
Stuck between the Mandibles of an Insect and of a Rodent: Where Does the Fate of Ash-Dominated Riparian Temperate Forests Lie?


Abstract
:1. Introduction
2. Materials and Methods
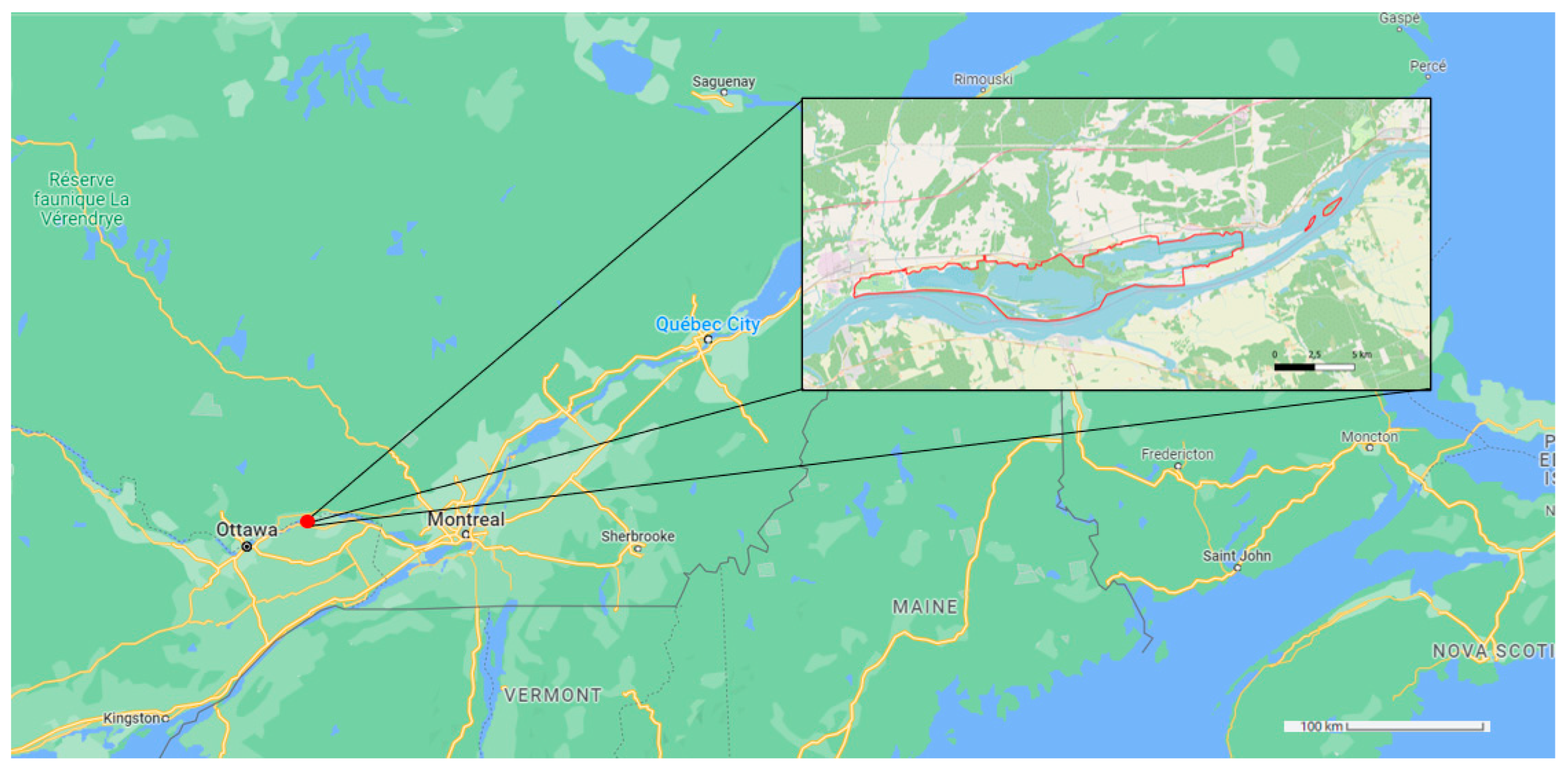
2.1. Study Site
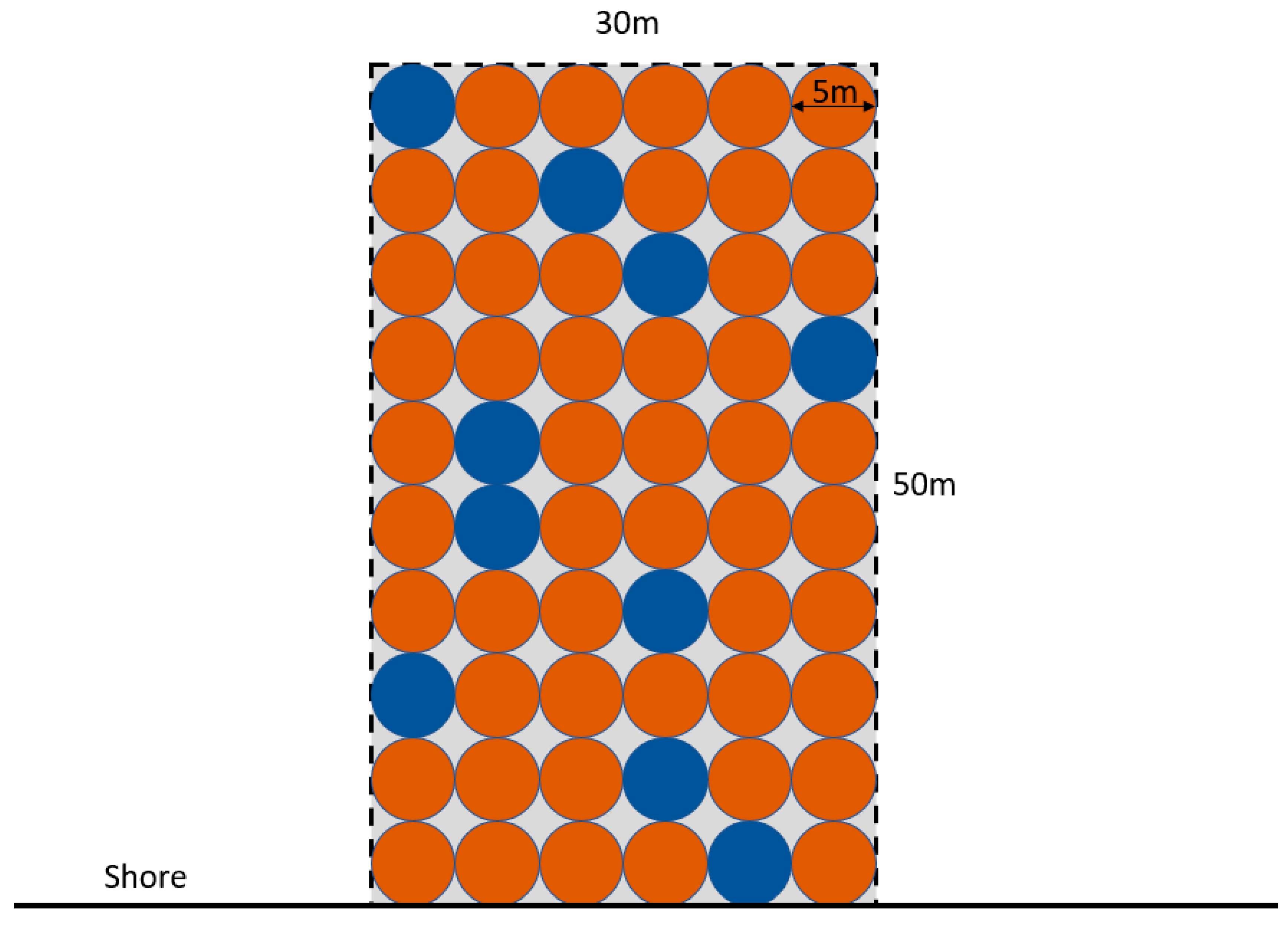
2.2. Experimental Design
2.3. Statistical Analysis
3. Results
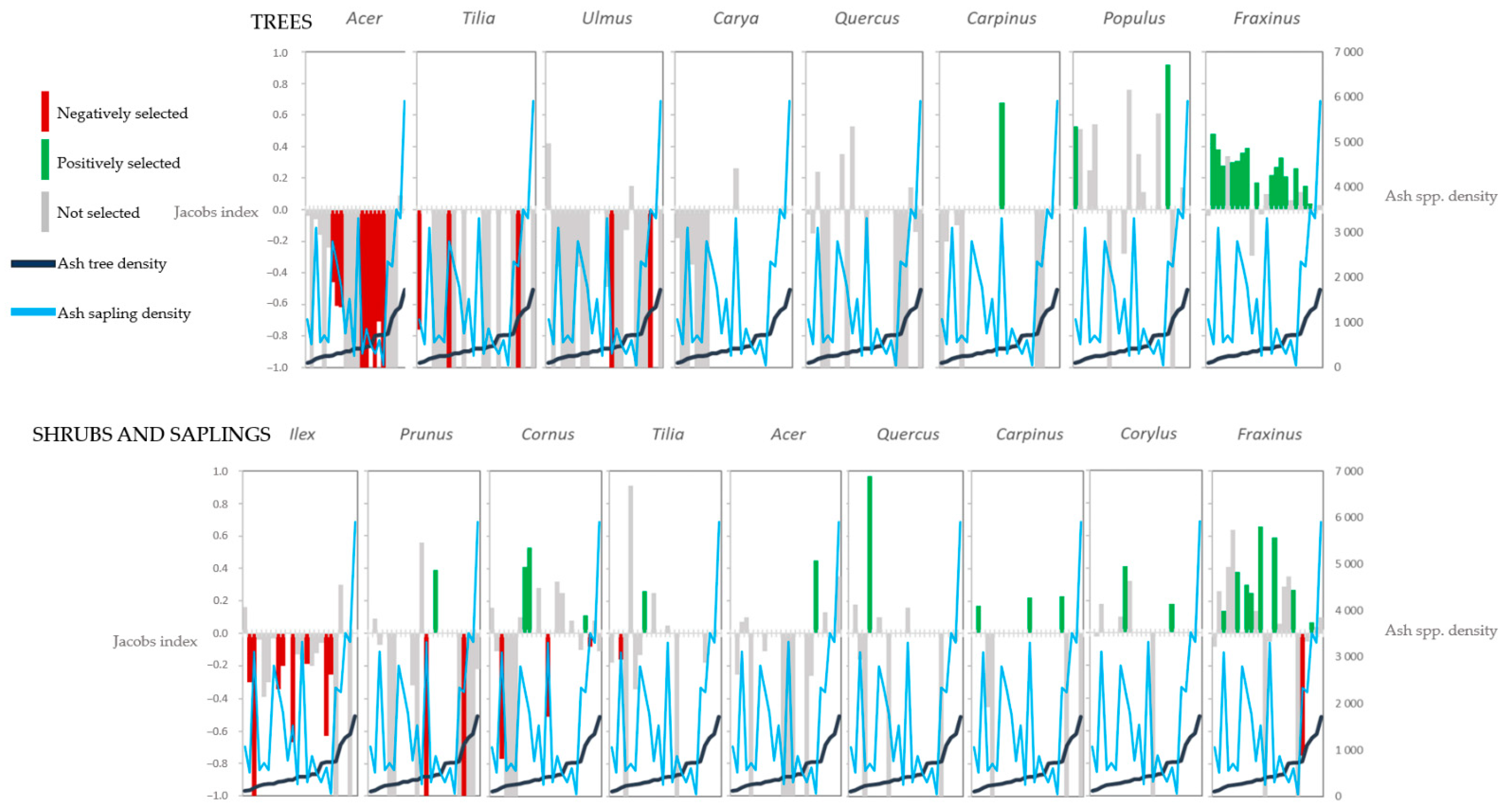
3.1. Selection for or against a Genus by Beavers (Objective 1)
3.2. Beaver Selection of Non-Ash Genera Relative to Ash spp. (Objective 2)
3.3. Effect of the Abundance of Ash spp. on the Chances of Other Genera Being Consumed by Beavers (Objective 3)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haack, R.A.; Eduard, J.; Houping, L.; Marchant Kenneth, R.; Petrice Toby, R.; Poland Therese, M.; Hui, Y. The Emerald Ash Borer: A New Exotic in North America. Mich. Entomol. Soc. 2002, 47, 1–5. [Google Scholar]
- Musolin, D.L.; Selikhovkin, A.V.; Peregudova, E.Y.; Popovichev, B.G.; Mandelshtam, M.Y.; Baranchikov, Y.N.; Vasaitis, R. North-Westward Expansion of the Invasive Range of Emerald Ash Borer, Agrilus Planipennis Fairmaire (Coleoptera: Buprestidae) towards the EU: From Moscow to Saint Petersburg. Forests 2021, 12, 502. [Google Scholar] [CrossRef]
- MacFarlane, D.W.; Meyer, S.P. Characteristics and Distribution of Potential Ash Tree Hosts for Emerald Ash Borer. For. Ecol. Manag. 2005, 213, 15–24. [Google Scholar] [CrossRef]
- Poland, T.M.; Chen, Y.; Koch, J.; Pureswaran, D. Review of the Emerald Ash Borer (Coleoptera: Buprestidae), Life History, Mating Behaviours, Host Plant Selection, and Host Resistance. Can. Entomol. 2015, 147, 252–262. [Google Scholar] [CrossRef] [Green Version]
- Knight, K.S.; Brown, J.P.; Long, R.P. Factors Affecting the Survival of Ash (Fraxinus spp.) Trees Infested by Emerald Ash Borer (Agrilus Planipennis). Biol. Invasions 2013, 15, 371–383. [Google Scholar] [CrossRef]
