

Carbon Budget of Undrained and Drained Nutrient-Rich Organic Forest Soil


Abstract
:1. Introduction
- Foliar litter (LF);
- Ground vegetation (above- and below-ground biomass of herbs and grasses, GV);
- Fine roots of trees (FR);
- Moss and dwarf shrubs.
2. Materials and Methods
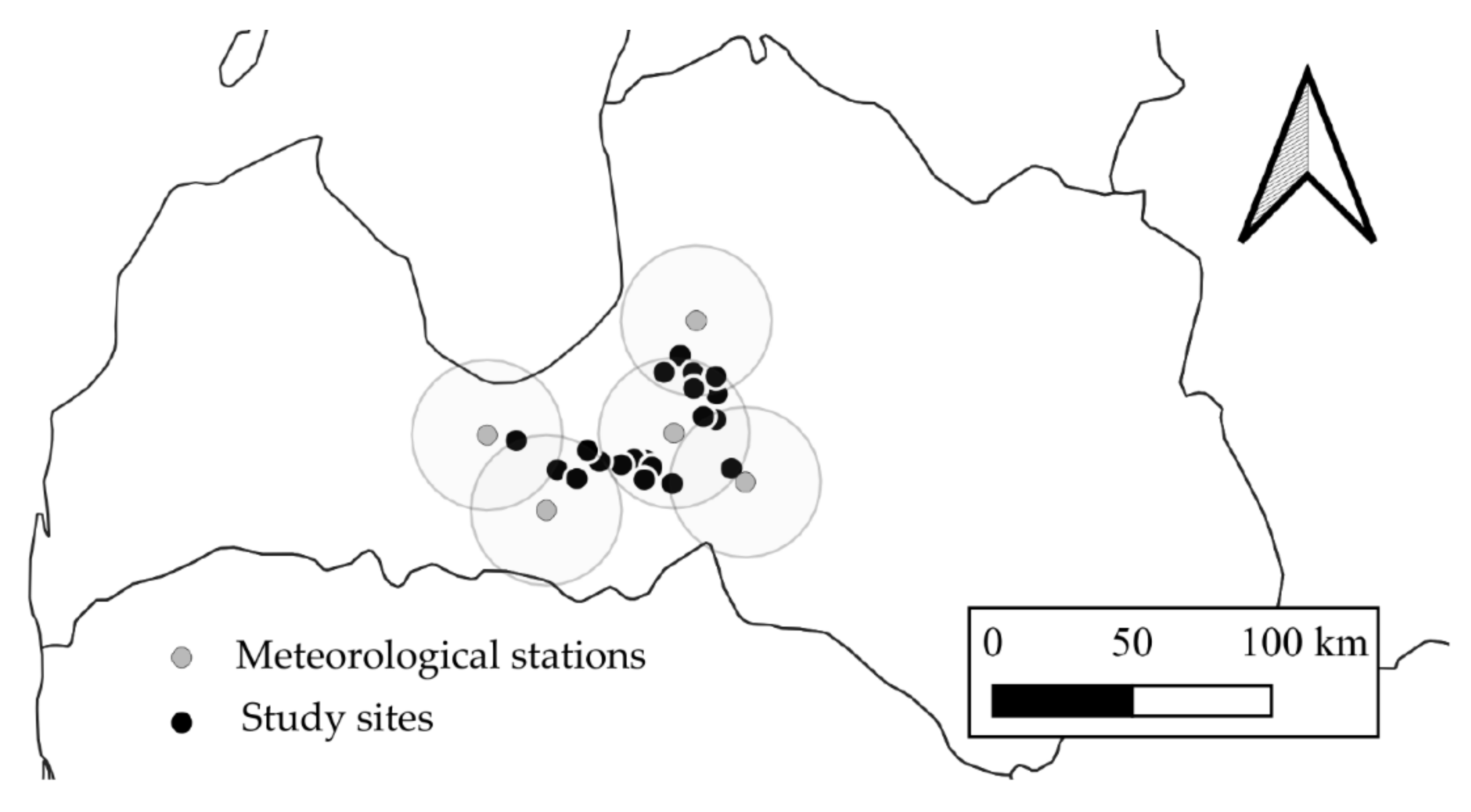
2.1. Study Site Description
2.2. Sampling and Laboratory Analysis of Soil and Soil Flux
2.3. Estimation of Soil Respiration
2.4. Estimation of Soil C Input by Litter
2.5. Estimation of Forest Soil Annual CSC
2.6. Statistical Analysis
3. Results
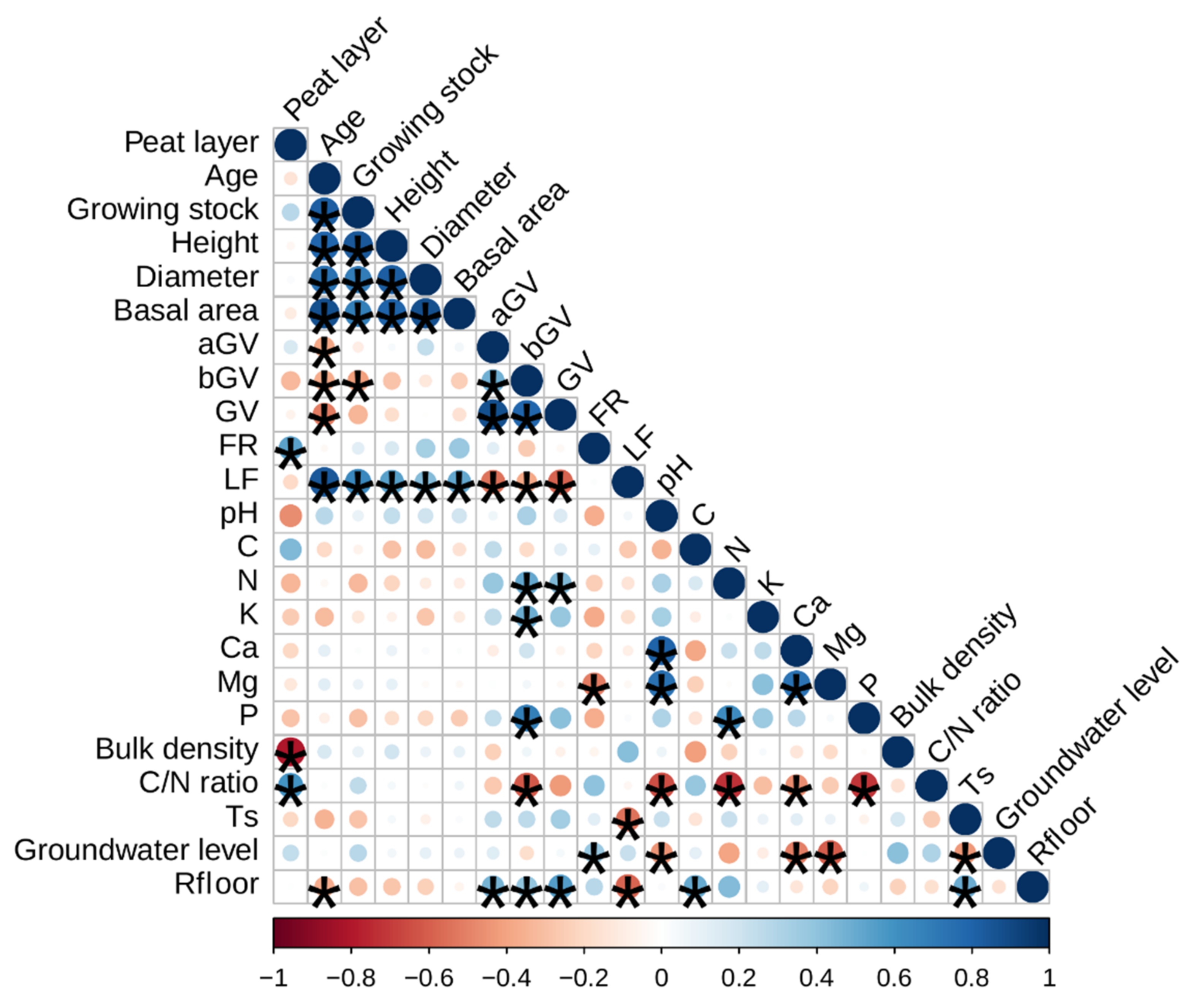
3.1. Soil Characteristics of the Study Sites
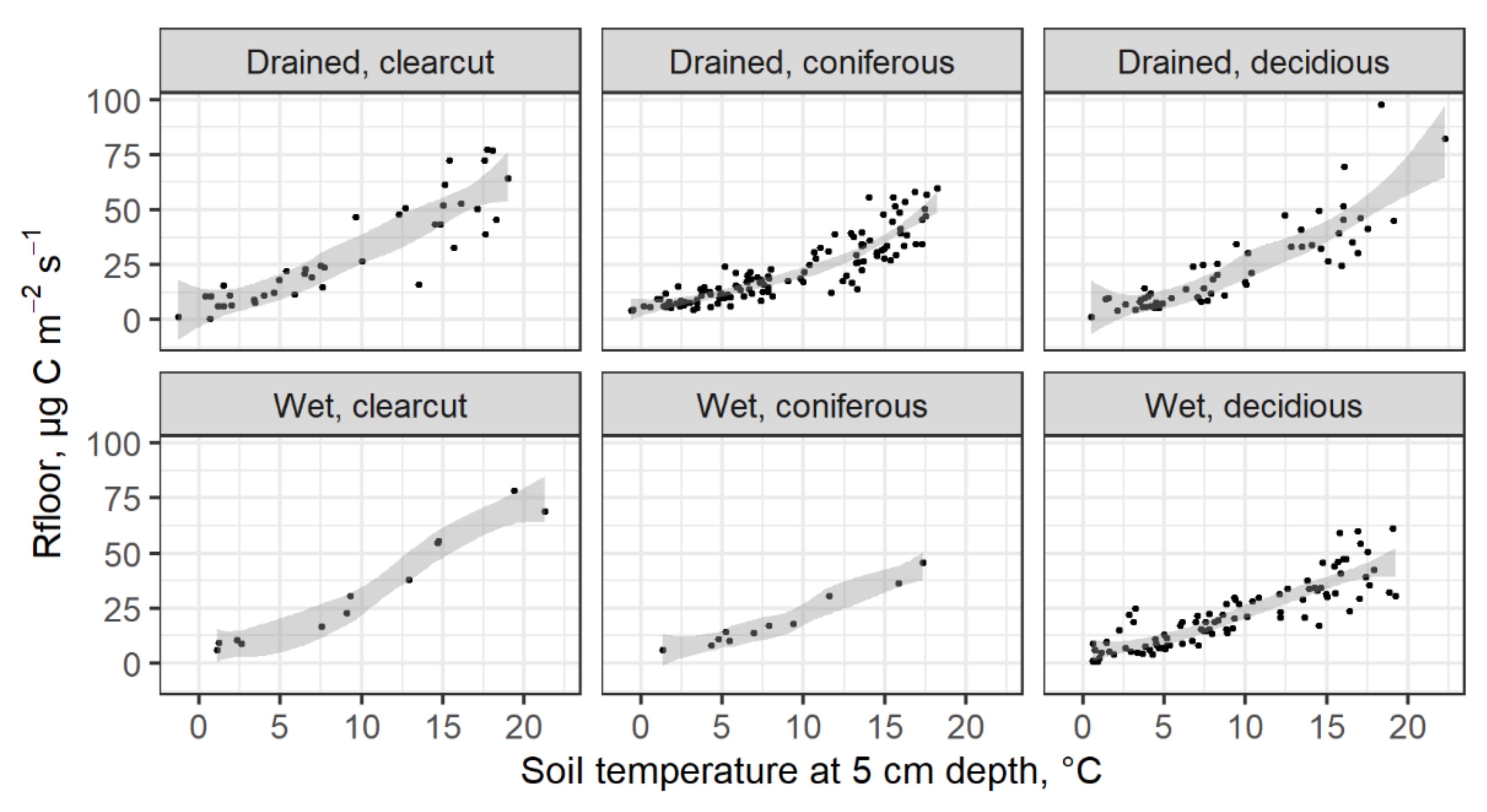
3.2. Soil Respiration
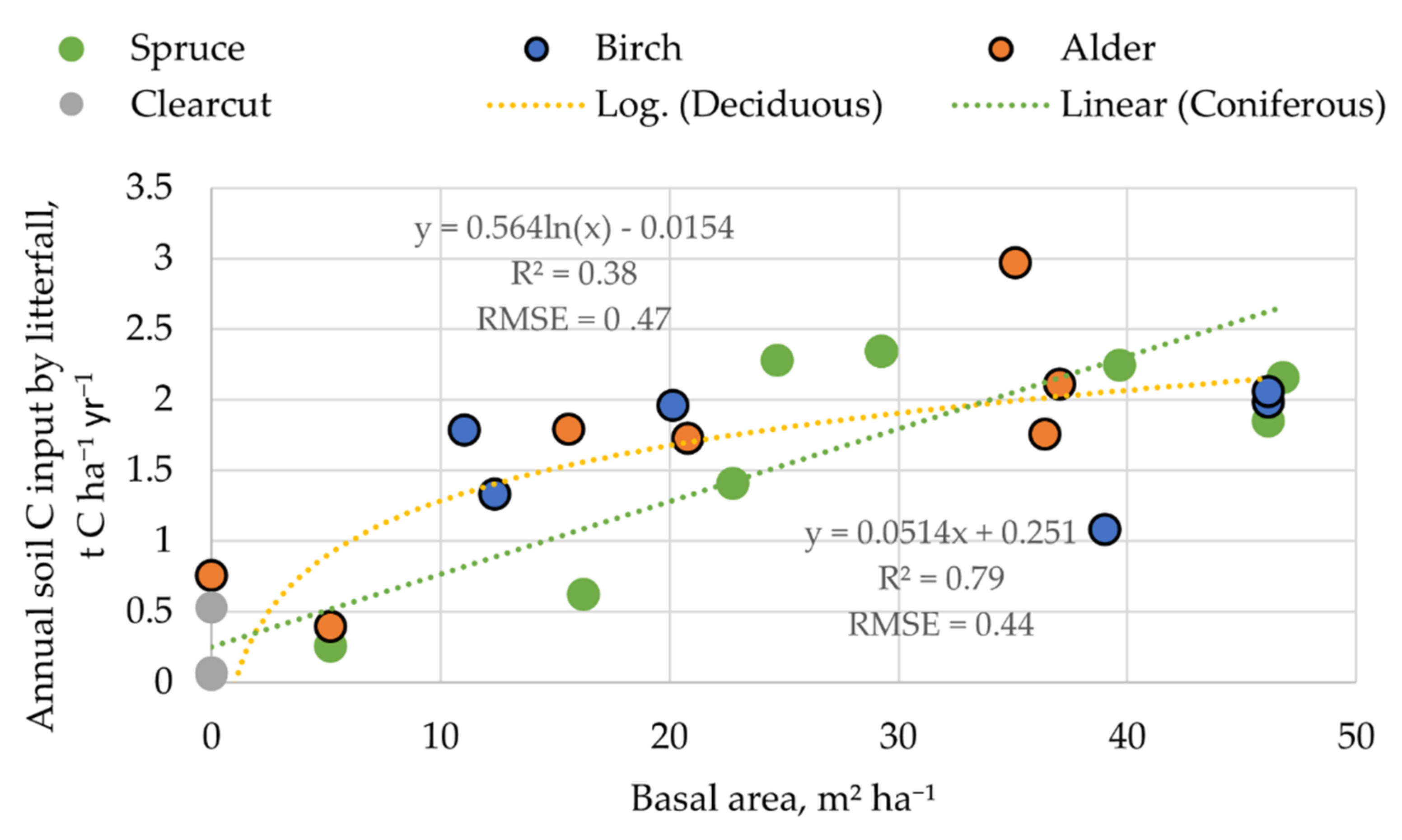
3.3. Soil C Input by Litter
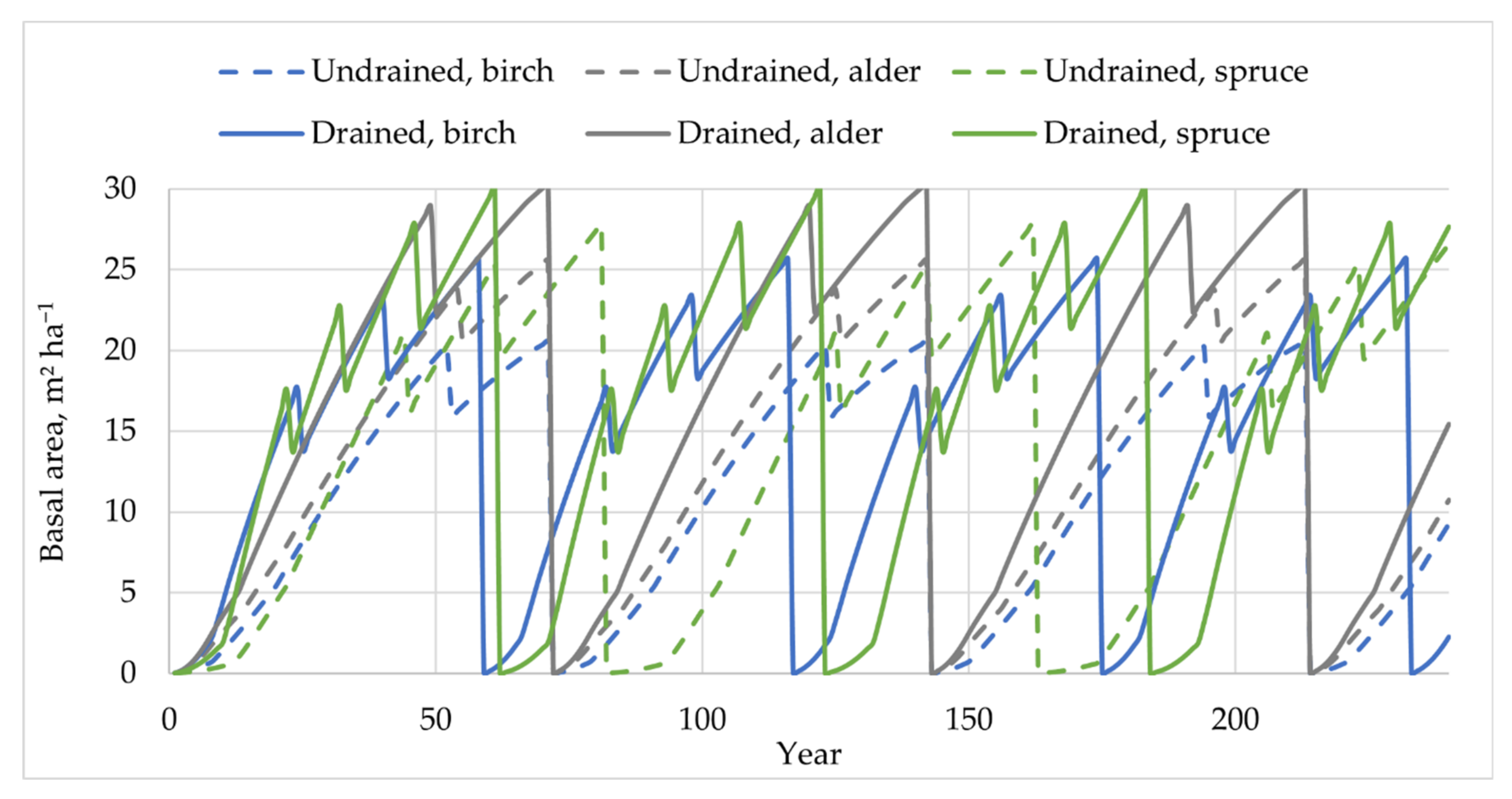
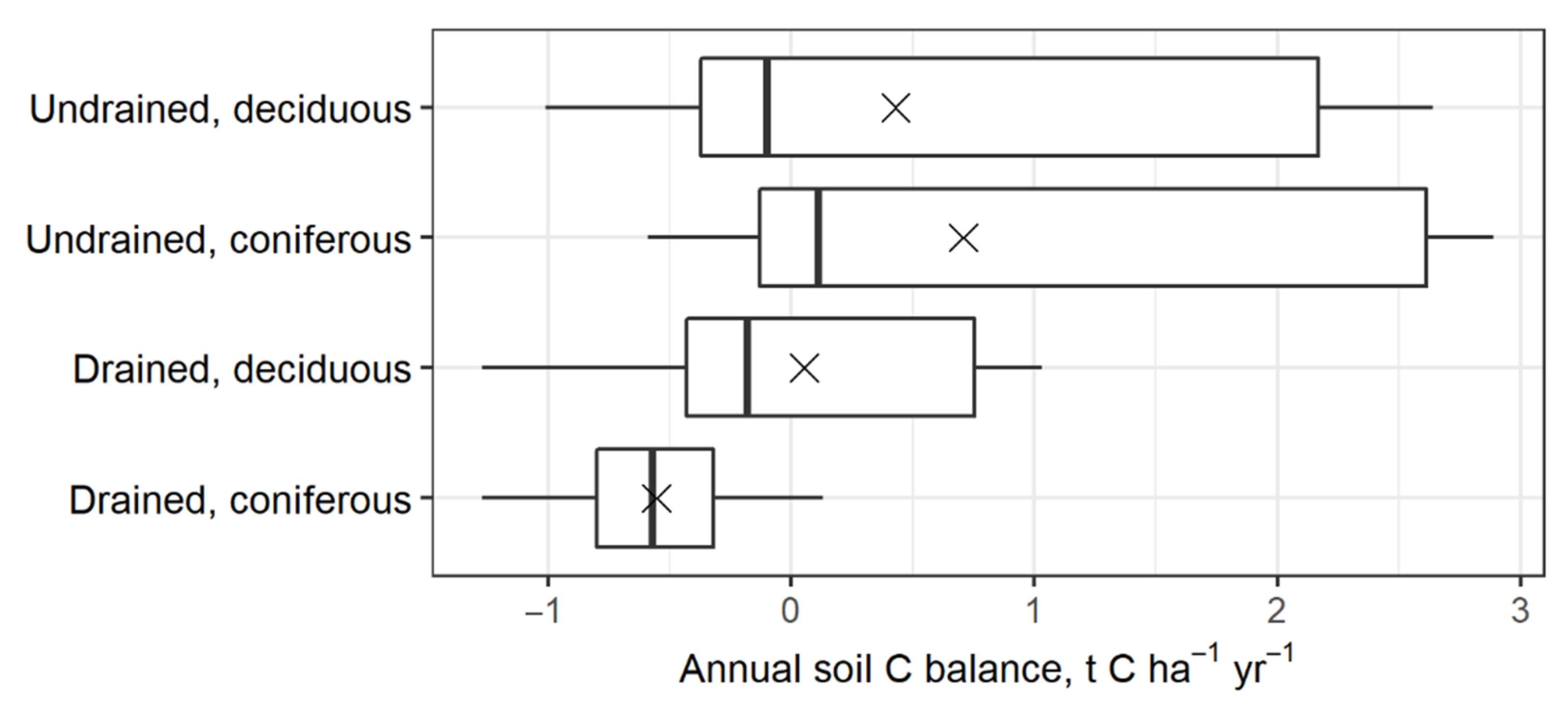
3.4. Summary of Estimated Annual Soil CSC
4. Discussion
4.1. Method of Rhet Calculation
4.2. Soil Respiration
4.3. Soil C Input by Litter
4.4. Annual Net Soil CSC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keith, H.; Lindenmayer, D.; MacKey, B.; Blair, D.; Carter, L.; McBurney, L.; Okada, S.; Konishi-Nagano, T. Managing Temperate Forests for Carbon Storage: Impacts of Logging versus Forest Protection on Carbon Stocks. Ecosphere 2014, 5, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Krug, J.H.A. Accounting of GHG Emissions and Removals from Forest Management: A Long Road from Kyoto to Paris. Carbon Balance Manag. 