
Spatial Distribution Pattern of Root Sprouts under the Canopy of Malus sieversii in a Typical River Valley on the Northern Slopes of the Tianshan Mountain


Abstract
:1. Introduction
2. Methodology
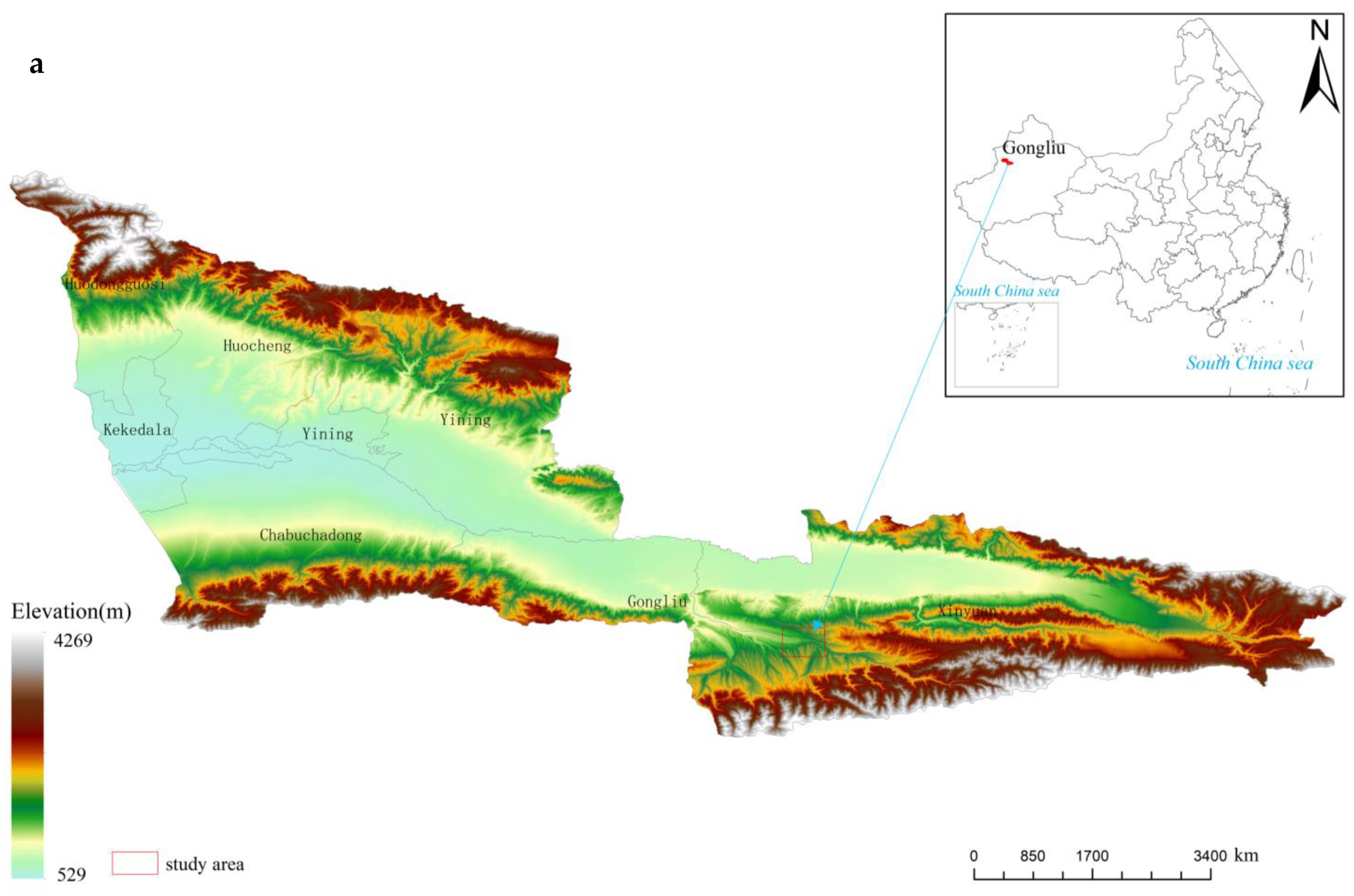
2.1. Study Site
2.2. Investigation Method
2.2.1. Plot Setting
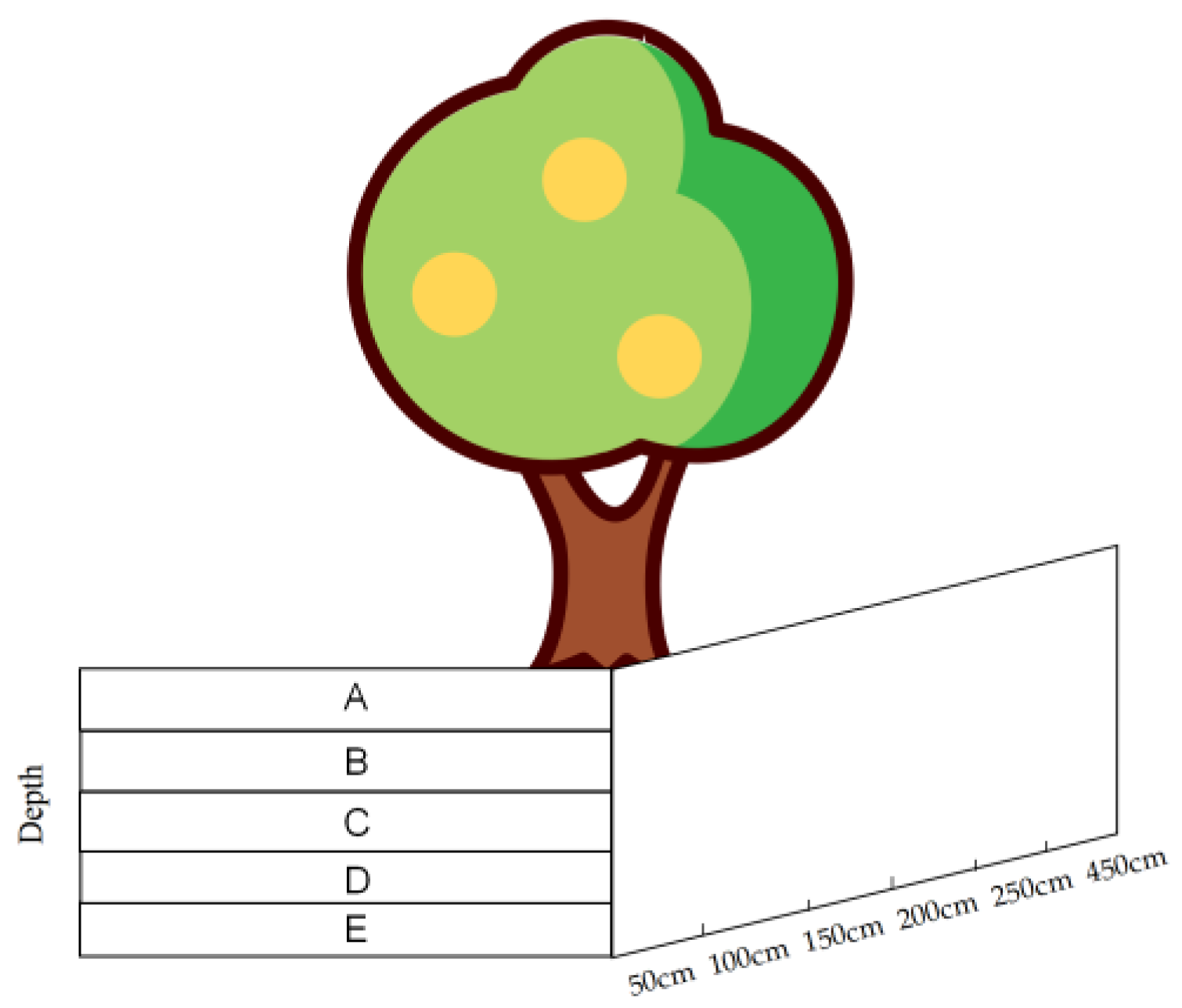
2.2.2. Root Survey
2.2.3. Root Sprout Survey
2.3. Research Methods