- Www.Fs.Fed.Us. Available online: https://www.fs.fed.us/nrs/atlas/tree (accessed on 14 June 2022).
- Www.Canada.Ca. Available online: https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/cosewic-assessments-status-reports/black-ash-2018.html (accessed on 14 June 2022).
- Engelken, P.J.; McCullough, D.G. Riparian Forest Conditions along Three Northern Michigan Rivers Following Emerald Ash Borer Invasion. Can. J. For. Res. 2020, 50, 800–810. [Google Scholar] [CrossRef]
- Ewel, K.C.; Cressa, C.; Kneib, R.T.; Lake, P.S.; Levin, L.A.; Palmer, M.A.; Snelgrove, P.; Wall, D.H. Managing Critical Transition Zones. Ecosystems 2001, 4, 452–460. [Google Scholar] [CrossRef]
- Hunter, M.L.; Acuña, V.; Bauer, D.M.; Bell, K.P.; Calhoun, A.J.K.; Felipe-Lucia, M.R.; Fitzsimons, J.A.; González, E.; Kinnison, M.; Lindenmayer, D.; et al. Conserving Small Natural Features with Large Ecological Roles: A Synthetic Overview. Biol. Conserv. 2017, 211, 88–95. [Google Scholar] [CrossRef]
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An Ecosystem Perspective of Riparian Zones. Bioscience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H. The Ecology Of Interfaces-Riparian Zones. Ann. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef] [Green Version]
- Paine, L.K.; Ribic, C.A. Comparison of Riparian Plant Communities under Four Land Management Systems in Southwestern Wisconsin. Ecosyst. Environ. 2002, 92, 93–105. [Google Scholar] [CrossRef]
- Maisonneuve, C.; Rioux, S. Importance of Riparian Habitats for Small Mammal and Herpetofaunal Communities in Agricultural Landscapes of Southern Québec. Ecosyst. Environ. 2001, 83, 165–175. [Google Scholar] [CrossRef]
