


Water Storage and Use by Platycladus orientalis under Different Rainfall Conditions in the Rocky Mountainous Area of Northern China

Abstract
:1. Introduction
2. Materials and Methods
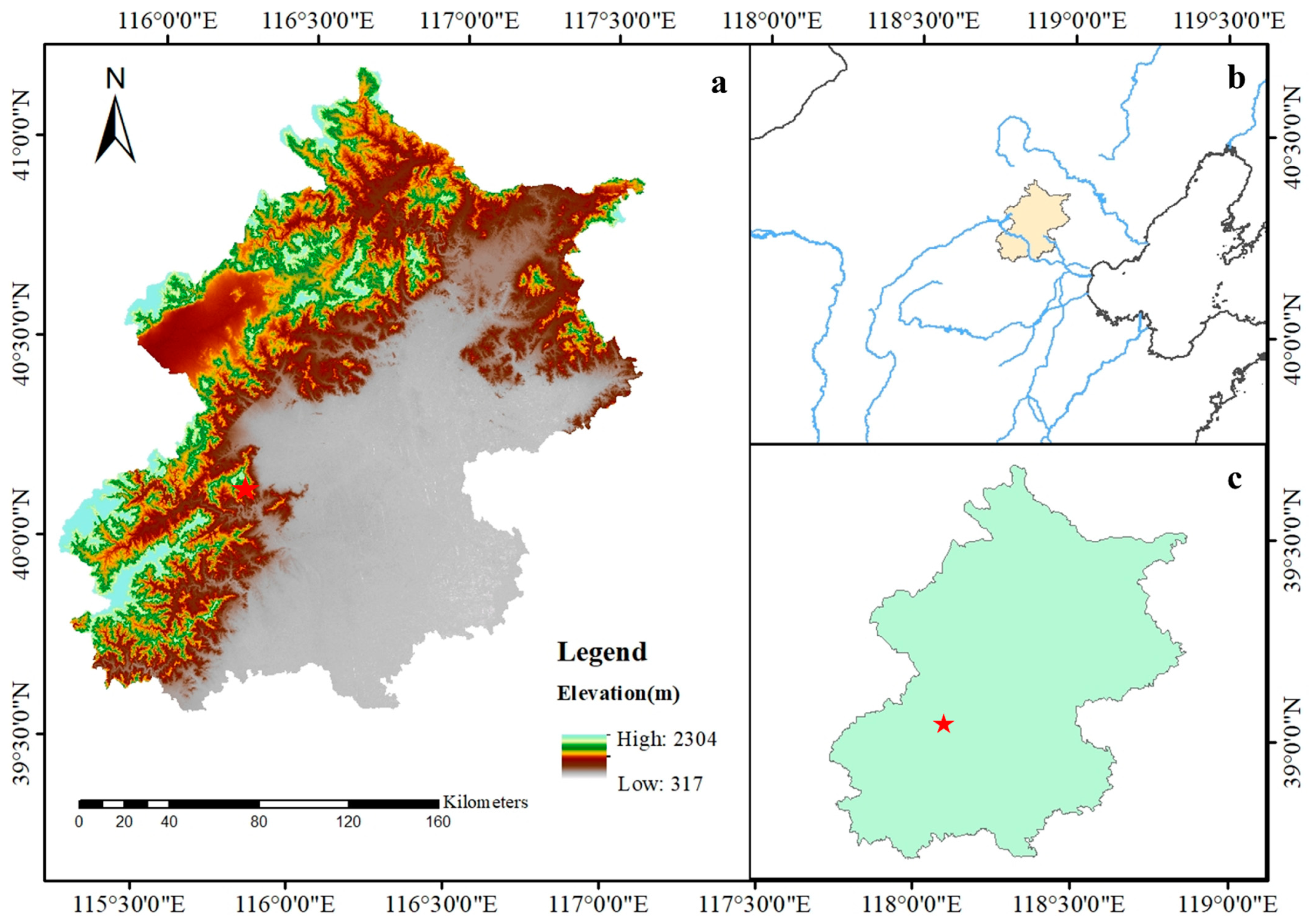
2.1. Study Site
2.2. Experimental Design
2.3. Measurement and Calculation of Tree Sap Flow
2.4. Hydrometeorological Measurements
2.5. Determination of the Water Source
2.6. Stable Isotope Analysis
2.7. Data Analysis
3. Results
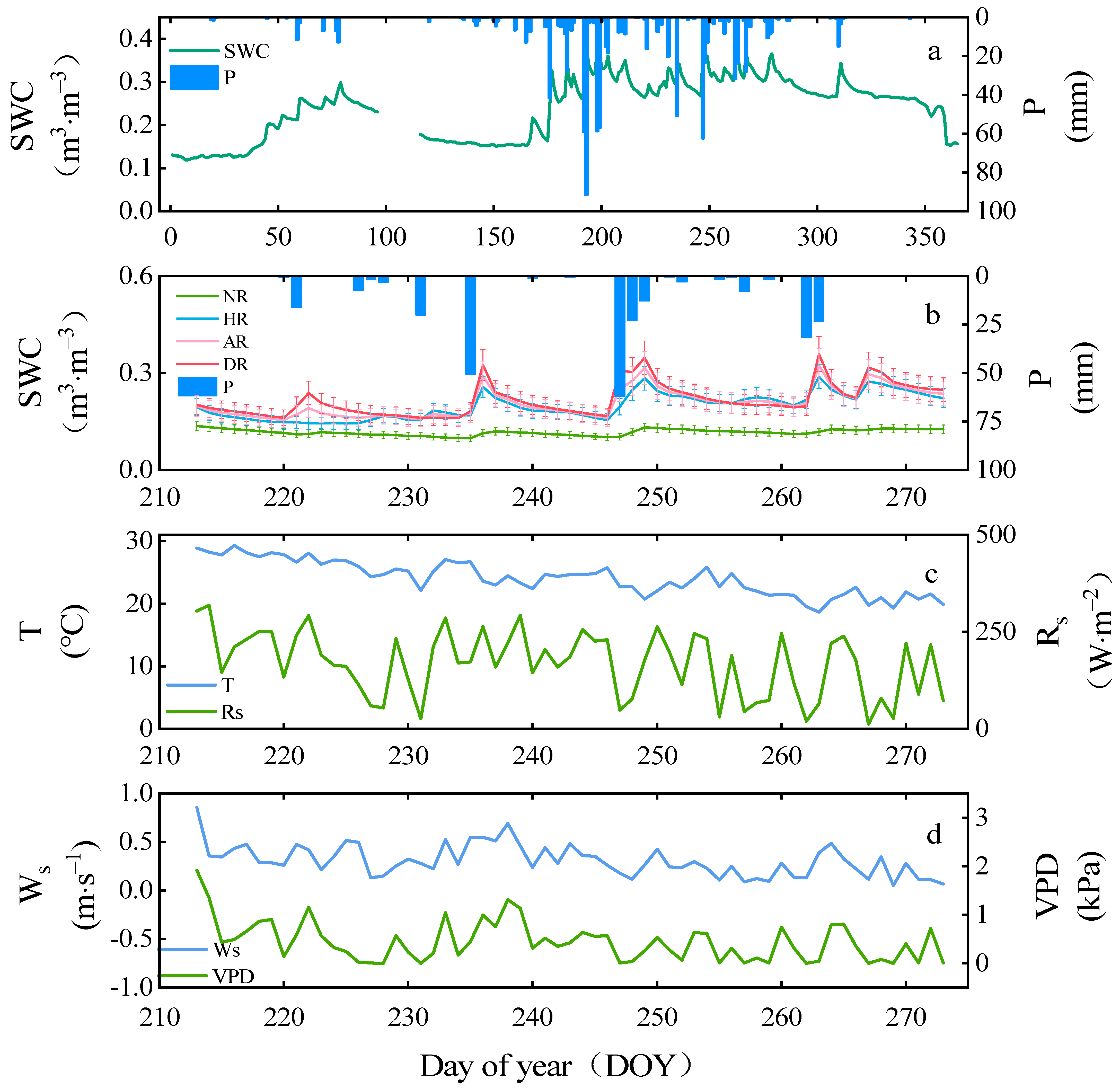
3.1. Environmental Data
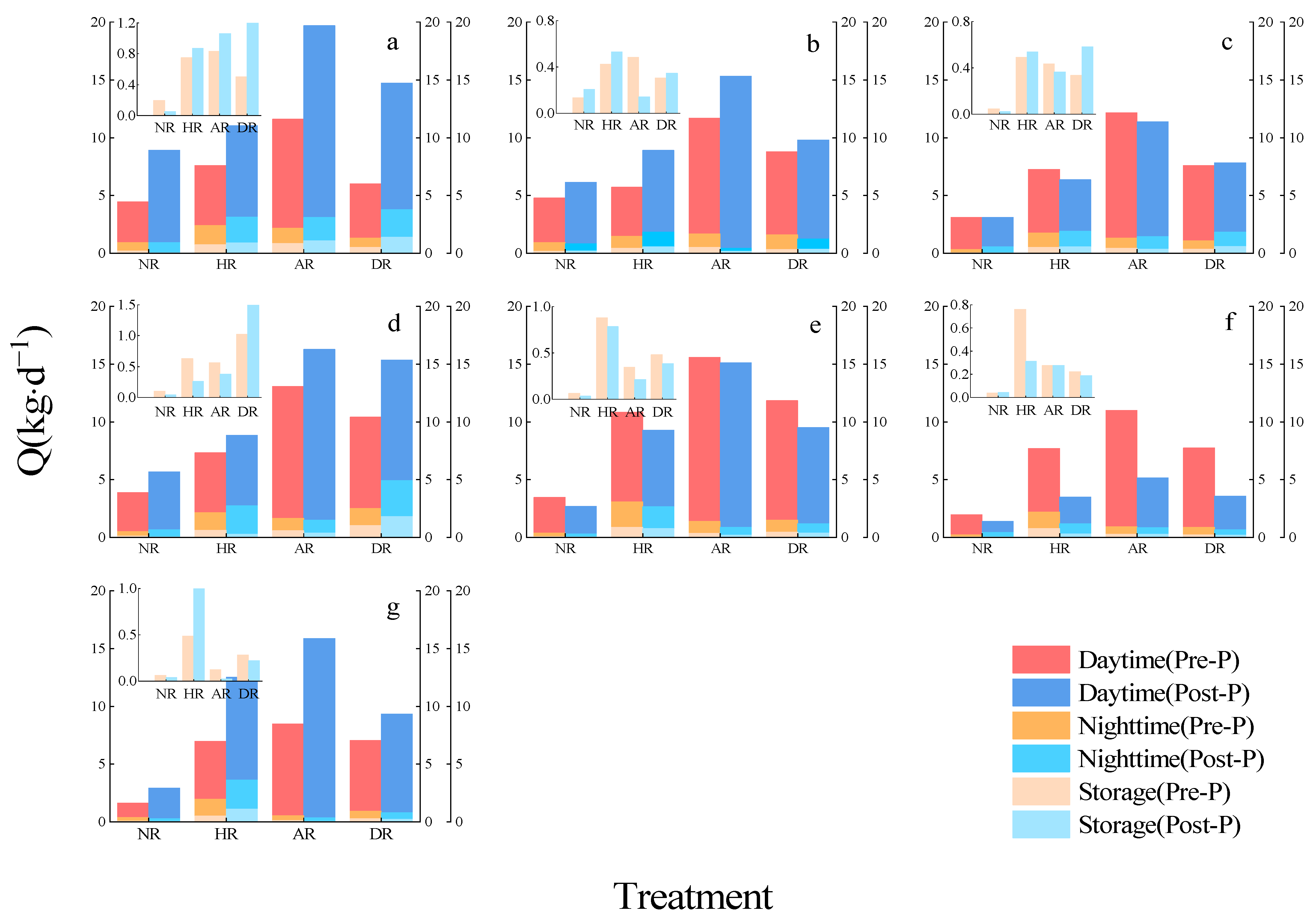
3.2. Time Lag and Calculation of Tree Water Storage
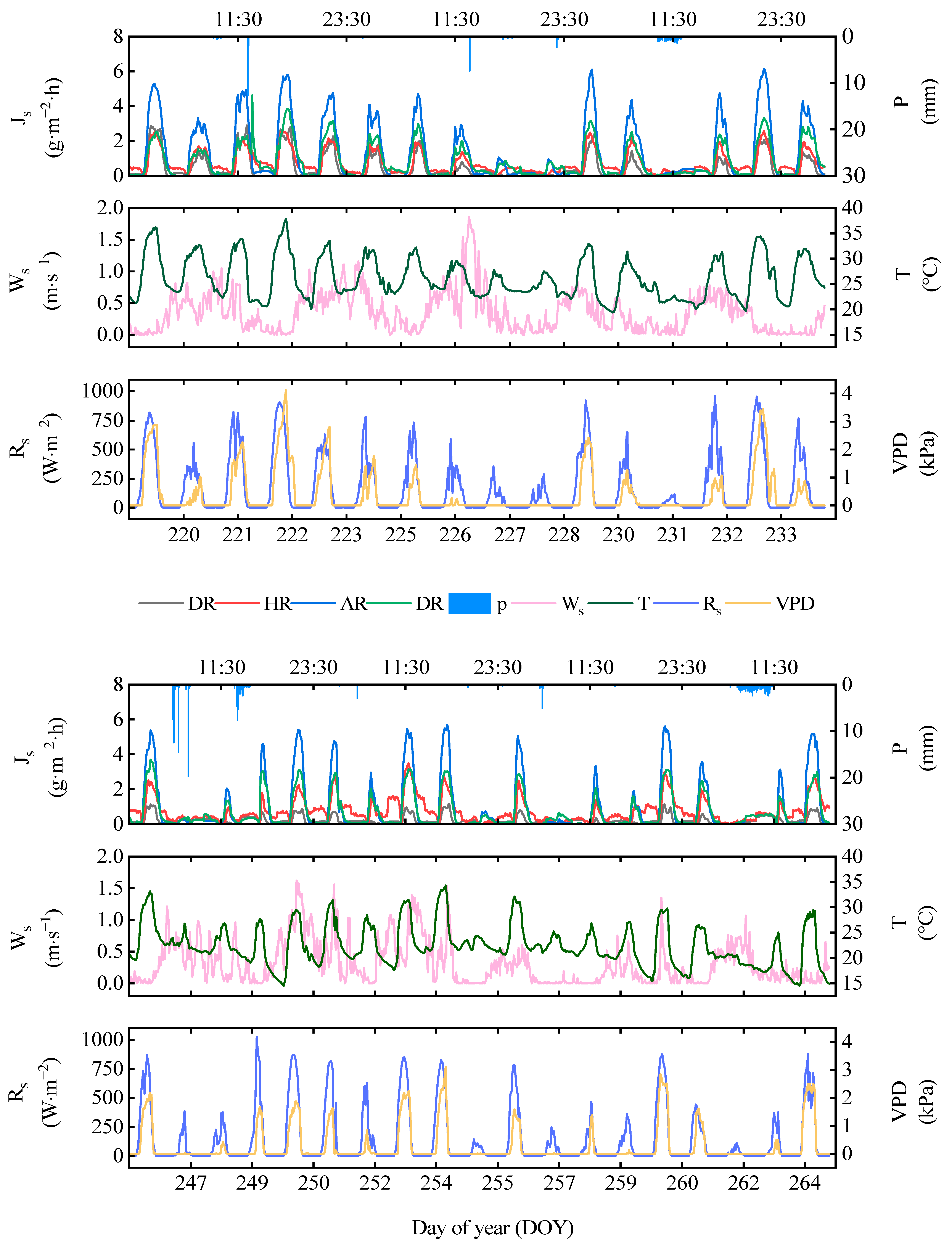
3.3. Characteristics of Sap Flow Changes
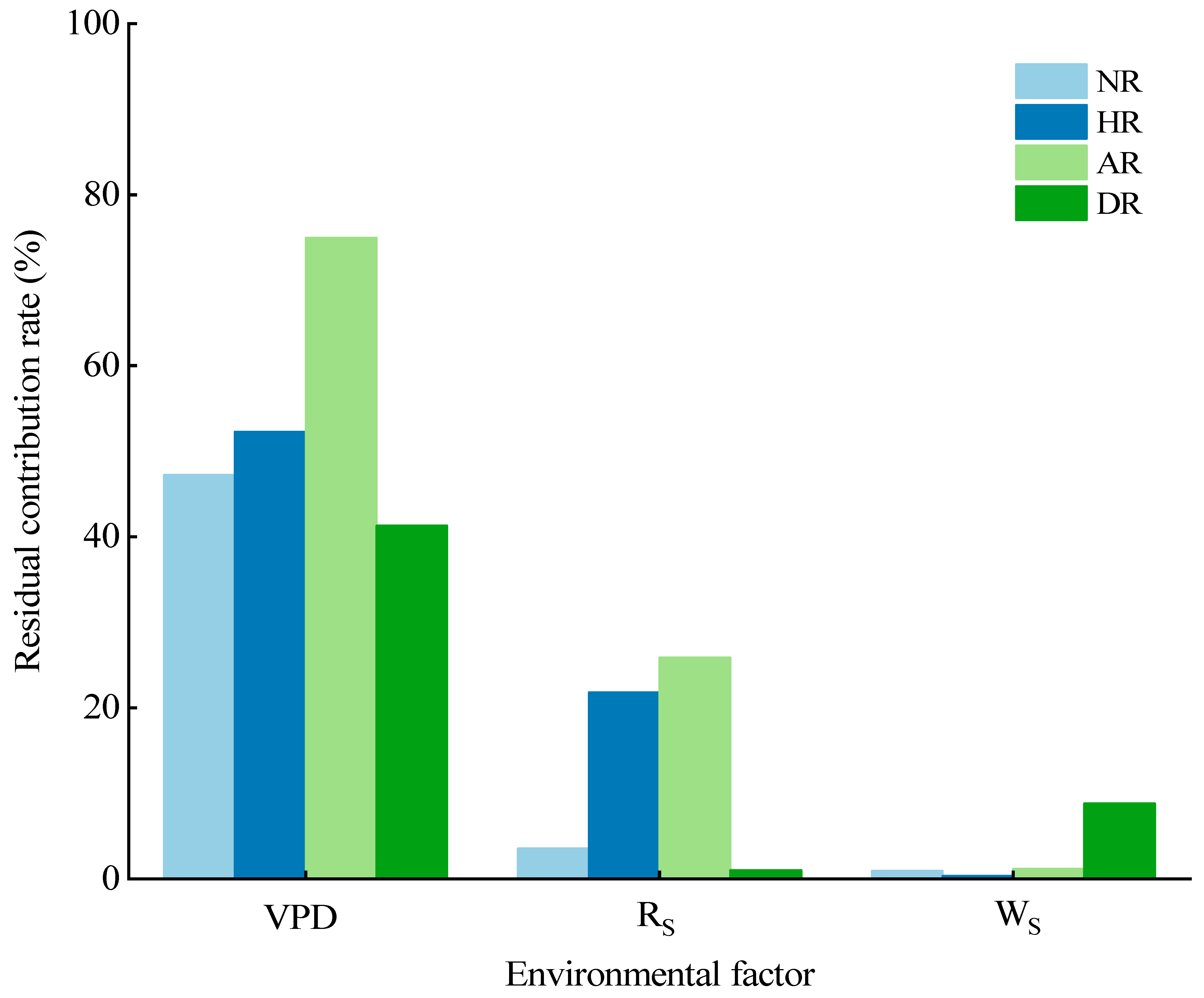
3.4. Response of Sap Flow to Environmental Factors
4. Discussion
4.1. Influence of Precipitation on Sap Flow
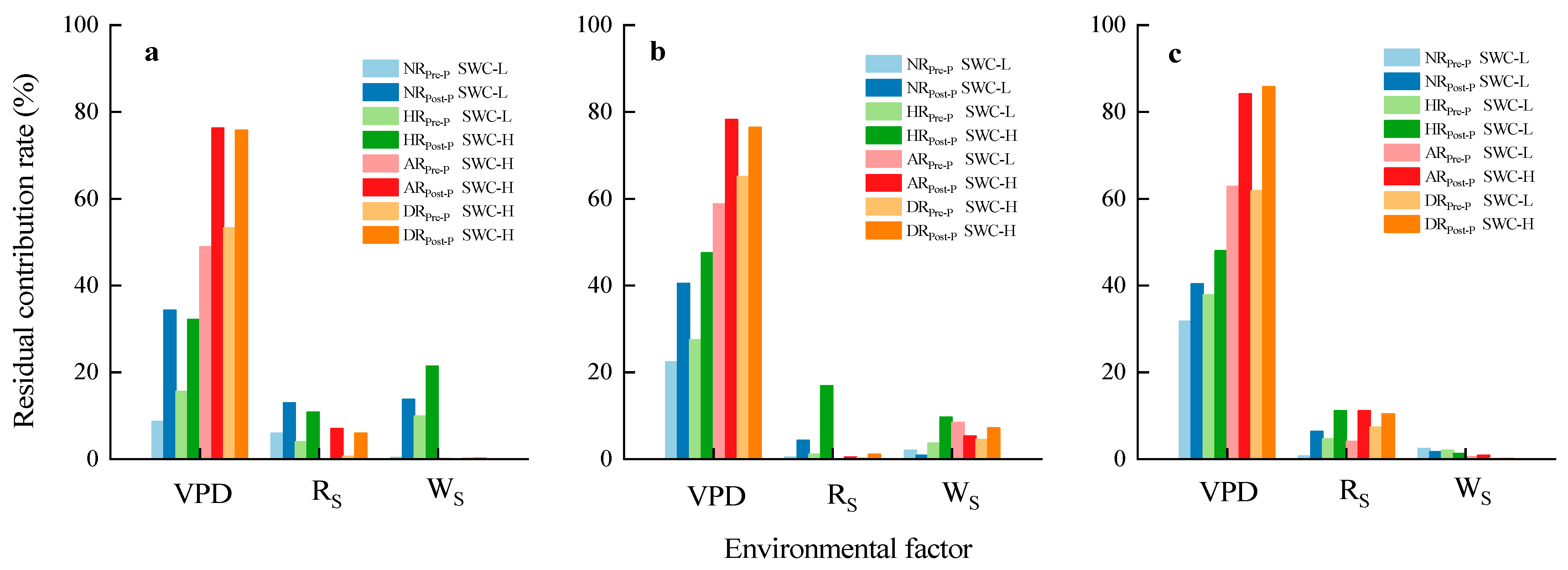
4.2. Influence of Precipitation Treatments on the Response of Sap Flow to Environmental Factors
4.3. Water Resources
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiong, Y.J.; Zhao, S.H.; Tian, F.; Qiu, G.Y. An Evapotranspiration Product for Arid Regions Based on the Three-Temperature Model and Thermal Remote Sensing. J. Hydrol. 2015, 530, 392–404. [Google Scholar] [CrossRef]
- McCabe, M.F.; Wood, E.F. Scale Influences on the Remote Estimation of Evapotranspiration Using Multiple Satellite Sensors. Remote Sens. Environ. 2006, 105, 271–285. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, D.; Zhang, X.; Tian, J.; Li, C. Impacts of Vegetation Changes on Global Evapotranspiration in the Period 2003–2017. Acta Geogr. Sin. 2021, 76, 584–594. [Google Scholar] [CrossRef]
- Zhang, J.G.; He, Q.Y.; Shi, W.Y.; Otsuki, K.; Yamanaka, N.; Du, S. Radial Variations in Xylem Sap Flow and Their Effect on Whole-tree Water Use Estimates. Hydrol. Process. 2016, 29, 4993–5002. [Google Scholar] [CrossRef]
- Di, N.; Xi, B.; Clothier, B.; Wang, Y.; Li, G.; Jia, L. Diurnal and Nocturnal Transpiration Behaviors and Their Responses to Groundwater-Table Fluctuations and Meteorological Factors of Populus Tomentosa in the North China Plain. For. Ecol. Manag. 2019, 448, 445–456. [Google Scholar] [CrossRef]
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial Water Fluxes Dominated by Transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, P.; Wang, Q.; Cai, X.; Ma, L.; Rao, X.; Zeng, X. Nocturnal Sap Flow Characteristics and Stem Water Recharge of Acacia Mangium. Front. For. China 2008, 3, 72–78. [Google Scholar] [CrossRef]
