1. Introduction
Forest regeneration and establishment are critical stages during which considerable losses in species diversity can often occur [
1]. Natural regeneration (or natural regeneration in conjunction with artificial regeneration) often results in a large initial seedling bank. This is favored by a high proportion of unoccupied growing space and a diversity of microsite characteristics. As the plants increase in size and fill in the available space, competition becomes more intense and the growing conditions become more homogeneous. As the initial seedling bank progressively declines, species diversity may dramatically decrease as less competitive species are eliminated [
2]. These competitive relationships must be considered in silvicultural techniques [
3,
4,
5]. When the transformation of pure Norway spruce (
Picea abies (L.) Karst.) stands is adopted, natural regeneration of Norway spruce (combined with planting of reintroduced tree species) must be controlled to prevent reintroduced species from being overtopped and out-competed [
6]. One of the main silvicultural tools for this purpose is adjusting light intensities through overstory canopy openness. Thus, forest structure and heterogeneity are encouraged [
7,
8].
Uneven-aged forest management is characterized by silvicultural practices that create or maintain stands made up of trees of different age classes [
9,
10]. Different terms and definitions are used for the numerous forms of uneven-aged silviculture [
10,
11]; however, all these silvicultural systems are based on the creation of canopy openings or gaps to allow light to reach the floor of the stand [
5,
7,
8]. The degree of growth heterogeneity, and structure of the understory induced by the variability of light levels, can be as variable and advanced as the shade tolerance of particular tree species is high [
12,
13]. In similar studies of forest ecology where light is an independent variable, it is important to consider reference levels of other site conditions, such as nutrients and water supply. This is because shade tolerance is shifted by the availability or scarcity of other site factors [
14,
15,
16]. It is therefore advantageous to perform such an investigation on nutrient-poor sites as a basic level of nutrient resources in combination with average water supply, which can then be used as a reference compared to more optimal conditions (such as nutrient-enriched conditions) where shade tolerance is generally increased [
14,
15].
Tree species with different levels of shade tolerance show different levels of both morphological and physiological plasticity under various light conditions [
4,
8,
17]. The response of crown morphology and height growth depending on shade tolerance or intolerance has been described in different studies [
3,
18,
19]. In general, the conclusion of those studies is that shade-tolerant tree species can adapt their crown morphology more acutely and in more morphological traits (such as crown length, width, and projection or terminal length) along the light gradient compared to shade-intolerant tree species. Shade-tolerant tree species can effectively reduce their growth and survive under low light intensities due to their diminished growth rate and photo-morphological responses resulting in lower mortality rate in shaded conditions; after their release, they react immediately to increased light intensities with an increased growth rate [