- Forio, M.A.E.; de Troyer, N.; Lock, K.; Witing, F.; Baert, L.; de Saeyer, N.; Rîșnoveanu, G.; Popescu, C.; Burdon, F.J.; Kupilas, B.; et al. Small Patches of Riparian Woody Vegetation Enhance Biodiversity of Invertebrates. Water 2020, 12, 3070. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Herms, D.A. North American Arthropods at Risk Due to Widespread Fraxinus Mortality Caused by the Alien Emerald Ash Borer. Biol. Invasions 2010, 12, 1839–1846. [Google Scholar] [CrossRef]
- Grinde, A.R.; Youngquist, M.B.; Slesak, R.A.; Kolbe, S.R.; Bednar, J.D.; Palik, B.J.; D’Amato, A.W. Potential Impacts of Emerald Ash Borer and Adaptation Strategies on Wildlife Communities in Black Ash Wetlands. Ecol. Appl. 2022, 32, e2567. [Google Scholar] [CrossRef] [PubMed]
- Stoffyn-Egli, P.; Willison, J.H.M. Including Wildlife Habitat in the Definition of Riparian Areas: The Beaver (Castor Canadensis) as an Umbrella Species for Riparian Obligate Animals. Environ. Rev. 2011, 19, 479–493. [Google Scholar] [CrossRef]
- Nummi, P.; Liao, W.; Huet, O.; Scarpulla, E.; Sundell, J. The Beaver Facilitates Species Richness and Abundance of Terrestrial and Semi-Aquatic Mammals. Glob. Ecol. Conserv. 2019, 20, e00701. [Google Scholar] [CrossRef]
- Rosell, F.; Bozsér, O.; Collen, P.; Parker, H. Ecological Impact of Beavers Castor Fiber and Castor Canadensis and Their Ability to Modify Ecosystems. Mammal Rev. 2005, 35, 248–276. [Google Scholar] [CrossRef]
- Zavyalov, N.A. Beavers (Castor Fiber and Castor Canadensis), the Founders of Habitats and Phytophages. Biol. Bull. Rev. 2014, 4, 157–180. [Google Scholar] [CrossRef]
- Busher, P.E. Food Caching Behavior of Beavers (Castor Canadensis): Selection and Use of Woody Species. Am. Midl. Nat. 1996, 135, 343–348. [Google Scholar] [CrossRef]
- Vorel, A.; Válková, L.; Hamšíková, L.; Maloň, J.; Korbelová, J. Beaver Foraging Behaviour: Seasonal Foraging Specialization by a Choosy Generalist Herbivore. Behav. Ecol. Sociobiol. 2015, 69, 1221–1235. [Google Scholar] [CrossRef]