- Hubbart, J.A.; Kavanagh, K.L.; Pangle, R.; Link, T.; Schotzko, A. Cold Air Drainage and Modeled Nocturnal Leaf Water Potential in Complex Forested Terrain. Tree Physiol. 2007, 27, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Poyatos, R.; Granda, V.; Flo, V.; Adams, M.A.; Adorjan, B.; Aguade, D.; Aidar, M.P.M.; Allen, S.; Susana Alvarado-Barrientos, M.; Anderson-Teixeira, K.J.; et al. Global Transpiration Data from Sap Flow Measurements: The SAPFLUXNET Database. Earth Syst. Sci. Data 2021, 13, 2607–2649. [Google Scholar] [CrossRef]
- Gartner, K.; Nadezhdina, N.; Englisch, M.; Cermak, J.; Leitgeb, E. Sap Flow of Birch and Norway Spruce during the European Heat and Drought in Summer 2003. For. Ecol. Manag. 2009, 258, 590–599. [Google Scholar] [CrossRef]
- Zeppel, M.J.B.; Lewis, J.D.; Phillips, N.G.; Tissue, D.T. Consequences of Nocturnal Water Loss: A Synthesis of Regulating Factors and Implications for Capacitance, Embolism and Use in Models. Tree Physiol. 2014, 34, 1047–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Levia, D.F.; He, B.; Wu, H.; Fu, C. Interspecific Variation in Tree- and Stand-Scale Stemflow Funneling Ratios in a Subtropical Deciduous Forest in Eastern China. J. Hydrol. 2020, 590. [Google Scholar] [CrossRef]
- Benyon, R.G. Nighttime Water Use in an Irrigated Eucalyptus Grandis Plantation. Tree Physiol. 1999, 19, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.E.; Burgess, S.S.O.; Tu, K.P.; Oliveira, R.S.; Santiago, L.S.; Fisher, J.B.; Simonin, K.A.; Ambrose, A.R. Nighttime Transpiration in Woody Plants from Contrasting Ecosystems. Tree Physiol. 2007, 27, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Forster, M.A. How Significant Is Nocturnal Sap Flow? Tree Physiol. 2014, 34, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhao, P.; Cai, X.; ma, L.; Rao, X.; Zeng, X.; Wang, Q. Partition of Nocturnal Sap Flow in Acacia Mangium and Its Implication for Estimating the Whole-Tree Transpiration. Front. For. China 2009, 4, 191–200. [Google Scholar] [CrossRef]
- Siddiq, Z.; Chen, Y.-J.; Zhang, Y.-J.; Zhang, J.-L.; Cao, K.-F. More Sensitive Response of Crown Conductance to VPD and Larger Water Consumption in Tropical Evergreen than in Deciduous Broadleaf Timber Trees. Agric. For. Meteorol. 2017, 247, 399–407. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Meinzer, F.C.; Hinojosa, J.A.; Hoffmann, W.A.; Franco, A.C. Processes Preventing Nocturnal Equilibration between Leaf and Soil Water Potential in Tropical Savanna Woody Species. Tree Physiol. 2004, 24, 1119–1127. [Google Scholar] [CrossRef]
- Tie, Q.; Hu, H.; Tian, F.; Guan, H.; Lin, H. Environmental and Physiological Controls on Sap Flow in a Subhumid Mountainous Catchment in North China. Agric. For. Meteorol. 2017, 240–241, 46–57. [Google Scholar] [CrossRef]