2018, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lundmark, T.; Bergh, J.; Nordin, A.; Fahlvik, N.; Poudel, B.C. Comparison of Carbon Balances between Continuous-Cover and Clear-Cut Forestry in Sweden. Ambio 2016, 45, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Pohjanmies, T.; Triviño, M.; Le Tortorec, E.; Mazziotta, A.; Snäll, T.; Mönkkönen, M. Impacts of Forestry on Boreal Forests: An Ecosystem Services Perspective. Ambio 2017, 46, 743–755. [Google Scholar] [CrossRef]
- Luyssaert, S.; Inglima, I.; Jung, M.; Richardson, A.D.; Reichstein, M.; Papale, D.; Piao, S.L.; Schulze, E.D.; Wingate, L.; Matteucci, G.; et al. CO2 Balance of Boreal, Temperate, and Tropical Forests Derived from a Global Database. Glob. Chang. Biol. 2007, 13, 2509–2537. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.P.; et al. Influence of Forest Management Activities on Soil Organic Carbon Stocks: A Knowledge Synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Kekkonen, H.; Ojanen, H.; Haakana, M.; Latukka, A.; Regina, K. Mapping of Cultivated Organic Soils for Targeting Greenhouse Gas Mitigation. Carbon Manag. 2019, 10, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Jauhiainen, J.; Alm, J.; Bjarnadottir, B.; Callesen, I.; Christiansen, J.R.; Clarke, N.; Dalsgaard, L.; He, H.; Jordan, S.; Kazanavičiūtė, V.; et al. Reviews and Syntheses: Greenhouse Gas Exchange Data from Drained Organic Forest Soils. A Review of Current Approaches and Recommendations for Future Research. Biogeosci. Discuss. 