- Hall, J.G. Willow And Aspen In The Ecology Of Beaver On Sagehen Creek, California. Ecology 1960, 41, 484–494. [Google Scholar] [CrossRef]
- Smith, D.W.; Tyers, D.B. The History and Current Status and Distribution of Beavers in Yellowstone National Park. Northwest Sci. 2012, 86, 276–288. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.A.; Naiman, R.J. Browse Selection by Beaver: Effects on Riparian Forest Composition. Can. J. For. Res. 1990, 20, 1036–1043. [Google Scholar] [CrossRef]
- King, S.L.; Antrobus, T.J. Relationships between Gap Makers and Gap Fillers in an Arkansas Floodplain Forest. J. Veg. Sci. 2005, 16, 471–480. [Google Scholar] [CrossRef]
- Cook, J.E. Torrey Botanical Society Vegetation Changes in a Central Wisconsin Floodplain from Pre-Settlement to the Mid-21st Century. J. Torrey Bot. Soc. 2005, 132, 492–504. [Google Scholar] [CrossRef]
- Naiman, R.J.; Melillo, J.M.; Hobbie, J.E. Ecosystem Alteration of Boreal Forest Streams by Beaver (Castor Canadensis). Ecology 1986, 67, 1254–1269. [Google Scholar] [CrossRef]
- Donkor, N.T.; Fryxell, J.M. Lowland Boreal Forests Characterization in Algonquin Provincial Park Relative to Beaver (Castor Canadensis) Foraging and Edaphic Factors. Plant Ecol. 2000, 148, 1–12. [Google Scholar] [CrossRef]
- Martell, K.A.; Foote, A.L.; Cumming, S.G. Riparian Disturbance Due to Beavers (Castor Canadensis) in Alberta’s Boreal Mixedwood Forests: Implications for Forest Management. Ecoscience 2006, 13, 164–171. [Google Scholar] [CrossRef]
- Naiman, R.J.; Johnston, C.A.; Kelley, J.C. Alteration of North American Streams by Beaver. Bioscience 1988, 38, 753–762. [Google Scholar] [CrossRef]