- Resco de Dios, V.; Loik, M.E.; Smith, R.; Aspinwall, M.J.; Tissue, D.T. Genetic Variation in Circadian Regulation of Nocturnal Stomatal Conductance Enhances Carbon Assimilation and Growth. Plant Cell Environ. 2016, 39, 3–11. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Z.; Sun, G.; Chen, L.; Xu, H.; Chen, S. Biophysical Controls on Nocturnal Sap Flow in Plantation Forests in a Semiarid Region of Northern China. Agric. For. Meteorol. 2020, 284, 107904. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Q.; Wei, Z.; Yu, X.; Jia, G.; Jiang, J. Partitioning Tree Water Usage into Storage and Transpiration in a Mixed Forest. For. Ecosyst. 2021, 8, 72. [Google Scholar] [CrossRef]
- McCormick, E.L.; Dralle, D.N.; Hahm, W.J.; Tune, A.K.; Schmidt, L.M.; Chadwick, K.D.; Rempe, D.M. Widespread Woody Plant Use of Water Stored in Bedrock. Nature 2021, 597, 225–229. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, W.; Zheng, X.; Li, S. Root Distribution of Nitraria Sibirica with Seasonally Varying Water Sources in a Desert Habitat. J. Plant Res. 2015, 128, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Guerrieri, R.; Belmecheri, S.; Ollinger, S.V.; Asbjornsen, H.; Jennings, K.; Xiao, J.; Stocker, B.D.; Martin, M.; Hollinger, D.Y.; Bracho-Garrillo, R.; et al. Disentangling the Role of Photosynthesis and Stomatal Conductance on Rising Forest Water-Use Efficiency. Proc. Natl. Acad. Sci. USA 2019, 116, 16909–16914. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Lin, G.; Huang, J.; Jenerette, G.D. Dependence of Carbon Sequestration on the Differential Responses of Ecosystem Photosynthesis and Respiration to Rain Pulses in a Semiarid Steppe. Glob. Change Biol. 2009, 15, 2450–2461. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Si, J.H.; Feng, Q.; Yu, T.F.; Li, P.D. Comparative Study of Daytime and Nighttime Sap Flow of Populus Euphratica. Plant Growth Regul. 2017, 82, 353–362. [Google Scholar] [CrossRef]
- Li, S.; Lu, S.; Zhao, Y.; Zhao, N.; Chen, B. Characteristics and Influencing Factors in Sap Flow of Four Broadleaved Tree Species for Typical Weather Conditions of Beijing. J. Ecol. Rural. Environ. 2019, 35, 189–196. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Wang, Y.; Wang, Y.; Liu, C.; Yang, P.; Pan, S. Characteristics of daily sap flow for typical species in Jinyun Mountain of Chongqing in relation to meteorological factors. J. Beijing For. Univ. 2016, 38, 11–20. [Google Scholar] [CrossRef]
- Xia, Y.; Zhang, X.; Dai, J.; Wang, Y.; Luo, Z. Response of Stem Sap Flow of Cinnamomum camphora to Precipitation Under Different Environments in the Subtropical Monsoon Region. Res. Soil Water Conserv. 2021, 28, 144–152. [Google Scholar] [CrossRef]