2019, 16, 4687–4703. [Google Scholar] [CrossRef] [Green Version]
- Paquel, K.; Bowyer, C.; Allen, B.; Nesbit, M.; Martineau, H.; Lesschen, J.P.; Arets, E. Analysis of LULUCF Actions in EU Member States as Reported under Art. 10 of the LULUCF Decision Final Study; Publications Office of the European Union: Brussels, Belgium, 2017. [Google Scholar]
- Lohila, A.; Minkkinen, K.; Aurela, M.; Tuovinen, J.P.; Penttilä, T.; Ojanen, P.; Laurila, T. Greenhouse Gas Flux Measurements in a Forestry-Drained Peatland Indicate a Large Carbon Sink. Biogeosciences 2011, 8, 3203–3218. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Tarvainen, L.; Nousratpour, A.; Björk, R.G.; Ernfors, M.; Grelle, A.; Kasimir Klemedtsson, Å.; Lindroth, A.; Räntfors, M.; Rütting, T.; et al. A Fertile Peatland Forest Does Not Constitute a Major Greenhouse Gas Sink. Biogeosciences 2013, 10, 7739–7758. [Google Scholar] [CrossRef]
- Uri, V.; Kukumägi, M.; Aosaar, J.; Varik, M.; Becker, H.; Morozov, G.; Karoles, K. Ecosystems Carbon Budgets of Differently Aged Downy Birch Stands Growing on Well-Drained Peatlands. For. Ecol. Manag. 2017, 399, 82–93. [Google Scholar] [CrossRef]
- Minkkinen, K.; Laine, J.; Shurpali, N.J.; Mäkiranta, P.; Alm, J.; Penttilä, T. Heterotrophic Soil Respiration in Forestry-Drained Peatlands. Boreal Environ. Res. 2007, 12, 115–126. [Google Scholar]