- Müller-Schwarze, D.; Schulte, B.A.; Sun, L.; Müller-Schwarze, A.; Müller-Schwarze, C. Red maple (Acer Rubrum) Inhibits Feeding By Beaver (Castor Canadensis). J. Chem. Ecol. 1994, 20, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Raffel, T.R.; Smith, N.; Cortright, C.; Gatz, A.J. Central Place Foraging by Beavers (Castor Canadensis) in a Complex Lake Habitat. Am. Midl. Nat. 2009, 162, 62–73. [Google Scholar] [CrossRef]
- Rossell, R.C.J.; Arico, S.; Clarke, D.H.; Horton, J.L.; Rhode Ward, J.; Patch, S.C. Forage Selection of Native and Nonnative Woody Plants by Beaver in a Rare-Shrub Community in the Appalachian Mountains of North Carolina. Southeast. Nat. 2014, 13, 649–662. [Google Scholar] [CrossRef]
- Barnes, W.J.; Dibble, E. The Effects of Beaver in Riverbank Forest Succession. Can. J. Bot. 1988, 66, 40–44. [Google Scholar] [CrossRef]
- Lafond, R.; Leblanc, Y.; Pilon, C.; Société de la faune et des parcs du Québec, Direction du développement de la faune. Bilan Du Plan D’inventaire Aérien des Colonies de Castors Au Québec (1989–1994); Société de la Faune et des Parcs du Québec, Direction du Développement de la Faune, 2003; ISBN 2-550-40631-1. [Google Scholar]
- Major, M.; Gosselin, J.; Landry, Y.; Québec (Province). Direction des Inventaires Forestiers; Québec (Province). Ministère des Ressources Naturelles et de la Faune. Direction des Communications. Guide de Reconnaissance des Types Écologiques. Région Écologique 1a: Plaine Du Bas Outaouais et de l’archipel de Montréal; Ministère des Ressources Naturelles et de la Faune, Forêt Québec, Direction des Inventaires Forestiers: Québec City, QC, Canada, 2012; ISBN 9782551253036. [Google Scholar]
- Jacobs, J. Quantitative Measurement of Food Selection A Modification of the Forage Ratio and Ivlev’s Electivity Index. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, J.L.; Gorchov, D.L. Beavers Cut, but Do Not Prefer, an Invasive Shrub, Amur Honeysuckle (Lonicera Maackii). Biol. Invasions 2021, 23, 193–204. [Google Scholar] [CrossRef]