15,
20,
21]. However, they do not reach the same growth rate under canopy-open conditions as shade-intolerant tree species [
22].
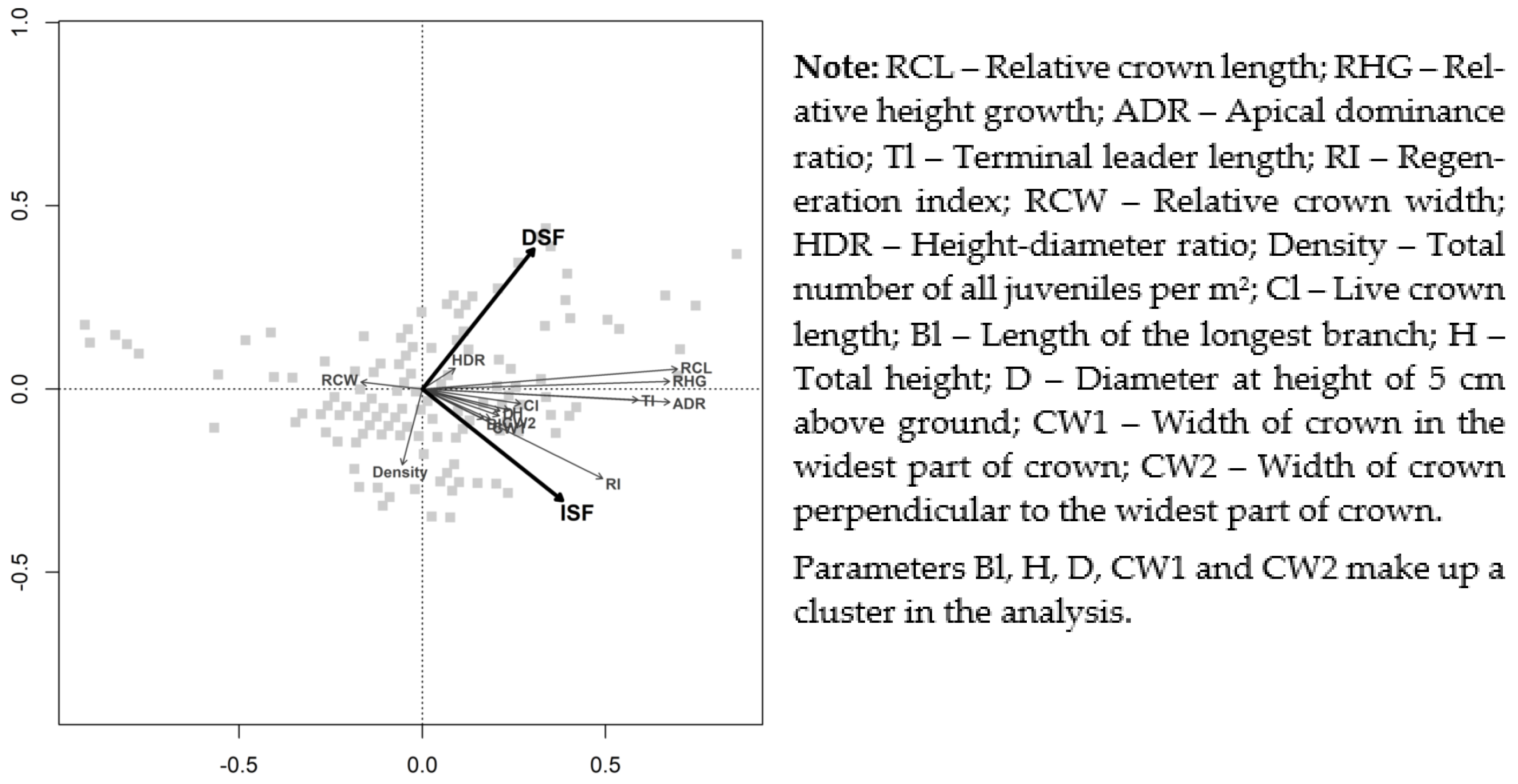
The three following objects of interest related to photo-morphological patterns of Norway spruce juveniles were analyzed in this paper:
The existence and intensity of morphological plasticity (i.e., the ability to respond to the different availability of resources by acclimation of the morphological traits) as a response to different light intensities;
The definition of microsites where the morphological plasticity was realized most distinctly;
The influence of light intensities on the general development of Norway spruce juveniles (including, for instance, their density) and a consideration of all observed patterns for further use in silvicultural guidelines.
4. Discussion
Other authors have established the same positive correlation between light conditions and height growth of Norway spruce natural regeneration [
19,
45]. The lower correlation of total height (H) compared to the terminal length (Tl) can be explained in that the response of height growth to particular light conditions is not immediate. On the contrary, it takes time from when the height growth is again in full compliance with light conditions, as has been established by Nilson and Lundqvist (2001) [
46], Robakowski et al. (2004) [
41], and Chrimes and Nilson (2005) [
19]. This is in compliance with the results of Diaci and Firm (2011) [
3], who demonstrated a significant correlation between Tl and light intensity both for Norway spruce and silver fir (
Abies alba Mill.) seedlings, but did not find a significant correlation with H. Tl has a higher probability of being adaptive to current light conditions, while H reflects the history of height growth through changing light conditions. Although light conditions were reasonably stable for a long time (at least for 15 years or more) in this study, some changes did occur (due to overstory growth and overstory canopy development). A similar situation can be concluded for the index RHG, as RHG plasticity was more carried out through Tl changes than H changes. The parameter of Tl was also proven by Jaloviar et al. (2013) [
16] as being associated with light intensities both for Norway spruce natural regeneration and silver fir underplantings. By contrast, they found no significant effect of light on the RHG of either tree species [
16].
A high level of photo-morphological plasticity in this study was found for lateral crown growth. However, within the evaluation involving height classes, none of those adaptations (at the level of parameters Bl, CW1, and CW2 or indexes RCW, CP, and ISD) were employed in the smallest height class (10–50 cm). Similar results were established by Duchesneau et al. (2001) [
37] for balsam fir (
Abies balsamea (L.) Mill.) when they found a generally higher level of correlations of morphological plasticity and light for the height class of 100–200 cm compared to the height class of 50–100 cm. Similar findings were confirmed for other conifer shade-tolerant tree species [
28,
44]. However, a continual photo-morphological acclimation through a gradient of the dimension of natural regeneration can be limited, because as Wright et al. (1998) [
15] found, a height of 6 m can be a threshold for many different tree species when the described trend starts to weaken or even disappears.