- Lupikis, A.; Lazdins, A. Soil Carbon Stock Changes in Transitional Mire Drained for Forestry in Latvia: A Case Study. Res. Rural Dev. 2017, 1, 55–61. [Google Scholar] [CrossRef]
- Bārdule, A.; Gerra-Inohosa, L.; Kļaviņš, I.; Kļaviņa, Z.; Bitenieks, K.; Butlers, A.; Lazdiņš, A.; Lībiete, Z. Variation in the Mercury Concentrations and Greenhouse Gas Emissions of Pristine and Managed Hemiboreal Peatlands. Land 2022, 11, 1414. [Google Scholar] [CrossRef]
- Ojanen, P.; Minkkinen, K.; Alm, J.; Penttilä, T. Soil-Atmosphere CO2, CH4 and N2O Fluxes in Boreal Forestry-Drained Peatlands. For. Ecol. Manag. 2010, 260, 411–421. [Google Scholar] [CrossRef]
- Skrebele, A.; Lupkina, L.; Cakars, I.; Siņics, L.; Lazdāne-Mihalko, J.; Puļķe, A.; Klavs, G.; Gračkova, L.; Lazdiņš, A.; Butlers, A.; et al. Latvia’s National Inventory Report—Submission under UNFCCC and the Kyoto Protocol; Ministry of Environmental Protection and Regional Development of the Republic of Latvia: Riga, Latvia, 2022.
- Bušs, K. Forest Ecology and Typology; Zinātne: Rīga, Latvija, 1981. [Google Scholar]
- Hutchinson, G.L.; Livingston, G.P. Use of Chamber Systems to Measure Trace Gas Fluxes. Agric. Ecosyst. Eff. Trace Gases Glob. Clim. Chang. 1993, 55, 63–78. [Google Scholar]