- Fryxell, J.M.; Doucet, C.M. Diet Choice and the Funcional Response of Beavers. Ecology 1993, 74, 1297–1306. [Google Scholar] [CrossRef]
- Wang, L.; Wang, D.; Liu, J.; Huang, Y.; Hodgkinson, K.C. Diet Selection Variation of a Large Herbivore in a Feeding Experiment with Increasing Species Numbers and Different Plant Functional Group Combinations. Acta Oecologica 2011, 37, 263–268. [Google Scholar] [CrossRef]
- Champagne, E.; Dumont, A.; Tremblay, J.P.; Côté, S.D. Forage Diversity, Type and Abundance Influence Winter Resource Selection by White-Tailed Deer. J. Veg. Sci. 2018, 29, 619–628. [Google Scholar] [CrossRef]
- Rand, T.A. Herbivore-Mediated Apparent Competition Between Two Salt Marsh Forbs. Ecology 2003, 84, 1517–1526. [Google Scholar] [CrossRef]
- Nisbet, D.; Kreutzweiser, D.; Sibley, P.; Scarr, T. Ecological Risks Posed by Emerald Ash Borer to Riparian Forest Habitats: A Review and Problem Formulation with Management Implications. For. Ecol. Manag. 2015, 358, 165–173. [Google Scholar] [CrossRef]
- Kashian, D.M.; Witter, J.A. Assessing the Potential for Ash Canopy Tree Replacement via Current Regeneration Following Emerald Ash Borer-Caused Mortality on Southeastern Michigan Landscapes. For. Ecol. Manag. 2011, 261, 480–488. [Google Scholar] [CrossRef]
- Aubin, I.; Cardou, F.; Ryall, K.; Kreutzweiser, D.; Scarr, T. Ash Regeneration Capacity after Emerald Borer (EAB) Outbreaks: Some Early Results. For. Chron. 2015, 91, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Fryxell, J.M.; Vamosi, S.M.; Walton, R.A.; Doucet, C.M. Retention Time and the Functional Response of Beavers. Oikos 1994, 71, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Durben, R.M.; Walker, F.M.; Holeski, L.; Keith, A.R.; Kovacs, Z.; Hurteau, S.R.; Lindroth, R.L.; Shuster, S.M.; Whitham, T.G. Beavers, Bugs and Chemistry: A Mammalian Herbivore Changes Chemistry Composition and Arthropod Communities in Foundation Tree Species. Forests 2021, 12, 877. [Google Scholar] [CrossRef]