- Ivans, S.; Hipps, L.; Leffler, A.J.; Ivans, C.Y. Response of Water Vapor and CO2 Fluxes in Semiarid Lands to Seasonal and Intermittent Precipitation Pulses. J. Hydrometeorol. 2006, 7, 995–1010. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, B. The Response of Sap Flow in Shrubs to Rainfall Pulses in the Desert Region of China. Agric. For. Meteorol. 2010, 150, 1297–1306. [Google Scholar] [CrossRef]
- Fravolini, A.; Hultine, K.R.; Brugnoli, E.; Gazal, R.; English, N.B.; Williams, D.G. Precipitation Pulse Use by an Invasive Woody Legume: The Role of Soil Texture and Pulse Size. Oecologia 2005, 144, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K.; Briggs, J.M.; Collins, S.L.; Archer, S.R.; Bret-Harte, M.S.; Ewers, B.E.; Peters, D.P.; Young, D.R.; Shaver, G.R.; Pendall, E. Shrub Encroachment in North American Grasslands: Shifts in Growth Form Dominance Rapidly Alters Control of Ecosystem Carbon Inputs. Glob. Change Biol. 2010, 14, 615–623. [Google Scholar] [CrossRef]
- Wightman, M.G.; Martin, T.A.; Gonzalez-Benecke, C.A.; Jokela, E.J.; Cropper, W.P.; Ward, E.J. Loblolly Pine Productivity and Water Relations in Response to Throughfall Reduction and Fertilizer Application on a Poorly Drained Site in Northern Florida. Forests 2016, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Besson, C.K.; Lobo-do-Vale, R.; Rodrigues, M.L.; Almeida, P.; Herd, A.; Grant, O.M.; David, T.S.; Schmidt, M.; Otieno, D.; Keenan, T.F.; et al. Cork Oak Physiological Responses to Manipulated Water Availability in a Mediterranean Woodland. Agric. For. Meteorol. 2014, 184, 230–242. [Google Scholar] [CrossRef]
- Porporato, A.; Daly, E.; Rodriguez-Iturbe, I. Soil Water Balance and Ecosystem Response to Climate Change. Am. Nat. 2004, 164, 625. [Google Scholar] [CrossRef]
- Ehleringer, W.J.R. Intra- and Interspecific Variation for Summer Precipitation Use in Pinyon-Juniper Woodlands. Ecol. Monogr. 2000, 70, 517–537. [Google Scholar] [CrossRef]
- Jackson, P.C.; Cavelier, J.; Goldstein, G.; Meinzer, F.C.; Holbrook, N.M. Partitioning of Water Resources among Plants of a Lowland Tropical Forest. Oecologia 1995, 101, 197–203. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Andrade, J.L.; Goldstein, G.; Holbrook, N.M.; Wright, J. Caveliers.J. Partitioning of Soil Water among Canopy Trees in a Seasonally Dry Tropical Forest. Oecologia 1999, 121, 293–301. [Google Scholar] [CrossRef]
- Schwendenmann, L.; Pendall, E.; Sanchez-Bragado, R.; Kunert, N.; Hölscher, D. Tree Water Uptake in a Tropical Plantation Varying in Tree Diversity: Interspecific Differences, Seasonal Shifts and Complementarity. Ecohydrology 2014, 8, 1–12. [Google Scholar] [CrossRef]