- Loftfield, N.; Flessa, H.; Augustin, J.; Beese, F. Automated Gas Chromatographic System for Rapid Analysis of the Atmospheric Trace Gases Methane, Carbon Dioxide, and Nitrous Oxide. J. Environ. Qual. 1997, 26, 560. [Google Scholar] [CrossRef]
- Cools, N.; De Vos, B. Sampling and Analysis of Soil, Manual Part X; ICP Forests: Hamburg, Germany, 2010; ISBN 9783865761620. [Google Scholar]
- Kriiska, K.; Frey, J.; Asi, E.; Kabral, N.; Uri, V.; Aosaar, J.; Varik, M.; Napa, Ü.; Apuhtin, V.; Timmusk, T.; et al. Variation in Annual Carbon Fluxes Affecting the SOC Pool in Hemiboreal Coniferous Forests in Estonia. For. Ecol. Manag. 2019, 433, 419–430. [Google Scholar] [CrossRef]
- Kukumägi, M.; Ostonen, I.; Uri, V.; Helmisaari, H.S.; Kanal, A.; Kull, O.; Lõhmus, K. Variation of Soil Respiration and Its Components in Hemiboreal Norway Spruce Stands of Different Ages. Plant Soil 2017, 414, 265–280. [Google Scholar] [CrossRef]
- Varik, M.; Kukumägi, M.; Aosaar, J.; Becker, H.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon Budgets in Fertile Silver Birch (Betula Pendula Roth) Chronosequence Stands. Ecol. Eng. 2015, 77, 284–296. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Wang, C.; Gower, S.T. A Global Relationship between the Heterotrophic and Autotrophic Components of Soil Respiration? Glob. Chang. Biol. 2004, 10, 1756–1766. [Google Scholar] [CrossRef]
- Sampling and Analysis of Litterfall. Available online: https://storage.ning.com/topology/rest/1.0/file/get/9995559487?profile=original (accessed on 15 August 2022).