- Kimball, B.A.; Perry, K.R. Manipulating Beaver (Castor Canadensis) Feeding Responses to Invasive Tamarisk (Tamarix spp.). J Chem. Ecol. 2008, 34, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.W. Dispersal Strategies and Cooperative Breeding in Beavers; University of Nevada: Reno, Nevada, 1997. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Variable | Odd-Ratio | Confidence Interval (95%) |
|---|---|---|
| Diameter | 0.95 | 0.94–0.97 |
| Distance to the shore | 0.21 | 0.18–0.25 |
| Acer | 0.07 | 0.05–0.11 |
| Ulmus | 0.13 | 0.07–0.25 |
| Quercus | 0.64 | 0.37–1.10 |
| Populus | 18.59 | 8.98–38.48 |
| Variable | Odd-Ratio | Confidence Interval (95%) |
|---|---|---|
| Diameter | 1.74 | 1.43–2.11 |
| Diameter squared | 0.83 | 0.77–0.89 |
| Distance to the shore | 0.32 | 0.28–0.36 |
| Ilex | 0.16 | 0.12–0.22 |
| Acer | 0.41 | 0.25–0.70 |
| Cornus | 0.43 | 0.32–0.59 |
| Tilia | 0.45 | 0.25–0.80 |
| Corylus | 0.53 | 0.31–0.91 |
| Prunus | 0.59 | 0.32–1.10 |
| Quercus | 0.79 | 0.29–2.17 |
| Carpinus | 1.85 | 1.27–2.68 |
| Genus | Acer | Quercus | Populus |
|---|---|---|---|
| Intercept | 0.01 [2.11 × 10−5–0.20] | 2009.20 [0.05–8.39 × 107] | 38.16 [0.03–4.87 × 104] |
| Diameter | 0.96 [0.93–0.99] | 0.95 [0.90–1.01] | 0.98 [0.94–1.02] |
| Distance to the shore | 0.29 [0.16–0.51] | 0.47 [0.30–0.75] | 0.43 [0.25–0.74] |
| Abundance of ash trees | 1.00 [0.99–1.01] | 0.98 [0.95–1.01] | 1.00 [0.98–1.02] |
| Abundance of ash saplings | 1.05 [1.03–1.07] | 1.03 [0.99–1.08] | 1.00 [0.94–1.06] |
| Shannon index for trees | 7.07 [1.06–47.19] | 0.08 [0.00–6.81] | 1.56 [0.11–21.52] |
| Shannon index for saplings | 1.62 [0.65–4.04] | 0.45 [0.04–5.21] | 0.44 [0.08–2.49] |
| Genus | Acer | Carpinus | Tilia | Ilex | Corylus |
|---|---|---|---|---|---|