- Wang, X.; Jia, G.; Deng, W.; Liu, Z.; Liu, Z.; Que, G.; Li, L. Long-term Water Use Characteristics and Patterns of Typical Tree Species in Seasonal Drought Regions. Chin. J. Appl. Ecol. 2021, 32, 1943–1950. [Google Scholar] [CrossRef]
- Liu, W.; Jia, G. Studies of Platycladus orientalis Stomatal Conductance and Its Regulation on Canopy Transpiration in Beijng Mountainous Area. Master’s Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Liu, Z.; Yu, X.; Jia, G.; Li, H.; Lu, W.; Hou, G. Response to Precipitation in Water Sources for Platycladus orientalis in Beijing Mountatin Area. Sci. Silvae Sin. 2018, 54, 16–23. [Google Scholar] [CrossRef]
- Tognetti, R.; Giovannelli, A.; Lavini, A.; Morelli, G.; Fragnito, F.; d’Andria, R. Assessing Environmental Controls over Conductances through the Soil–Plant–Atmosphere Continuum in an Experimental Olive Tree Plantation of Southern Italy. Agric. For. Meteorol. 2009, 149, 1229–1243. [Google Scholar] [CrossRef]
- Martín-Gómez, P.; Barbeta, A.; Voltas, J.; Peñuelas, J.; Dennis, K.; Palacio, S.; Dawson, T.E.; Ferrio, J.P. Isotope-Ratio Infrared Spectroscopy: A Reliable Tool for the Investigation of Plant-Water Sources? New Phytol. 2015, 207, 914–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, N.M.; Griffis, T.J.; Lee, X.; Baker, J.M. Identification and Correction of Spectral Contamination in 2H/1H and 18O/16O Measured in Leaf, Stem, and Soil Water. Rapid Commun. Mass Spectrom. 2011, 25, 3360–3368. [Google Scholar] [CrossRef]
- He, Q.-Y.; Yan, M.-J.; Miyazawa, Y.; Chen, Q.-W.; Cheng, R.-R.; Otsuki, K.; Yamanaka, N.; Du, S. Sap Flow Changes and Climatic Responses over Multiple-Year Treatment of Rainfall Exclusion in a Sub-Humid Black Locust Plantation. For. Ecol. Manag. 2020, 457, 117730. [Google Scholar] [CrossRef]
- Limousin, J.M.; Rambal, S.; Ourcival, J.M.; Rocheteau, A.; Joffre, R.; Rodriguez-Cortina, R. Long-Term Transpiration Change with Rainfall Decline in a Mediterranean Quercus Ilex Forest. Glob. Change Biol. 2009. [Google Scholar] [CrossRef]
- Wu, X.; Chen, Y.; Tang, Y. Sap flow characteristics and its responses to precipitation in Robinia pseudoacacia and Platycladus orientalis plantations. Chin. J. Plant Ecol. 2015, 39, 1176–1187. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.-Q.; Xi, B.-Y.; Liu, J.-Q.; Liu, Y.; Zou, S.-Y.; Song, W.-Y.; Chen, L.-X. Transpiration Process and Environmental Response of Poplar Plantation under Different Irrigation Condition. Chin. J. Plant Ecol. 2021, 45, 370–382. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, X. Water Migration Process and Utilization Mechanism of Typcial Trees in North China. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Brinkmann, N.; Eugster, W.; Zweifel, R.; Buchmann, N.; Kahmen, A. Temperate Tree Species Show Identical Response in Tree Water Deficit but Different Sensitivities in Sap Flow to Summer Soil Drying. Tree Physiol. 2016, 36, 1508–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ding, Y.; Chen, H.; Wang, K.; Nie, Y. Responses of Sap Flow to Natural Rainfall and Continuous Drought of Tree Species Growing on Bedrock Outcrops. Chin. J. Appl. Ecol. 2018, 29, 1117–1124. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Johnson, D.M.; Lachenbruch, B.; McCulloh, K.A.; Woodruff, D.R. Xylem Hydraulic Safety Margins in Woody Plants: Coordination of Stomatal Control of Xylem Tension with Hydraulic Capacitance. Funct. Ecol. 2009, 23, 922–930. [Google Scholar] [CrossRef]
- Yi, K.; Dragoni, D.; Phillips, R.P.; Roman, D.T.; Novick, K.A. Dynamics of Stem Water Uptake among Isohydric and Anisohydric Species Experiencing a Severe Drought. Tree Physiol. 2017, 37, 1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcculloh, K.A.; Johnson, D.M.; Meinzer, F.C.; Woodruff, D.R. The Dynamic Pipeline: Hydraulic Capacitance and Xylem Hydraulic Safety in Four Tall Conifer Species. Plant Cell Environ. 2014, 37, 1171–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, S.R. Radiation Balance, Transpiration and Photosynthesis of an Isolated Tree. Agric. For. Meteorol. 1993, 64, 201–221. [Google Scholar] [CrossRef]
- Bosch, D.D.; Marshall, L.K.; Teskey, R. Forest Transpiration from Sap Flux Density Measurements in a Southeastern Coastal Plain Riparian Buffer System. Agric. For. Meteorol. 2014, 187, 72–82. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Y.; Liu, S.; Wei, X.; Wang, X. Response of Relative Sap Flow to Meteorological Factors under Different Soil Moisture Conditions in Rainfed Jujube (Ziziphus Jujuba Mill.) Plantations in Semiarid Northwest China. Agric. Water Manag. 2014, 136, 23–33. [Google Scholar] [CrossRef]
- Ma, C.; Luo, Y.; Shao, M.; Li, X.; Sun, L.; Jia, X. Environmental Controls on Sap Flow in Black Locust Forest in Loess Plateau, China. Sci. Rep. 2017, 7, 13160. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Fu, S.; Chen, D.; Zheng, S.; Wang, T.; Bai, Y. Grapevine Sap Flow in Response to Physio-Environmental Factors under Solar Greenhouse Conditions. Water 2020, 12, 3081. [Google Scholar] [CrossRef]
- Fan, B.; Liu, Z.; Xiong, K.; Li, Y.; Li, K.; Yu, X. Influence of Environmental Factors on the Sap Flow Activity of the Golden Pear in the Growth Period of Karst Area in Southern China. Water 2022, 14, 1707. [Google Scholar] [CrossRef]
