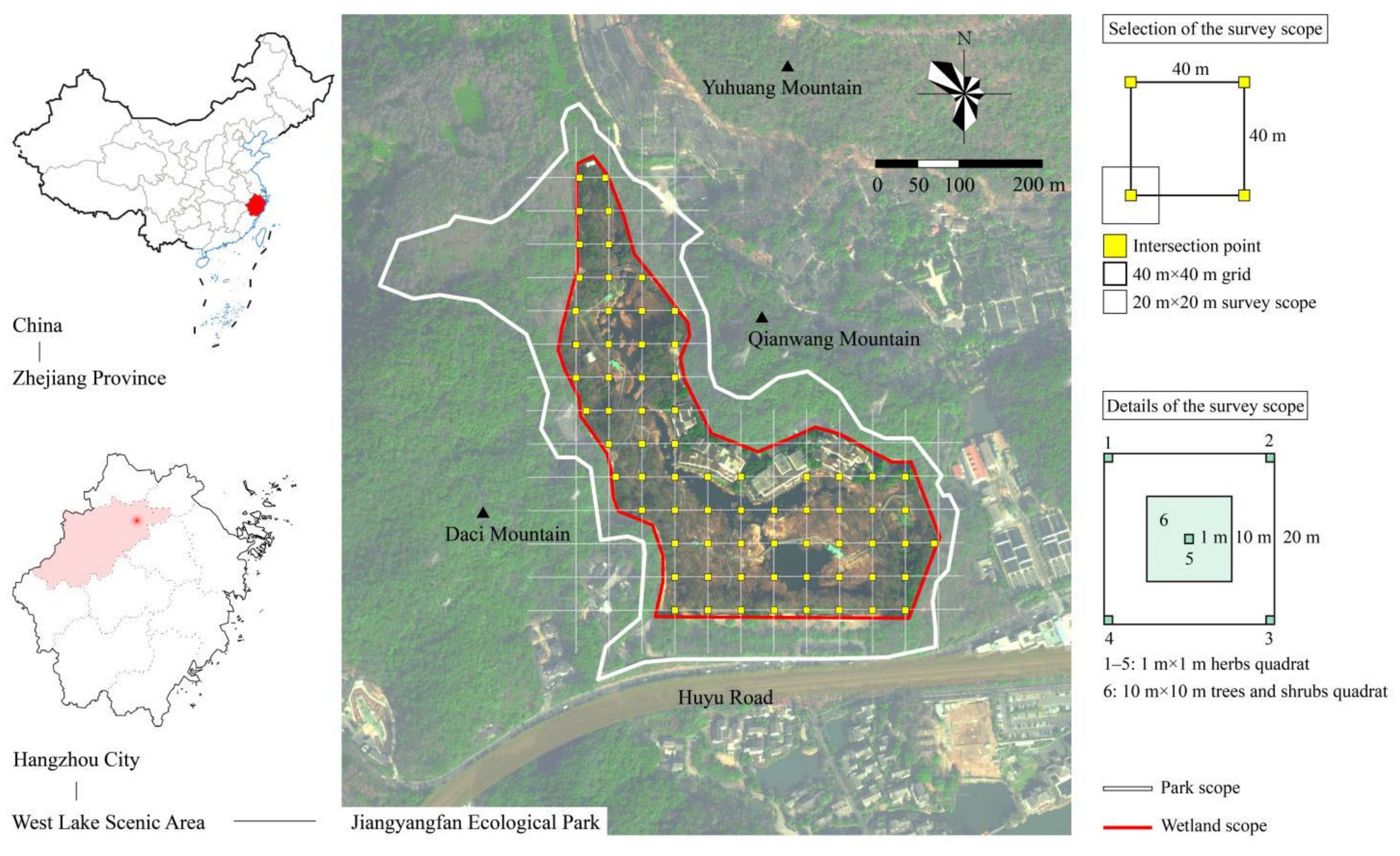
2.1. Study Area
2.2. Plant Data
2.3. Environmental Data
2.4. Statistical Analysis
3. Results
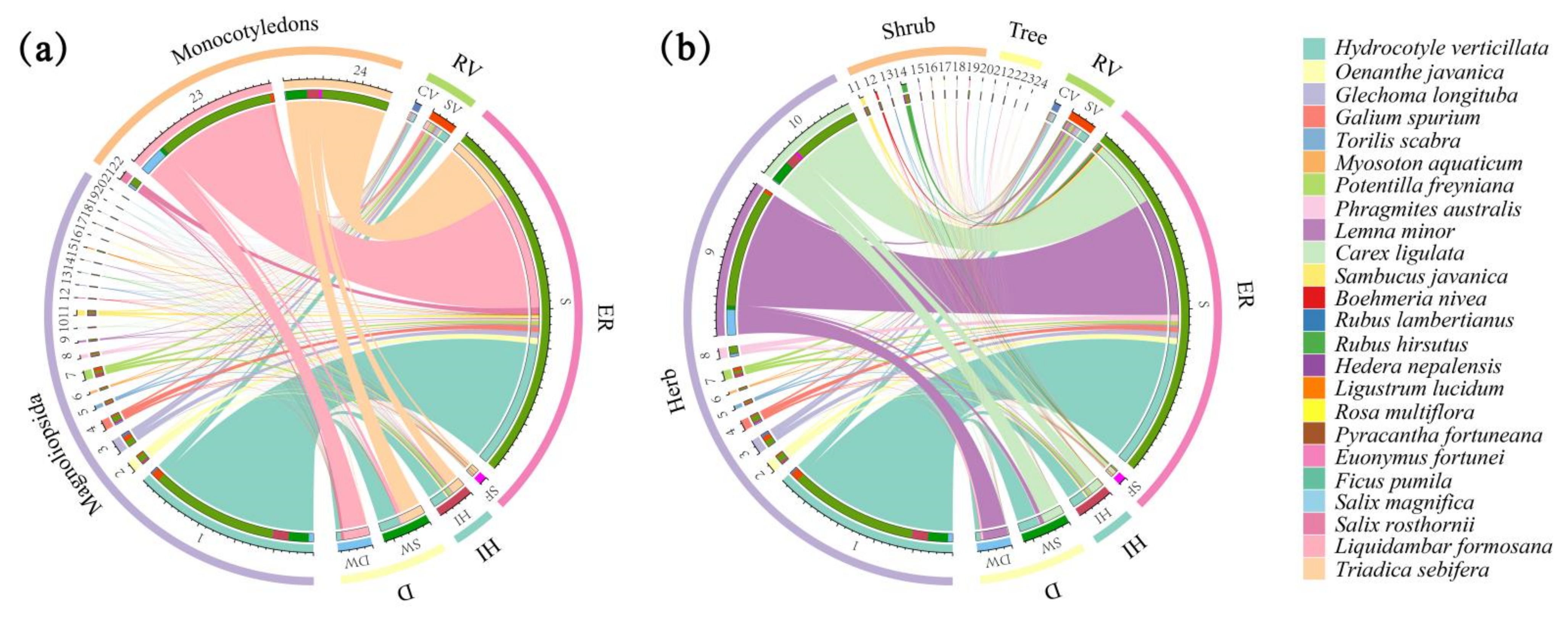
3.1. Composition of Spontaneous Plant Species
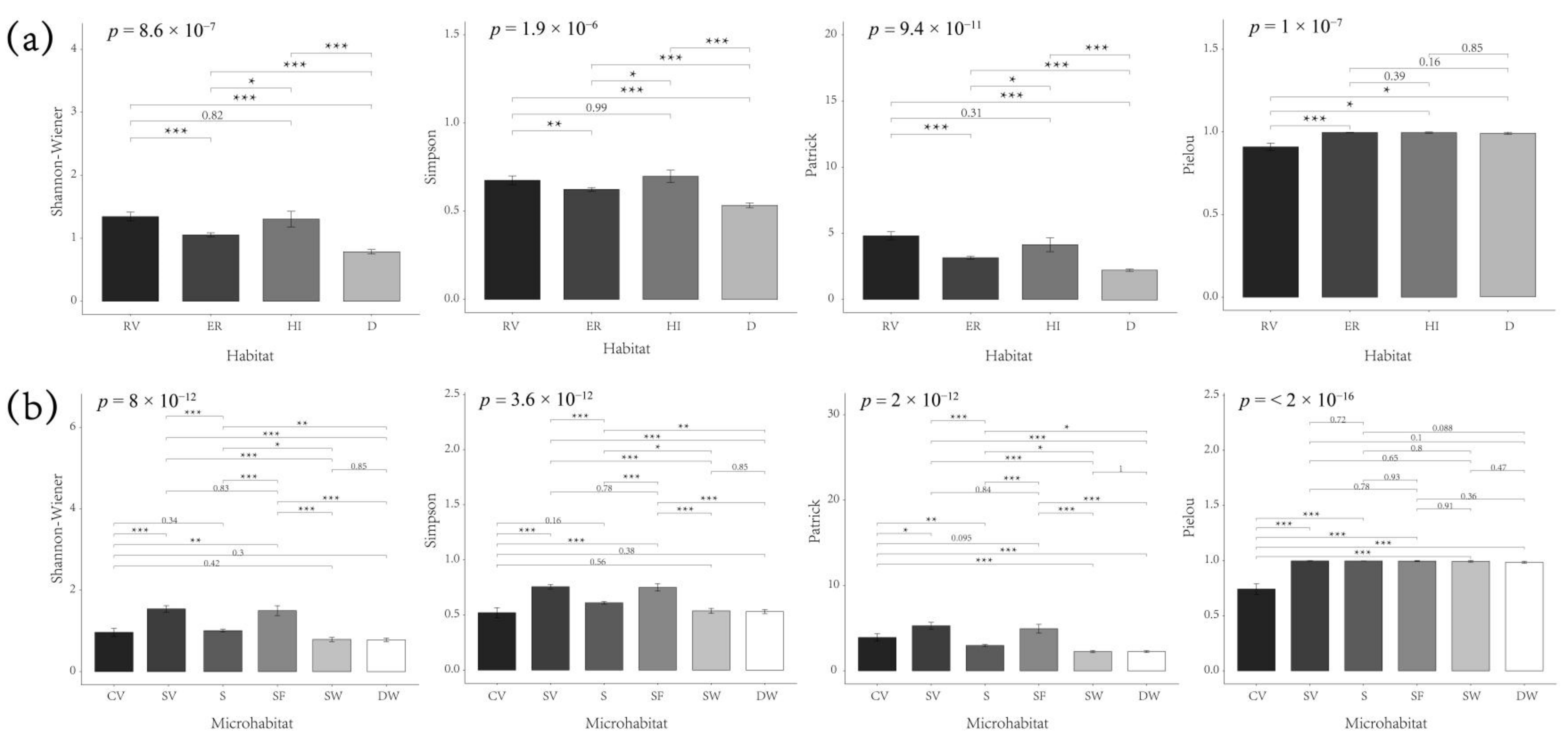
3.2. Diversity and Distribution Pattern of Spontaneous Plants in Different Habitats
3.3. Fruit Types and Seed Dispersal of Spontaneous Plants at Different Habitat Scales
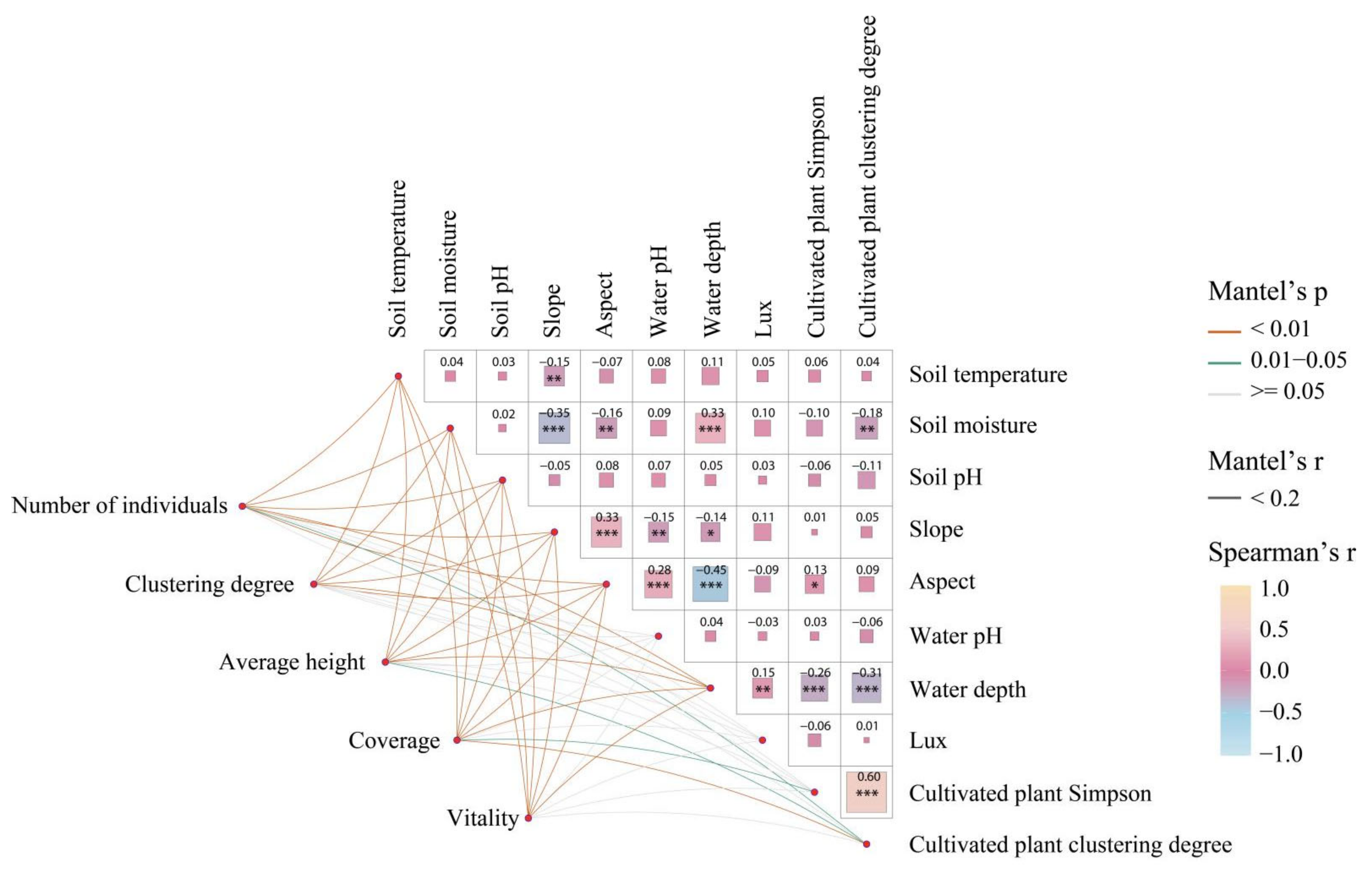
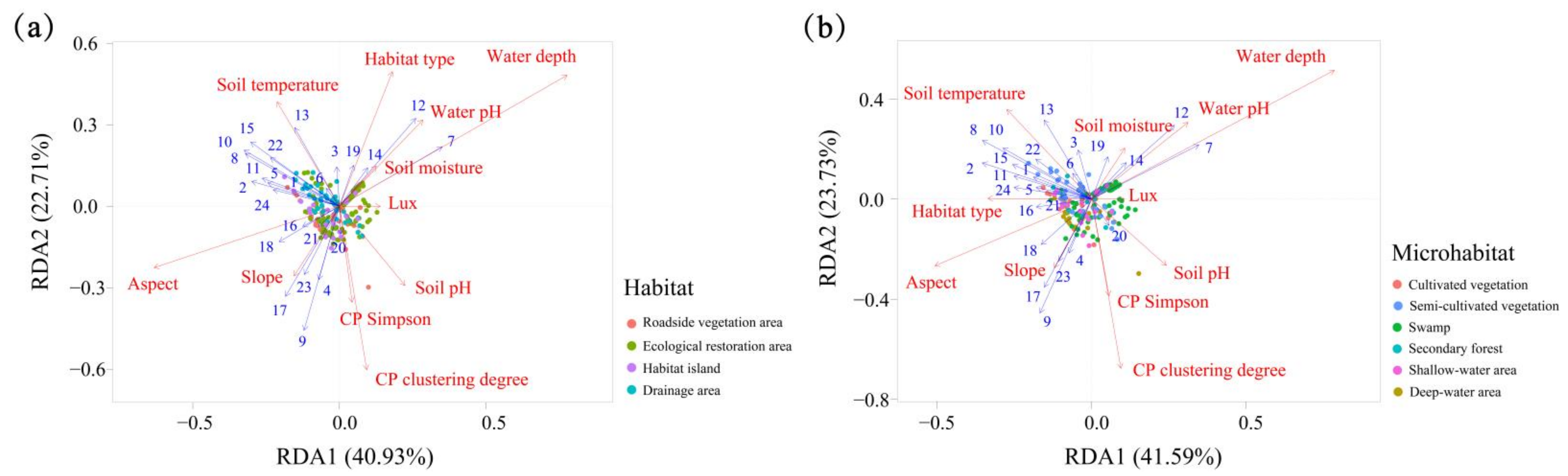
3.4. Effects of Different Environmental Factors on the Distribution of Spontaneous Plants
4. Discussion
4.1. Habitat Heterogeneity of Spontaneous Plants
4.2. Effects of Biological Characteristics of Spontaneous Plants on Distribution
4.3. Influence of Environmental Factors on the Distribution of Spontaneous Plants
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, A.L.; Borowy, D.; Swan, C.M. Land use history and seed dispersal drive divergent plant community assembly patterns in urban vacant