It was widely established that the plasticity of crown morphology plays a key role in the adaptation ability to survive under a wide range of light intensities, and thus this feature is uniquely distinctive for shade-tolerant tree species [
12,
13,
47,
48]. Crown morphology has a crucial effect on light use efficiency and thus on the competitive strength of an individual [
44,
48,
49,
50]. This competitiveness within natural regeneration can increase if light availability becomes limiting due to overstory shelter and can vary according to the ratio of diffuse and direct light in understory microsites [
8]. Consequently, shade-tolerant tree species have the ability to adapt their growth just through acclimations via photo-morphological plasticity of the crown, shoots, and needles or leaves [
51,
52]. Thus, shade-tolerant conifer tree species crown morphology varies from conical under high light conditions to umbrella-shaped under low light conditions [
18,
53,
54]. This morphological development consists of acclimations via a reduction in height growth and crown length [
29,
55] and initiation of crown lateral growth to maximize carbon gain under shade [
41,
54,
56]. In addition, the described pattern is aimed at reducing self-shading within the crown [
18,
55,
57]. As light intensities increase, height increment is initiated, making the light more available for the lowest whorls, which initiates the establishment of a continual conical-shaped crown [
58]. Acclimation ability (expressed as different increment allocation patterns resulting in different architecture) varies between tree species and plays a key role in photosynthetic efficiency (essential under low light conditions) and thus offers a competitive advantage to particular tree species [
22,
28,
49,
52].
Within the 10–50 cm height class, juveniles invest in vertical growth (proven significant correlation of Tl and light) to reach the best social position through height increment with no lateral crown growth having yet taken place. However, to optimize increment allocation (i.e., carbon gain) and to avoid self-shading, the ability to reduce crown length is already fully developed even in this early stage of natural regeneration (broadly proven by significant correlations of Cl, RCL, and CWLR). As Takahashi (1996) [
43] stated, the whole process is a balance between investment into height growth to reach future profit and investment into surviving when the current total height is preserved, as is broadly shown in other studies [
18,
22,
29,
41]. Although the index CWLR integrates both lateral crown growth and crown length, it significantly correlates with light. This takes place through morphological acclimation of crown length and can be concluded from the established plasticity of both Cl and RCL in the smallest height class when no proof of morphological plasticity for lateral crown growth was found. The two described patterns of photo-morphological acclimations (height growth and crown length) were also present in juveniles taller than 51 cm when the ability to employ photo-morphological acclimations to crown lateral growth was supplemented.
As a consequence, other studies considered both ADR and RCL as indexes for expressing the level of shade tolerance, and deducing the light intensities under which juvenile shade-tolerant trees have been growing [
29,
41].
The crown length is considered the essential photo-morphological characteristic for distinguishing between shade tolerance and semi-tolerance, since an absence of this acclimation for shade semi-tolerant tree species, such as eastern white pine (
Pinus strobus L.), has been shown [
27,
44,
55].
Our finding of the correlation between HDR and light verified the classification of Norway spruce as a shade-tolerant tree species because this level of plasticity is also considered a typical ability of shade-tolerant tree species [
59] and is absent in shade semi-tolerant tree species [
27,
44]. Although we did not prove a significant correlation between HDR and light conditions in the general evaluation, it was established for juveniles taller than 51 cm when evaluated according to height classes.
Thus, only the initial stage of natural regeneration did not exhibit acclimation of HDR, whereas a correlation of HDR with all observed light variables for the highest stratum of natural regeneration (height class >151 cm) was established. The observed plasticity of HDR was created through acclimations of height growth when stem thickness was not related to light intensities. No correlation between D and light can be explained by intense intraspecific competition within natural regeneration. To some extent, the increment is invested into height growth to reach a favorable position in relation to neighboring trees. To a greater extent, it is invested into adaptations of the crown to make photosynthesis under shade as effective as possible to reach a high probability of survival [
18,
43,
58] while investment into stem radial growth is reduced and affected by the density of juveniles. In such conditions of dense natural regeneration, the intraspecific competition was a critical factor when compared to the results of Coates and Burton (1999) [
60], who established the significant influence of light on D for five different conifer tree species five years after planting. It is clear that in young planted seedlings, competition was substantially lower compared to natural regeneration in our study. Furthermore, the crucial influence of density on D was also described by Chrimes and Nilson (2005) [
19], as well as Petritan et al. (2012) [
61], who established the significance of this finding for conifers rather than broadleaf species. Thus, the correlation of stem radial increment and light intensities is fully revoked when intraspecific (or interspecific) competition appears; only the pattern of height growth and its dependency on light remains. Consequently, HDR plasticity, fully arranged through height growth plasticity, occurs. This also confirmed the finding of the independence of D on light intensities in the natural regeneration of other conifers, as Kučeravá et al. (2012) [
62] found the same for silver fir and Claveau et al. (2002) [
44] observed the same for four conifer species from a group of five. On the other hand, in this study, only D was tested when there were no available data about radial increment. Radial increment can be more sensitive to certain light conditions, as was established by Duchesneau et al. (2001) [
37] for balsam fir or Greis and Kellomäki (1981) [
63] for Norway spruce.