- Bhuiyan, R.; Minkkinen, K.; Helmisaari, H.S.; Ojanen, P.; Penttilä, T.; Laiho, R. Estimating Fine-Root Production by Tree Species and Understorey Functional Groups in Two Contrasting Peatland Forests. Plant Soil 2017, 412, 299–316. [Google Scholar] [CrossRef]
- Laiho, R.; Bhuiyan, R.; Straková, P.; Mäkiranta, P.; Badorek, T.; Penttilä, T. Modified Ingrowth Core Method plus Infrared Calibration Models for Estimating Fine Root Production in Peatlands. Plant Soil 2014, 385, 311–327. [Google Scholar] [CrossRef]
- Lupiķis, A.; Lazdiņš, A. Soil Carbon Balance on Drained and Afforested Transitional Bog in Forest Research Station Vesetnieki in Latvia. In EGU General Assembly Conference Abstracts; EGU: Munich, Germany, 2015; Volume 17. [Google Scholar]
- Bardulis, A.; Lupiķis, A.; Stola, J. Carbon Balance in Forest Mineral Soils in Latvia Modelled with Yasso07 Soil Carbon Model. Res. Rural Dev. 2017, 1, 28–34. [Google Scholar] [CrossRef]
- Lazdiņš, A.; Petaja, G.; Kārkliņa, I. Verification of applicability of forest growth model AGM in elaboration of forestry projections for national forest reference level. In Proceedings of the 9th International Scientific Conference Rural Development 2019, Kaunas, Lithuania, 26–28 September 2019; Raupelienė, A., Ed.; Vytautas Magnus University: Kaunas, Lithuania, 2019. [Google Scholar] [CrossRef]
- Lazdiņš, A.; Lupiķis, A.; Butlers, A.; Bārdule, A.; Kārkliņa, I. Elaboration of Forest Reference Level for Latvia Dor Period between 2021 and 2025; Ministry of Agriculture of Republic of Latvia: Riga, Latvia, 2018.
- Muukkonen, P.; Mäkipää, R. Empirical Biomass Models of Understorey Vegetation in Boreal Forests According to Stand and Site Attributes. Boreal Environ. Res. 2006, 11, 355–369. [Google Scholar]
- FAO. Knowledge Reference for National Forest Assessments; FAO: Rome, Italy, 2015; ISBN 9789253088324. [Google Scholar]
- Mälkönen, E. Annual Primary Production and Nutrient Cycle in Some Scots Pine Stands. Commun. Inst. For. Fenn. 1974, 84, 1–87. [Google Scholar]
- Havas, P.; Kubin, E. Structure, Growth and Organic Matter Content in the Vegetation Cover of an Old Spruce Forest in Northern Finland. In Annales Botanici Fennici; Finnish Botanical Publishing Board: Helsinki, Finland, 1983; pp. 115–149. [Google Scholar]
- Palviainen, M.; Finér, L.; Mannerkoski, H.; Piirainen, S.; Starr, M. Responses of Ground Vegetation Species to Clear-Cutting in a Boreal Forest: Aboveground Biomass and Nutrient Contents during the First 7 Years. Ecol. Res. 2005, 20, 652–660. [Google Scholar] [CrossRef]
- Eggleston, H.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K. 2006 IPCC Guidelines for National Greenhouse Gas Inventories, Prepared by the National Greenhouse Gas Inventories Programme; IGES: Kanagawa, Japan, 2006. [Google Scholar]
- Butlers, A.; Bārdule, A.; Spalva, G.; Muižniek, E. N2O and CH4 emissions from naturally wet and drained nutrient-rich organic forest soils. In Proceedings of the 10th International Scientific Conference Rural Development; Vytautas Magnus University, Kaunas, Lithuania, 21–23 September 2021. [Google Scholar]
- Subke, J.A.; Inglima, I.; Cotrufo, M.F. Trends and Methodological Impacts in Soil CO2 Efflux Partitioning: A Metaanalytical Review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of Mechanisms and Quantification of Priming Effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Ngao, J.; Longdoz, B.; Granier, A.; Epron, D. Estimation of Autotrophic and Heterotrophic Components of Soil Respiration by Trenching Is Sensitive to Corrections for Root Decomposition and Changes in Soil Water Content. Plant Soil 2007, 301, 99–110. [Google Scholar] [CrossRef]
- Lohila, A.; Laurila, T.; Aro, L.; Aurela, M.; Tuovinen, J.P.; Laine, J.; Kolari, P.; Minkkinen, K. Carbon Dioxide Exchange above a 30-Year-Old Scots Pine Plantation Established on Organic-Soil Cropland. Boreal Environ. Res. 2007, 12, 141–157. [Google Scholar]
- Mäkiranta, P.; Hytönen, J.; Aro, L.; Maljanen, M.; Pihlatie, M.; Potila, H.; Shurpali, N.J.; Laine, J.; Lohila, A.; Martikainen, P.J.; et al. Soil Greenhouse Gas Emissions from Afforested Organic Soil Croplands and Cutaway Peatlands. Boreal Environ. Res. 2007, 12, 159–175. [Google Scholar]
- Yamulki, S.; Anderson, R.; Peace, A.; Morison, J.I.L. Soil CO2 CH4 and N2O Fluxes from an Afforested Lowland Raised Peatbog in Scotland: Implications for Drainage and Restoration. Biogeosciences 2013, 10, 1051–1065. [Google Scholar] [CrossRef] [Green Version]