lots. J. Appl. Ecol. 2018, 55, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wang, Y.; Li, G.; Fang, X. Vegetation response to urban landscape spatial pattern change in the Yangtze River Delta, China. Sustainability 2020, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Estevo, C.A.; Nagy-Reis, M.B.; Silva, W.R. Urban parks can maintain minimal resilience for Neotropical bird communities. Urban For. Urban Green. 2017, 27, 84–89. [Google Scholar] [CrossRef]
- Duncan, J.M.A.; Boruff, B.; Saunders, A.; Sun, Q.; Hurley, J.; Amati, M. Turning down the heat: An enhanced understanding of the relationship between urban vegetation and surface temperature at the city scale. Sci. Total Environ. 2019, 656, 118–128. [Google Scholar] [CrossRef]
- Ren, Z.; Zheng, H.; He, X.; Zhang, D.; Yu, X. Estimation of the relationship between urban vegetation configuration and land surface temperature with remote sensing. J. Indian Soc. Remote Sens. 2015, 43, 89–100. [Google Scholar] [CrossRef]
- Li, L.; Zhou, X.; Chen, L.; Chen, L.; Zhang, Y.; Liu, Y. Estimating urban vegetation biomass from sentinel-2A image data. Forests 2020, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Liu, J. Quantifying urban vegetation coverage change with a linear spectral mixing model: A case study in Xi’an, China. Ecol. Chem. Eng. S 2021, 28, 87–100. [Google Scholar] [CrossRef]
- Wei, J.; Li, H.; Wang, Y.; Xu, X. The cooling and humidifying effects and the thresholds of plant community structure parameters in urban aggregated green infrastructure. Forests 2021, 12, 111. [Google Scholar] [CrossRef]
- Kendal, D.; Williams, K.J.H.; Williams, N.S.G. Plant traits link people’s plant preferences to the composition of their gardens. Landscape Urban Plan. 2012, 105, 34–42. [Google Scholar] [CrossRef]
- Samimi, P.M.; Shahhosseini, H. Evaluation of resident’s indoor green space preferences in residential complexes based on plants’ characteristics. Indoor Built Environ. 2020, 30, 859–868. [Google Scholar] [CrossRef]
- Xu, W.; Svenning, J.C.; Chen, G.; Zhang, M.; Huang, J.; Chen, B.; Ordonez, A.; Ma, K. Human activities have opposing effects on distributions of narrow-ranged and widespread plant species in China. Proc. Natl. Acad. Sci. USA 2019, 116, 26674–26681. [Google Scholar] [CrossRef] [Green Version]
- Bossu, A.; Marco, A.; Manel, S.; Bertaudière-Montes, V. Effects of built landscape on taxonomic homogenization: Two case studies of private gardens in the French Mediterranean. Landscape Urban Plan. 2014, 129, 12–21. [Google Scholar] [CrossRef]
- Toit, M.J.; Kotze, D.J.; Cilliers, S.S. Quantifying long-term urban grassland dynamics: Biotic homogenization and extinction debts. Sustainability 2020, 12, 1989. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Z.; Wang, Y.; Wang, P.; Xu, Y.; Zhou, Z. Influence of anthropogenic disturbances on understory plant diversity of urban forests in Wuhan, central China. Sains Malays. 2012, 41, 1495–1501. Available online: http://www.ukm.edu.my/jsm/pdf_files/SM-PDF-41-12-2012/01%20Zhen%20Li.pdf (accessed on 9 October 2022).