- Lv, J.; He, Q.; Yan, M.-J.; li, G.; Du, S. Sap Flow Characteristics of Quercus liaotungensis in Response to Sapwood Area and Soil Moisture in the Loess Hilly Region, China. Chin. J. Appl. Ecol. 2018, 29, 725–731. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, J.; Zhou, M.; Zhao, P.; Liu, B. Response of Sap Flow of Betula platyphylla to Soil Moisture in Southern Greater Xing’an Mountains. Res. Soil Water Conserv. 2020, 27, 128–133. [Google Scholar] [CrossRef]
- Granier, A.; Biron, P.; Lemoine, D. Water Balance, Transpiration and Canopy Conductance in Two Beech Stands. Agric. For. Meteorol. 2000, 100, 291–308. [Google Scholar] [CrossRef]
- Gessler, A.; Bächli, L.; Freund, E.R.; Treydte, K.; Schaub, M.; Haeni, M.; Weiler, M.; Seeger, S.; Marshall, J.; Hug, C.; et al. Drought Reduces Water Uptake in Beech from the Drying Topsoil, but No Compensatory Uptake Occurs from Deeper Soil Layers. New Phytol. 2022, 233, 194–206. [Google Scholar] [CrossRef]
- Zhao, N.; Meng, P.; He, Y.; Lou, Y.; Yu, X. Response of Water Sources in Platycladus orientalis and Vitex negundo var. Heterophylla System to Precipitation Events in Mountain Area of Beijing, China. Chin. J. Appl. Ecol. 2017, 28, 2155–2163. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Handle | Sample Tree | DBH (cm) | Sapwood Area (cm2) | Heartwood Diameter (cm) | Mean Sapwood Area (cm2) | Tree Height (m) | Average Tree Height (m) |
|---|---|---|---|---|---|---|---|
| NR | 1 | 12.69 | 175.95 | 1.45 | 197.27 | 16.94 | 16.08 |
| 2 | 13.19 | 190.81 | 1.78 | 16.46 | |||
| 3 | 14.27 | 225.06 | 1.93 | 14.85 | |||
| HR | 4 | 13.85 | 211.39 | 1.51 | 195.36 | 13.45 | 16.95 |
| 5 | 13.12 | 188.69 | 1.84 | 19.26 | |||
| 6 | 13.03 | 185.99 | 1.64 | 18.14 | |||
| AR | 7 | 12.97 | 184.19 | 1.58 | 194.48 | 14.22 | 15.36 |
| 8 | 14.01 | 216.55 | 2.03 | 16.31 | |||
| 9 | 12.92 | 182.71 | 1.85 | 15.56 | |||
| DR | 10 | 12.39 | 167.34 | 1.83 | 197.56 | 15.76 | 17.25 |
| 11 | 13.93 | 213.96 | 1.92 | 18.43 | |||
| 12 | 13.85 | 211.39 | 1.59 | 17.57 |
| Environmental Factor | Experimental Treatment | Fitting Equation | R2 | Pearson’s r | p | Significance |
|---|---|---|---|---|---|---|
| VPD | NR | y = 1.94 + 4.27x | 0.47232 | 0.687 ** | 0 | *** |
| HR | y = 4.95 + 5.72x | 0.52248 | 0.723 ** | 0 | *** | |
| AR | y = 6.18 + 11.60x | 0.74947 | 0.866 ** | 0 | *** | |
| DR | y = 7.51 + 7.29x | 0.17263 | 0.415 ** | 0.0014 | ** | |
| RS | NR | y = 0.11 + 0.02x | 0.45723 | 0.684 ** | 0.0639 | . |
| HR | y = 1.47 + 0.04x | 0.73527 | 0.860 ** | 0 | *** | |
| AR | y = 0.16x + 0.07x | 0.89308 | 0.946 ** | 0 | *** | |
| DR | y = 5.21x + 0.03x | 0.11333 | 0.361 ** | 0.9105 | ||