| Intercept | 0.19 [0.01–31.84] | 1.69 [9.32 × 10−4–3052.54] | 1.02 [5.41 × 10−7–1.91 × 106] | 0.21 [8.65 × 10−4–53.11] | 422.36 [0.13–1.32 × 106] |
| Diameter | 1.15 [0.86–1.54] | 0.81 [0.65–1.01] | 1.49 [1.04–2.13] | Not tested | Not tested |
| Distance to the shore | 0.69 [0.48–0.99] | 0.77 [0.67–0.88] | 0.56 [0.34–0.93] | 0.62 [0.56–0.69] | 0.74 [0.65–0.84] |
| Abundance of ash trees | 0.99 [0.97–1.02] | 0.95 [0.91–0.99] | 0.99 [0.96–1.02] | 1.00 [0.98–1.02] | 0.98 [0.96–1.01] |
| Abundance of ash saplings | 1.03 [0.97–1.08] | 1.10 [1.01–1.20] | 1.08 [1.02–1.15] | 1.04 [0.99–1.08] | 1.03 [0.98–1.09] |
| Shannon index for trees | Not tested | Not tested | 3.10 [0.01–879.43] | 1.96 [0.12–32.16] | 0.29 [0.00–18.26] |
| Shannon index for saplings | 1.98 [0.14–28.03] | 1.32 [0.02–77.36] | 0.16 [0.00–5.38] | 0.72 [0.08–6.76] | 0.18 [0.01–4.07] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosner, S.; Dupuch, A.; Lorenzetti, F. Stuck between the Mandibles of an Insect and of a Rodent: Where Does the Fate of Ash-Dominated Riparian Temperate Forests Lie? Forests 2022, 13, 1760. https://doi.org/10.3390/f13111760
Rosner S, Dupuch A, Lorenzetti F. Stuck between the Mandibles of an Insect and of a Rodent: Where Does the Fate of Ash-Dominated Riparian Temperate Forests Lie? Forests. 2022; 13(11):1760. https://doi.org/10.3390/f13111760
Chicago/Turabian StyleRosner, Samuel, Angélique Dupuch, and François Lorenzetti. 2022. "Stuck between the Mandibles of an Insect and of a Rodent: Where Does the Fate of Ash-Dominated Riparian Temperate Forests Lie?" Forests 13, no. 11: 1760. https://doi.org/10.3390/f13111760
APA StyleRosner, S., Dupuch, A., & Lorenzetti, F. (2022). Stuck between the Mandibles of an Insect and of a Rodent: Where Does the Fate of Ash-Dominated Riparian Temperate Forests Lie? Forests, 13(11), 1760. https://doi.org/10.3390/f13111760