- Bonthoux, S.; Brun, M.; Pietro, F.D.; Greulich, S.; Bouché-Pillon, S. How can wastelands promote biodiversity in cities? A review. Landscape Urban Plan. 2014, 132, 79–88. [Google Scholar] [CrossRef]
- Deng, H.; Jim, C.Y. Spontaneous plant colonization and bird visits of tropical extensive green roof. Urban Ecosyst. 2017, 20, 337–352. [Google Scholar] [CrossRef]
- Chen, C.; Mao, L.; Qiu, Y.; Cui, J.; Wang, Y. Walls offer potential to improve urban biodiversity. Sci. Rep. 2020, 10, 9905. [Google Scholar] [CrossRef]
- Atha, D.; Alvaresz, R.V.; Chaya, K.; Catusco, J.-P.; Whitaker, E. The spontaneous vascular plant flora of New York’s Central Park. J. Torrey Bot. Soc. 2020, 147, 94–116. [Google Scholar] [CrossRef]
- Laffont-Schwob, I.; Triboit, F.; Prudent, P.; Soulié-Märsche, I.; Rabier, J.; Despréaux, M.; Thiéry, A. Trace metal extraction and biomass production by spontaneous vegetation in temporary Mediterranean stormwater highway retention ponds: Freshwater macroalgae (Chara spp.) vs. cattails (Typha spp.). Ecol. Eng. 2015, 81, 173–181. [Google Scholar] [CrossRef]
- Qian, S.; Qin, D.; Wu, X.; Hu, S.; Hu, L.; Lin, D.; Zhao, L.; Shang, K.; Song, K.; Yang, Y. Urban growth and topographical factors shape patterns of spontaneous plant community diversity in a mountainous city in Southwest China. Urban For. Urban Green. 2020, 55, 126814. [Google Scholar] [CrossRef]
- Šebelíková, L.; Řehounková, K.; Prach, K. Spontaneous revegetation vs. forestry reclamation in post-mining sand pits. Environ. Sci. Pollut. Res. 2016, 23, 13598–13605. [Google Scholar] [CrossRef]
- Tischew, S.; Baasch, A.; Grunert, H.; Kirmer, A. How to develop native plant communities in heavily altered ecosystems: Examples from large-scale surface mining in Germany. Appl. Veg. Sci. 2014, 17, 288–301. [Google Scholar] [CrossRef]
- Šebelíková, L.; Csicsek, G.; Kirmer, A.; Vítovcová, K.; Ortmann-Ajkai, A.; Prach, K.; Řehounková, K. Spontaneous revegetation versus forestry reclamation—Vegetation development in coal mining spoil heaps across Central Europe. Land Degrad. Dev. 2019, 30, 348–356. [Google Scholar] [CrossRef]
- Priede, A.; Mežaka, A.; Dobkeviča, L.; Grīnberga, L. Spontaneous revegetation of cutaway fens: Can it result in valuable habitats? Mires. Peat. 2016, 18, 1–14. [Google Scholar] [CrossRef]
- Gagnon, F.; Rochefort, L.; Lavoie, C. Spontaneous revegetation of a peatland in Manitoba after peat extraction: Diversity of plant assemblages and restoration perspectives. Botany 2018, 96, 779–791. [Google Scholar] [CrossRef]
- Desjardins, D.; Guidi Nissim, W.; Pitre, F.E.; Naud, A.; Labrecque, M. Distribution patterns of spontaneous vegetation and pollution at a former decantation basin in southern Québec, Canada. Ecol. Eng. 2014, 64, 385–390. [Google Scholar] [CrossRef]
- Janečková, P.; Řehounková, K.; Vítovcová, K.; Šebelíková, L.; Prach, K. Spontaneous succession on road verges-an effective approach with minimum effort. Land Degrad. Dev. 2021, 32, 2726–2734. [Google Scholar] [CrossRef]
- Xu, M.; Luo, T.; Wang, Z. Urbanization diverges residents’ landscape preferences but towards a more natural landscape: Case to complement landsenses ecology from the lens of landscape perception. Int. J. Sust. Dev. World. 2020, 27, 250–260. [Google Scholar] [CrossRef]
- Phillips, D.; Lindquist, M. Just weeds? Comparing assessed and perceived biodiversity of urban spontaneous vegetation in informal greenspaces in the context of two American legacy cities. Urban For. Urban Green. 2021, 62, 127151. [Google Scholar] [CrossRef]
- Muratet, A.; Pellegrini, P.; Dufour, A.-B.; Arrif, T.; Chiron, F. Perception and knowledge of plant diversity among urban park users. Landscape Urban Plan. 2015, 137, 95–106. [Google Scholar] [CrossRef]
- Larson, C. China’s vanishing coastal wetlands are nearing critical red line. Science 2015, 350, 489. [Google Scholar] [CrossRef]