| WS | NR | y = 1.00 + 9.05x | 0.3438 | 0.5517 | 0.322 | |
| HR | y = 4.49 + 9.66x | 0.2141 | 0.4627 | 0.4298 | ||
| AR | y = 3.77 + 24.19x | 0.5663 | 0.6829 | 0.0088 | ** | |
| DR | y = 3.31 + 23.32x | 0.2534 | 0.5034 | 0.0192 | * |
| Precipitation Events | Environmental Factors | Treatment | Pre-Precipitation | Post-Precipitation | ||
|---|---|---|---|---|---|---|
| p | Significance | p | Significance | |||
| f | VPD | NR | 0.0285 | * | 0 | *** |
| HR | 0.0017 | ** | 0 | *** | ||
| AR | 0 | *** | 0 | *** | ||
| DR | 0 | *** | 0 | *** | ||
| RS | NR | 0.0653 | . | 0.0002 | *** | |
| HR | 0.0994 | . | 0.0003 | *** | ||
| AR | 0.7257 | 0.0066 | ** | |||
| DR | 0.2679 | 0.0089 | ** | |||
| WS | NR | 0.6408 | 0.0001 | *** | ||
| HR | 0.0108 | * | 0 | *** | ||
| AR | 0.7072 | 0.6837 | ||||
| DR | 0.5847 | 0.4773 | ||||
| c | VPD | NR | 0 | *** | 0 | *** |
| HR | 0 | *** | 0 | *** | ||
| AR | 0 | *** | 0 | *** | ||
| DR | 0 | *** | 0 | *** | ||
| RS | NR | 0.5364 | 0.0003 | *** | ||
| HR | 0.3708 | 0 | *** | |||
| AR | 0.8739 | 0.2284 | ||||
| DR | 0.5717 | 0.064 | . | |||
| WS | NR | 0.2109 | 0.0793 | . | ||
| HR | 0.103 | 0.0001 | *** | |||
| AR | 0.0007 | *** | 0.0002 | *** | ||
| DR | 0.016 | * | 0 | *** | ||
| g | VPD | NR | 0.0001 | *** | 0 | *** |
| HR | 0.0012 | ** | 0 | *** | ||
| AR | 0 | *** | 0 | *** | ||
| DR | 0 | *** | 0 | *** | ||
| RS | NR | 0.4836 | 0 | *** | ||
| HR | 0.0854 | . | 0 | *** | ||
| AR | 0.0153 | * | 0 | *** | ||
| DR | 0.0011 | ** | 0 | *** | ||
| WS | NR | 0.2001 | 0.104 | |||
| HR | 0.2577 | 0.2149 | ||||
| AR | 0.3921 | 0.1252 | ||||
| DR | 0.6064 | 0.5845 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Yu, X.; Ding, B.; Liu, Z.; Jia, G. Water Storage and Use by Platycladus orientalis under Different Rainfall Conditions in the Rocky Mountainous Area of Northern China. Forests 2022, 13, 1761. https://doi.org/10.3390/f13111761
Zhang X, Yu X, Ding B, Liu Z, Jia G. Water Storage and Use by Platycladus orientalis under Different Rainfall Conditions in the Rocky Mountainous Area of Northern China. Forests. 2022; 13(11):1761. https://doi.org/10.3390/f13111761
Chicago/Turabian StyleZhang, Xiao, Xinxiao Yu, Bingbing Ding, Zihe Liu, and Guodong Jia. 2022. "Water Storage and Use by Platycladus orientalis under Different Rainfall Conditions in the Rocky Mountainous Area of Northern China" Forests 13, no. 11: 1761. https://doi.org/10.3390/f13111761
APA StyleZhang, X., Yu, X., Ding, B., Liu, Z., & Jia, G. (2022). Water Storage and Use by Platycladus orientalis under Different Rainfall Conditions in the Rocky Mountainous Area of Northern China. Forests, 13(11), 1761. https://doi.org/10.3390/f13111761