- James, J.; Harrison, R. The Effect of Harvest on Forest Soil Carbon: A Meta-Analysis. Forests 2016, 7, 308. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How Strongly Can Forest Management Influence Soil Carbon Sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Ball, T.; Smith, K.A.; Moncrieff, J.B. Effect of Stand Age on Greenhouse Gas Fluxes from a Sitka Spruce [Picea Sitchensis (Bong.) Carr.] Chronosequence on a Peaty Gley Soil. Glob. Chang. Biol. 2007, 13, 2128–2142. [Google Scholar] [CrossRef]
- Klemedtsson, L.; Ernfors, M.; Björk, R.G.; Weslien, P.; Rütting, T.; Crill, P.; Sikström, U. Reduction of Greenhouse Gas Emissions by Wood Ash Application to a Picea Abies (L.) Karst. Forest on a Drained Organic Soil. Eur. J. Soil Sci. 2010, 61, 734–744. [Google Scholar] [CrossRef]
- Von Arnold, K.; Nilsson, M.; Hånell, B.; Weslien, P.; Klemedtsson, L. Fluxes of CO2, CH4 and N2O from Drained Organic Soils in Deciduous Forests. Soil Biol. Biochem. 2005, 37, 1059–1071. [Google Scholar] [CrossRef]
- Von Arnold, K.; Weslien, P.; Nilsson, M.; Svensson, B.H.; Klemedtsson, L. Fluxes of CO2, CH4 and N2O from Drained Coniferous Forests on Organic Soils. For. Ecol. Manag. 2005, 210, 239–254. [Google Scholar] [CrossRef]
- Ojanen, P.; Minkkinen, K.; Lohila, A.; Badorek, T.; Penttilä, T. Chamber Measured Soil Respiration: A Useful Tool for Estimating the Carbon Balance of Peatland Forest Soils? For. Ecol. Manag. 2012, 277, 132–140. [Google Scholar] [CrossRef]
- Silvola, J.; Alm, J.; Ahlholm, U.; Nykanen, H.; Martikainen, P.J. CO2 Fluxes from Peat in Boreal Mires under Varying Temperature and Moisture Conditions. J. Ecol. 1996, 84, 219–228. [Google Scholar] [CrossRef]
- Neumann, M.; Ukonmaanaho, L.; Johnson, J.; Benham, S.; Vesterdal, L.; Novotný, R.; Verstraeten, A.; Lundin, L.; Thimonier, A.; Michopoulos, P.; et al. Quantifying Carbon and Nutrient Input From Litterfall in European Forests Using Field Observations and Modeling. Glob. Biogeochem. Cycles 2018, 32, 784–798. [Google Scholar] [CrossRef]
- Bārdule, A.; Petaja, G.; Butlers, A.; Purviņa, D.; Lazdiņš, A. Estimation of Litter Input in Hemi-Boreal Forests with Drained Organic Soils for Improvement of GHG Inventories. Balt. For. 2021, 27, 2. [Google Scholar] [CrossRef]
- Neumann, M.; Godbold, D.L.; Hirano, Y.; Finér, L. Improving Models of Fine Root Carbon Stocks and Fluxes in European Forests. J. Ecol. 2020, 108, 496–514. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, A.; Palviainen, M.; Ojanen, P.; Kalliokoski, T.; Nöjd, P.; Kukkola, M.; Penttilä, T.; Mäkipää, R.; Leppälammi-Kujansuu, J.; Helmisaari, H.-S. Modelling Fine Root Biomass of Boreal Tree Stands Using Site and Stand Variables. For. Ecol. Manag. 2016, 359, 361–369. [Google Scholar] [CrossRef]
- Leppälammi-Kujansuu, J.; Aro, L.; Salemaa, M.; Hansson, K.; Kleja, D.B.; Helmisaari, H.-S. Fine Root Longevity and Carbon Input into Soil from Below- and Aboveground Litter in Climatically Contrasting Forests. For. Ecol. Manag. 2014, 326, 79–90. [Google Scholar] [CrossRef]
- Mäkelä, A.; Pulkkinen, M.; Mäkinen, H. Bridging Empirical and Carbon-Balance Based Forest Site Productivity—Significance of below-Ground Allocation. For. Ecol. Manag. 2016, 372, 64–77. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Derome, J.; Nojd, P.; Kukkola, M. Fine Root Biomass in Relation to Site and Stand Characteristics in Norway Spruce and Scots Pine Stands. Tree Physiol. 2007, 27, 1493–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Arnold, K.; Hånell, B.; Stendahl, J.; Klemedtsson, L. Greenhouse Gas Fluxes from Drained Organic Forestland in Sweden. Scand. J. For. Res. 2005, 20, 400–411. [Google Scholar] [CrossRef]
- Minkkinen, K.; Korhonen, R.; Savolainen, I.; Laine, J. Carbon Balance and Radiative Forcing of Finnish Peatlands 1900–2100—The Impact of Forestry Drainage. Glob. Chang. Biol. 2002, 8, 785–799. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Site Type | Ground Vegetation |
|---|---|
| Dryopterioso-caricosa | Thelypteris palustris Schott, Carex (L.), Iris pseudacorus (L.) Fuss, Scirpus (L.), Lysimachia vulgaris (L.), Cirsium oleraceum (L.) Scopoli, Filipendula ulmaria (L.) Maximowicz, Angelica sylvestris (L.) |
| Filipendulosa | Filipendula ulmaria (L.), Urtica dioica (L.), Geum rivale (L.), Paris quadrifolia (L.), Caltha palustris (L.), Solanum dulcamara (L.) |