- Muhammed, S.N.; Sabiu, N.; Khalil, M.S. An overview of urbanization and its challenges on sustainable development in Nigeria. Dutse J. Pure Appl. Sci. 2015, 1, 19–29. [Google Scholar]
- Padmanaban, R.; Bhowmik, A.K.; Cabral, P.; Zamyatin, A.; Almegdadi, O.; Wang, S. Modelling urban sprawl using remotely sensed data: A case study of Chennai City, Tamilnadu. Entropy 2017, 19, 163. [Google Scholar] [CrossRef] [Green Version]
- Tian, G.; Xu, X.; Liu, X.; Kong, L. The comparison and modeling of the driving factors of urban expansion for thirty-five big cities in the tree regions in China. Adv. Meteorol. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Shen, S. Less and elegant review of Hangzhou Jiangyangfan Ecological Park. Landsc. Archit. 2011, 18, 32–35. [Google Scholar] [CrossRef]
- Yin, L.; Bao, Z.; Yang, F.; Shi, Y. Effects of different plant communities on butterfly diversity in Jiangyangfan Ecological Park. J. Chinese Urban For. 2020, 18, 13–16. [Google Scholar] [CrossRef]
- Wang, X.; Lin, Q. The story of the design of Hangzhou Jiangyangfan Ecological Park. Landsc. Archit. 2011, 18, 18–31. [Google Scholar] [CrossRef]
- Quan, C.; Huang, F.; Fan, L. Study on the resources of plants and dynamic monitoring of landscape in Jiangyangfan Ecological Park. Chin. Landsc. Archit. 2016, 32, 99–102. Available online: https://kns-cnki-net-443.webvpn.zafu.edu.cn/kns8/Detail?sfield=fn&QueryID=9&CurRec=12&recid=&FileName=ZGYL201603022&DbName=CJFDLAST2016&DbCode=CJFD&yx=&pr=&URLID= (accessed on 9 October 2022).
- Quan, C.; Zhang, H.; Gao, S. Study on the secondary vegetation community classification and succession in Jiangyangfan Ecological Park. J. Anhui Agric. Sci. 2022, 50, 110–114. [Google Scholar] [CrossRef]
- Yan, X.; Liu, Q.; Shou, H.; Zeng, H.; Zhang, Y.; Chen, L.; Liu, Y.; Ma, H.; Qi, S.; Ma, J. The categorization and analysis on the geographic distribution patterns of Chinese alien invasive plants. Biodivers. Sci. 2014, 22, 667–676. [Google Scholar] [CrossRef]
- Zhang, S.; Xiao, Y. Life-form and diversity of sexual system of invasive alien plants in China. Bull. Bot. Res. 2013, 33, 351–359. [Google Scholar] [CrossRef]
- Guo, Z.; Zheng, J. Predicting modes of seed dispersal using plant life history traits. Biodivers. Sci. 2017, 25, 966–971. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y. Study on Vascular Plant Diversity and Their Dispersal Mechanisms on the Vertical Surfaces of Nanjing Ming Dynasty City Wall. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, January 2010. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, Z. Major terminologies and concepts in seed dispersal biology. Chinese J. Ecol. 2012, 31, 2397–2403. [Google Scholar] [CrossRef]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. vegan: Community Ecology Package, R Package Version 2.6-2; R Foundation for Statistical Computing: Vienna, Austria, 2022. Available online: https://CRAN.R-project.org/package=vegan(accessed on 9 October 2022).
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.; Deng, X.; Ren, X.; Xiang, W.; Zhang, L.; Ge, R.; Niu, Z.; He, H.; He, L. Biogeographic patterns and influencing factors of the species diversity of tree layer community in typical forest ecosystems in China. Acta Ecol. Sin. 2018, 38, 7727–7738. [Google Scholar] [CrossRef]
- Ferguson, D.E.; Morgan, P.; Johnson, F.D. Proceedings-Land Classifications Based on Vegetation: Applications for Resource Management; U.S. Department of Agriculture, Forest Service: Ogden, UT, USA; Intermountain Research Station: Moscow, Russia, 1989. [CrossRef]
- Huang, H. linkET: Everything is Linkable, R Package Version 0.0.3.3; Linket: Los Angeles, CA, USA, 2021. Available online: https://github.com/Hy4m/linkET(accessed on 9 October 2022).