This study established only a weak relationship between regeneration density and light. Lunquist and Fridman (1996) [
36] and Hasenauer and Kindermann (2002) [
64] also did not find a correlation between Norway spruce regeneration density and the basal area of the overstory. Similarly, Szewczyk and Szwagrzyk (2010) [
65] did not find a correlation for Norway spruce either, while for silver fir, they found a significant correlation. Jaduď et al. (2014) [
66], who evaluated the influence of light intensity on the density of seedlings after directseeding (using the same amount of seeds within various treatments), found no correlation between light intensities and density for Norway spruce or silver fir. We established that there was a fragile relationship for natural regeneration up to a maximum of 1 m in height, which corresponds well with the findings of Roženbergar et al. (2007) [
67]. They did not find a significant correlation between light and the density of silver fir and observed a continual decrease in its density with its increasing height until a height of 110 cm when density developed asymptotically to axis x. Thus, the height of approximately 1 m can work as a general threshold when the (intraspecific) competition (resulting to autoreduction) starts to dominate, and the influence of light (which slightly affects the density of juveniles up to 1 m and is reflected in height growth as well) is retracted. By contrast, the regeneration index (RI) where dimension was considered was significantly correlated with light in this study, which was confirmed by Grassi et al. (2004) [
68] both for Norway spruce and silver fir. The importance of dimension (height) on the development of density has been clearly demonstrated. Furthermore, the importance of microclimates in microsites is an important factor, as shown by Kupferschmid and Bugmann (2005) [
69], Hunziker and Brang (2005) [
70], and Kathke and Bruelheide (2010) [
71]. More studies focused on the complex mechanisms of various factors affecting Norway spruce natural regeneration density should be conducted in the future.
5. Conclusions
The results of our research indicate that Norway spruce juveniles have immense photo-morphological plasticity in lateral crown growth, crown length, and height growth. The ranges of plasticity (expressed by the maximal and minimal values of RHG and ADR) for height growth and crown lateral growth acclimated to light conditions to a similar extent. However, the plasticity of lateral crown growth was absent in the smallest height class (10–50 cm). The capability for acclimations in crown length was established robustly throughout the whole range of natural regeneration dimensions. HDR showed a significant positive correlation with light intensities for juveniles from 51 cm in height. This shows the positive impact of overstory shelter on the mechanical stability of juveniles (making the HDR value smaller due to the shade of the overstory shelter) and is a representative example of nature’s automation (also known as biological automation) principles in tending in young forest stands. However, this means that when the overstory canopy is intensively opened (or even removed) by silvicultural intervention or by disturbance, the risk of mechanical lability of Norway spruce understory will occur if no further tending is applied in this natural regeneration.
The existence and intensity of photo-morphological acclimations firmly established Norway spruce as a shade-tolerant species. Thus, light conditions in the forest stand understory (which can be easily influenced by silvicultural techniques aimed at the forest stand overstory) can effectively encourage forest stand structural heterogeneity as Norway spruce juveniles follow light conditions with their growth. Moreover, through the use of appropriate silviculture techniques, it is possible to take full control of the growth vigor of Norway spruce juveniles. This can be effectively used for the transformation of Norway spruce monocultures to uneven-aged mixed stands, once it is deemed appropriate for Norway spruce to be one of the species in the future species composition of the forest stand. For such silviculture practice, two main growth regimes can be generally distinguished: suppressed and enhanced. These regimes are not distinguished by distinct thresholds but can be found at the level of ca. 0.2 TSF. From this intensity, both RHG and ADR progressively increase, whereas until ca. 0.15 TSF they very slightly increase with light (RCL increases constantly with TSF within the whole range). When it is required to suppress the vigor of Norway spruce juveniles (e.g., the target being to encourage advanced artificial regeneration of reintroduced tree species during Norway spruce monoculture transformation), light intensity should not exceed this threshold by too much (to support reintroduced species). When Norway spruce is to be encouraged to reach advanced growing stages earlier, the light intensity should be over 0.2 TSF. However, regardless of these two main growth regimes, the growing response to different light conditions is continual after the threshold is exceeded, and therefore the encouraged heterogeneity of the overstory canopy openness leads to high growth diversity and also a high structure of Norway spruce regeneration. Therefore, variability of patches of overstory canopy created by a combination of sections with a closed canopy, mid-open canopy, and small gaps can be favorable for this purpose. Regeneration density did not correlate well with light intensities (and if some weak trends were found, they demonstrated a negative correlation with light). These results demonstrated that the regeneration establishment is more an issue of appropriate forest microclimate, where perhaps both temperature and humidity regimes play key roles and light is the further additional variable, which indicates that a shelter-wood cut can be more favorable for regeneration establishment than the creation of gaps. And after it reaches certain growing stage of about 1 m in height, intraspecific competition and autoreduction start to be driving forces of its density.




{kind=link}
{kind=link}
{kind=link}