| Oxalidosa turf. mel. | Cirsium oleraceum (L.) Scopoli, Hylocomium splendens (H.) Schimper, Rhytidiadelphus, Brachythecium, Vaccinium myrtillus (L.), Dryopteris filix-mas (L.) Schott |
| Parameter | Undrained Forest Sites | Drained Forest Sites | ||||||
|---|---|---|---|---|---|---|---|---|
| Spruce | Birch | Alder | Clearcut | Spruce | Birch | Alder | Clearcut | |
| Number of study sites | 1 | 3 | 5 | 1 | 12 | 3 | 2 | 4 |
| Age, years | 67 | 21–77 | 10–80 | 14–86 | 18–60 | 26–53 | ||
| Diameter, cm | 31 | 12–29 | 4–23 | 2–27 | 9–27 | 17–24 | ||
| Height, m | 28 | 12–28 | 4–29 | 2–24 | 9–22 | 17–26 | ||
| Basal area, m2 ha−1 | 61 | 17–71 | 8–57 | 8–72 | 19–60 | 32–56 | ||
| Growing stock, m3 ha−1 | 335 | 78–365 | 35–325 | 7–521 | 38–210 | 123–254 | ||
| Thickness of peat layer, cm | 68 | 31–52 | 30–99 | 47 | 37–99 | 25–75 | 60–70 | 63–99 |
| Parameter | Unit | Method Principle | Standard Method |
|---|---|---|---|
| Bulk density | kg m−3 | Gravimetry | LVS ISO 11272:2017 |
| Total C | g kg−1 | Elementary analysis (dry combustion) | LVS ISO 10694:2006 |
| Total N | g kg−1 | Elementary analysis (dry combustion) | LVS ISO 13878:1998 |
| Carbonate (CaCO3) | g kg−1 | Volumetry | LVS EN ISO 10693:2014 |
| pH | unit | Potentiometry | LVS ISO 10390:2021 |
| HNO3 extractable potassium (K), calcium (Ca), magnesium (Mg) and phosphorus (P) | g kg−1 | ICP-OES | LVS EN ISO 11885:2009 |
| Moisture Regime | Forest Site Characteristics | Characteristics of Measured Data (Range) | Model Coefficients | Model Characteristics | |||
|---|---|---|---|---|---|---|---|
| Ts, °C | Rfloor, µg C m−2 s−1 | a | b | RMSE | Q10 | ||
| Drained | Clear-cut | −1.3…19.0 | 0.5…77.2 | 5.784 | 0.141 | 11.9 | 4.10 |
| Deciduous | 0.5…22.3 | 1.2…97.8 | 4.476 | 0.143 | 10.3 | 4.18 | |
| Coniferous | −0.6…18.2 | 4.2…59.7 | 6.235 | 0.118 | 6.2 | 3.25 | |
| Undrained | Clear-cut | 1.1…21.3 | 6.3…78.5 | 7.298 | 0.124 | 10.6 | 3.46 |
| Deciduous | 0.7…19.3 | 0.6…61.3 | 4.700 | 0.136 | 8.5 | 3.90 | |
| Coniferous | 1.4…17.4 | 6.2…45.9 | 5.798 | 0.124 | 3.0 | 3.46 | |
| Moisture Regime | Forest Site Characteristics | Rfloor | Rhet | aGV | bGV | FR | LF | Net Balance |
|---|---|---|---|---|---|---|---|---|
| Drained | Clear-cut | −7.6 ± 2.2 | −4.3 ± 0.90 | 2.2 ± 0.3 | 1 ± 0.2 | 0.2 ± 0.2 | −0.9 ± 0.7 | |
| Deciduous | −6.2 ± 1.2 | −3.7 ± 0.53 | 0.7 ± 0.3 | 1.3 ± 0.3 | 0.3 ± 0.3 | 1.6 ± 0.3 | 0.2 ± 0.4 | |
| Coniferous | −6.3 ± 0.5 | −3.7 ± 0.23 | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.4 ± 0.1 | 1.7 ± 0.5 | 0.0 ± 0.3 | |
| Undrained | Clear-cut | −7.9 ± 1.2 | −4.4 ± 0.50 | 2.4 ± 0.3 | 1.4 ± 0.3 | 0.1 ± 0.4 | −0.4 ± 0.4 | |
| Deciduous | −6.1 ± 0.7 | −3.7 ± 0.32 | 0.7 ± 0.3 | 1.0 ± 0.4 | 0.3 ± 0.1 | 1.7 ± 0.6 | 0.0 ± 0.4 | |
| Coniferous | −5.1 ± 1.2 | −3.3 ± 0.50 | 0.5 ± 0.3 | 0.8 ± 0.3 | 0.5 ± 0.1 | 2.2 ± 0.4 | 0.8 ± 0.4 |
| Soil CSC Component | Drained | Undrained | ||
|---|---|---|---|---|
| Deciduous | Coniferous | Deciduous | Coniferous | |
| Rhet | −3.82 ± 0.45 | −3.85 ± 0.36 | −3.88 ± 0.7 | −3.54 ± 1.38 |
| LF | 1.14 ± 0.21 | 0.81 ± 0.24 | 1.12 ± 0.46 | 0.73 ± 0.22 |
| FR | 0.31 ± 0.25 | 0.44 ± 0.13 | 0.32 ± 0.29 | 0.53 ± 0.16 |
| aGV | 0.93 ± 0.61 | 1.1 ± 0.31 | 1.79 ± 0.61 | 1.59 ± 0.45 |
| Aboveground shrubs | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 |
| Aboveground mosses | 0.02 ± 0.02 | 0.09 ± 0.04 | 0.02 ± 0.02 | 0.11 ± 0.04 |
| bGV | 1.27 ± 0.42 | 0.59 ± 0.17 | 1.13 ± 0.52 | 1.00 ± 0.28 |
| Belowground shrubs, mosses | 0.07 ± 0.07 | 0.25 ± 0.1 | 0.07 ± 0.07 | 0.28 ± 0.12 |
| Net balance | −0.08 ± 0.03 | −0.55 ± −0.12 | 0.43 ± 0.17 | 0.71 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butlers, A.; Lazdiņš, A.; Kalēja, S.; Bārdule, A. Carbon Budget of Undrained and Drained Nutrient-Rich Organic Forest Soil. Forests 2022, 13, 1790. https://doi.org/10.3390/f13111790
Butlers A, Lazdiņš A, Kalēja S, Bārdule A. Carbon Budget of Undrained and Drained Nutrient-Rich Organic Forest Soil. Forests. 2022; 13(11):1790. https://doi.org/10.3390/f13111790
Chicago/Turabian StyleButlers, Aldis, Andis Lazdiņš, Santa Kalēja, and Arta Bārdule. 2022. "Carbon Budget of Undrained and Drained Nutrient-Rich Organic Forest Soil" Forests 13, no. 11: 1790. https://doi.org/10.3390/f13111790
APA StyleButlers, A., Lazdiņš, A., Kalēja, S., & Bārdule, A. (2022). Carbon Budget of Undrained and Drained Nutrient-Rich Organic Forest Soil. Forests, 13(11), 1790. https://doi.org/10.3390/f13111790