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Cervelli, E.W.; Lundholm, J.T.; Du, X. Spontaneous urban vegetation and habitat heterogeneity in Xi’an, China. Landscape Urban Plan. 2013, 120, 25–33. [Google Scholar] [CrossRef]
- Gao, Z.; Song, K.; Pan, Y.; Malkinson, D.; Zhang, X.; Jia, B.; Xia, T.; Guo, X.; Liang, H.; Huang, S.; et al. Drivers of spontaneous plant richness patterns in urban green space within a biodiversity hotspot. Urban For. Urban Green. 2021, 61, 127098. [Google Scholar] [CrossRef]
- Li, X.; Fan, S.; Hao, P.; Li, D. Temporal variations of spontaneous plants colonizing in different type of planted vegetation-a case of Beijing Olympic Forest Park. Urban For. Urban Green. 2019, 46, 126459. [Google Scholar] [CrossRef]
- Atha, D.E.; Forrest, T.; Naczi, R.F.C.; Pace, M.C.; Rubin, M.; Schuler, J.A.; Nee, M. The historic and extant spontaneous vascular flora of the New York Botanical Garden. Brittonia 2016, 68, 245–277. [Google Scholar] [CrossRef]
- Peng, X.; Pan, Y.; Zhu, X.; Zhou, X.; Gao, J.; Wang, C. Morphological characters and wind dispersal property of pappose seeds for seven common Compositae weeds in Yunnan, China. J. Yunnan Univ. Nat. Sci. 2018, 40, 1024–1033. [Google Scholar] [CrossRef]
- Zhu, S.; Qin, H.; Chen, Y. Alien species of Compositae in China. Guihaia 2005, 25, 69–76. [Google Scholar]
- Zhang, A.; Hu, X.; Yao, S.; Yu, M.; Ying, Z. Alien, naturalized and invasive plants in China. Plants 2021, 10, 2241. [Google Scholar] [CrossRef]
- Onozuka, M.; Osawa, T. Utilization potential of alien plants in nature-based tourism sites: A case study on Agave americana (century plant) in the Ogasawara Islands. Ecol. Econ. 2022, 195, 107362. [Google Scholar] [CrossRef]
- Wan, J.; Wang, M.; Qin, T.; Bu, X.; Li, H.; Yu, F. Spatial environmental heterogeneity may driver of functional trait variation in Hydrocotyle vulgaris (Araliaceae), an invasive aquatic plant. Aquat. Biol. 2019, 28, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Quan, H.; Dong, B.; Liu, L.; Li, H. The effects of habitat type and nitrogen deposition on the invasion of Hydrocotyle vulgaris in wetland plant communities. Acta Ecol. Sin. 2016, 36, 4045–4054. [Google Scholar] [CrossRef]
- Mohandass, D.; Puyravaud, J.-P.; Hughes, A.C.; Davider, P.; Ganesh, P.S.; Campbell, M. Edge transition impacts on swamp plant communities in the Nilgiri mountains, Southern India. Appl. Ecol. Environ. Res. 2014, 12, 909–929. [Google Scholar] [CrossRef]
- Garbin, M.L.; Carrijo, T.T.; Sansevero, J.B.B.; Sánchez-Tapia, A.; Scarano, F.R. Subordinate, not dominant, woody species promote the diversity of climbing plants. Perspect. Plant Ecol. 2012, 14, 257–265. [Google Scholar] [CrossRef]
- Choi, H.; Kim, H.-T.; Nam, B.E.; Bae, Y.J.; Kim, J.G. Effect of initial planting on vegetation establishment in different depth zones of constructed farm ponds. Restor. Ecol. 2022, 30, 13488. [Google Scholar] [CrossRef]
- Hurtado-M, A.B.; Echeverry-Galvis, M.A.; Salgado-Negret, B.; Munoz, J.C.; Posada, J.M.; Norden, N. Little trace of floristic homogenization in peri-urban Andean secondary forests despite high anthropogenic transformation. J. Ecol. 2021, 109, 1468–1478. [Google Scholar] [CrossRef]
- Meerbeek, K.V.; Muys, B.; Schowanek, S.D.; Svenning, J.-C. Reconciling conflicting paradigms of biodiversity conservation: Human intervention and rewilding. Bioscience 2019, 69, 997–1007. [Google Scholar] [CrossRef]
- Kim, M.; Lee, S.; Lee, S.; Yi, K.; Kim, H.-S.; Chung, S.; Chung, J.; Kim, H.S.; Yoon, T.K. Seed dispersal models for natural regeneration: A review and prospects. Forests 2022, 13, 659. [Google Scholar] [CrossRef]
- Beckman, N.G.; Neuhauser, C.; Muller-Landau, H.C. The interacting effects of clumped seed dispersal and distance- and density-dependent mortality on seedling recruitment patterns. J. Ecol. 2012, 100, 862–873. [Google Scholar] [CrossRef]
- Ma, W.; Yan, S.; Wang, Y.; Wang, Z.; Chen, Y. Study on seed dispersal modes and landscape application strategies of spontaneous plants in street tree planting ponds: Taking Fuzhou City as an example. J. Sichuan For. Sci. Technol. 2021, 42, 107–114. [Google Scholar] [CrossRef]
- Zhou, Q.; Liu, Z.; Xin, Z.; Daryanto, S.; Wang, L.; Qian, J.; Wang, Y.; Liang, W.; Qin, X.; Zhao, Y.; et al. Relationship between seed morphological traits and wind dispersal trajectory. Funct. Plant Biol. 2019, 46, 1063–1071. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Q.; Li, Z. Seed dispersal modes and landscape application strategies of autogenesis herbs in the parks of Nanjing City. Chin. Landsc. Archit. 2020, 36, 119–123. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Naoe, S.; Masaki, T.; Isagi, Y. Temporal variations in seed dispersal patterns of a bird-dispersed tree, Swida controversa (Cornaceae), in a temperate forest. Ecol. Res. 2016, 31, 165–176. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Yan, S.; Zhu, J.; Liu, D.; Liao, Z.; Li, C.; Liu, Q. Influences of Phragmites australis density and groundwater level on soil water in semiarid wetland, North China: Which is more influential? Ecohydrol. Hydrobiol. 2022, 22, 85–95. [Google Scholar] [CrossRef]
- Li, X.; Liang, T.; Liu, H. How plants coordinate their development in response to light and temperature signals. Plant Cell. 2021, 34, 955–966. [Google Scholar] [CrossRef]
- Fitch, G.; Vandermeer, J.H. Light availability influences the intensity of nectar robbery and its effects on reproduction in a tropical shrub via multiple pathways. Am. J. Bot. 2020, 107, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, S.D.; Castillo, J.A. Influence of light on plant-phyllosphere interaction. Front. Plant Sci. 2018, 9, 1482. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Song, J.; Jeong, B.R. Lighting from top and side enhances photosynthesis and plant performance by improving light usage efficiency. Int. J. Mol. Sci. 2022, 23, 2448. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zheng, L.; Pan, X.; Li, W.; Kang, X.; Li, J.; Ning, Y.; Zhang, M.; Cui, L. Hydrological conditions affect the interspecific interaction between two emergent wetland species. Front. Plant Sci. 2017, 8, 2253. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, A.H. Nitrogen and phosphorus differentially affect annual and perennial plants in tidal freshwater and oligohaline wetlands. Estuar. Coast. 2013, 36, 547–558. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Type | Microhabitat Type | Feature Description | Artificial Interference Intensity | Number of Quadrats |
|---|---|---|---|---|
| Roadside vegetation area | Cultivated vegetation | Artificially cultivated ornamental vegetation ≤5 m from roadside | High intensity, artificial planting to create seasonal landscape | 24 |
| Semi-cultivated vegetation | Semiartificial vegetation ≤5 m from roadside | Medium intensity, appropriate artificial planting | 42 | |
| Ecological restoration area | Swamp | Long-term overwet land under the surface, with water depth between 0 m and 0.5 m | Low intensity, unattended area | 183 |
| Secondary forest | Woodland mainly covered by spontaneous plant communities | Low intensity, unattended area | 16 | |
| Habitat island | – | Area enclosed by weather-resistant steel | Low intensity, unattended area | 20 |
| Drainage area | Shallow-water area | Water with depth between 0.5 m and 1 m | Medium intensity, appropriate artificial planting | 22 |
| Deep-water area | Water with depth >1 m | Medium intensity, proper water quality maintenance | 27 | |
| Total | 334 |
| Environmental Factor | Habitat | Microhabitat | ||||||
|---|---|---|---|---|---|---|---|---|
| Unique 1 | Average Share 2 | Individual 3 | I Perc (%) 4 | Unique | Average Share | Individual | I Perc (%) | |
| Soil temperature | −0.0025 | 5 × 10−4 | −0.002 | −5.26 | −0.0028 | 3 × 10−4 | −0.0025 | −5.95 |
| Soil moisture | 9 × 10−4 | 7 × 10−4 | 0.0016 | 4.21 | 0.0018 | 5 × 10−4 | 0.0023 | 5.48 |
| Soil pH | −0.0029 | −1 × 10−4 | −0.003 | −7.89 | −0.0026 | 1 × 10−4 | −0.0025 | −5.95 |
| Slope | −0.002 | 8 × 10−4 | −0.0012 | −3.16 | −0.0028 | 9 × 10−4 | −0.0019 | −4.52 |
| Aspect | 0.0031 | 0.0066 | 0.0097 | 25.53 | −0.001 | 0.011 | 0.01 | 23.81 |
| Water pH | 0.0012 | 0.0015 | 0.0027 | 7.11 | 7 × 10−4 | 0.002 | 0.0027 | 6.43 |
| Water depth | 0.0209 | 0.0084 | 0.0293 | 77.11 | 0.0136 | 0.0149 | 0.0285 | 67.86 |
| Lux | −0.0013 | −5 × 10−4 | −0.0018 | −4.74 | −7 × 10−4 | −8 × 10−4 | −0.0015 | −3.57 |
| Habitat type | 0.0016 | 0.0023 | 0.0039 | 10.26 | 0.0039 | 0.0028 | 0.0067 | 15.95 |
| CP Simpson | −0.0029 | 0.0012 | −0.0017 | −4.47 | −0.003 | 0.0013 | −0.0017 | −4.05 |
| CP clustering degree | −8 × 10−4 | 0.0014 | 6 × 10−4 | 1.58 | 0 | 0.0019 | 0.0019 | 4.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Xu, B.; Yu, Q.; Fan, L.; Guo, T.; Fu, D.; Chen, H.; Yan, H.; Shao, F.; Li, X. Distribution Pattern and Factors Influencing Spontaneous Plant Diversity in Different Wetland Habitats. Forests 2022, 13, 1678. https://doi.org/10.3390/f13101678
Yang Y, Xu B, Yu Q, Fan L, Guo T, Fu D, Chen H, Yan H, Shao F, Li X. Distribution Pattern and Factors Influencing Spontaneous Plant Diversity in Different Wetland Habitats. Forests. 2022; 13(10):1678. https://doi.org/10.3390/f13101678
Chicago/Turabian StyleYang, Yifan, Bin Xu, Qingqing Yu, Likun Fan, Tingting Guo, Dongshi Fu, Hao Chen, Hai Yan, Feng Shao, and Xiaopeng Li. 2022. "Distribution Pattern and Factors Influencing Spontaneous Plant Diversity in Different Wetland Habitats" Forests 13, no. 10: 1678. https://doi.org/10.3390/f13101678
APA StyleYang, Y., Xu, B., Yu, Q., Fan, L., Guo, T., Fu, D., Chen, H., Yan, H., Shao, F., & Li, X. (2022). Distribution Pattern and Factors Influencing Spontaneous Plant Diversity in Different Wetland Habitats. Forests, 13(10), 1678. https://doi.org/10.3390/f13101678