2.3.1. Spatial Clustering Pattern
2.3.2. Standard Deviation Ellipse
2.3.3. Principal Component Analysis
2.3.4. Grey Correlation Analysis
3. Results and Analysis
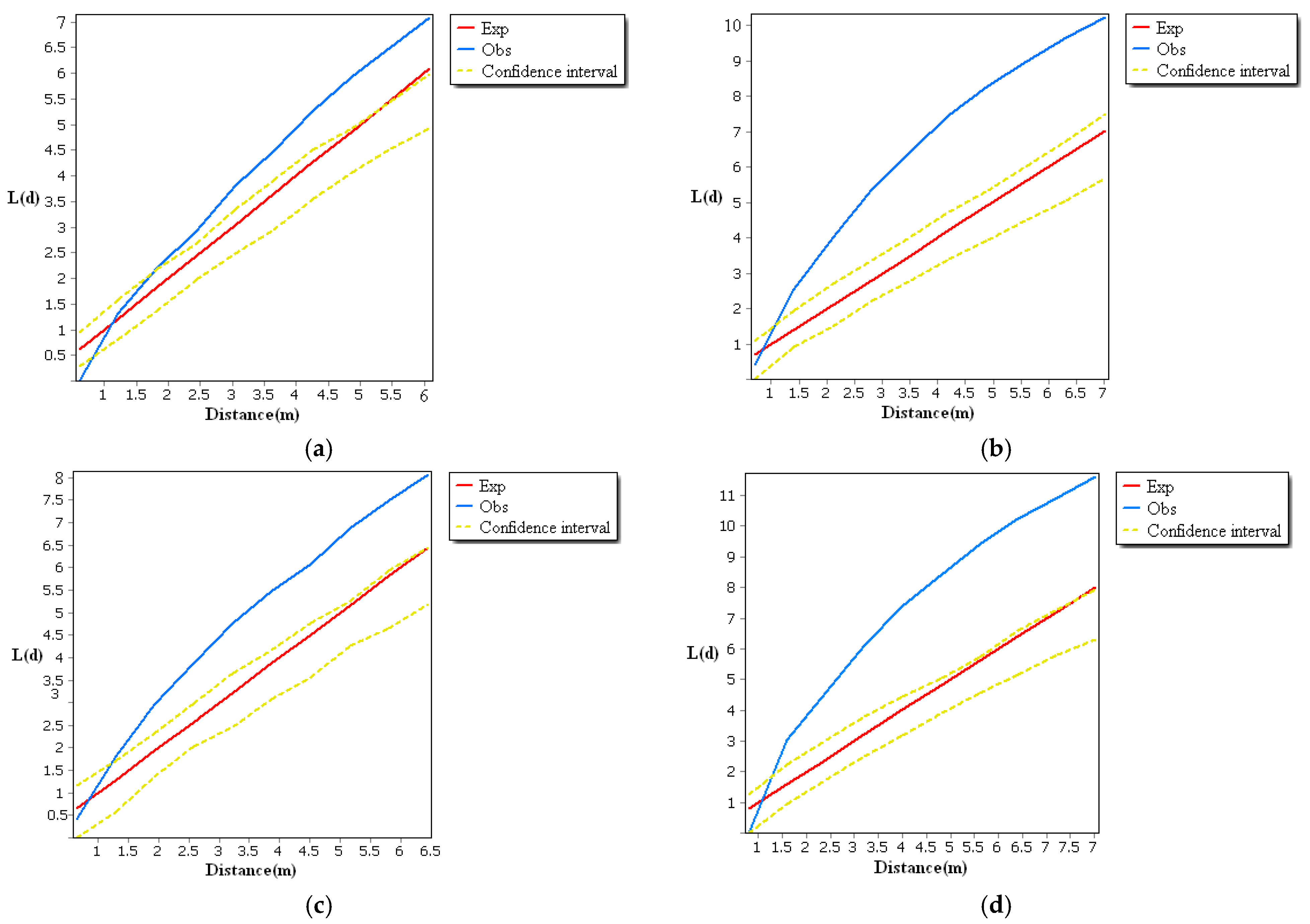
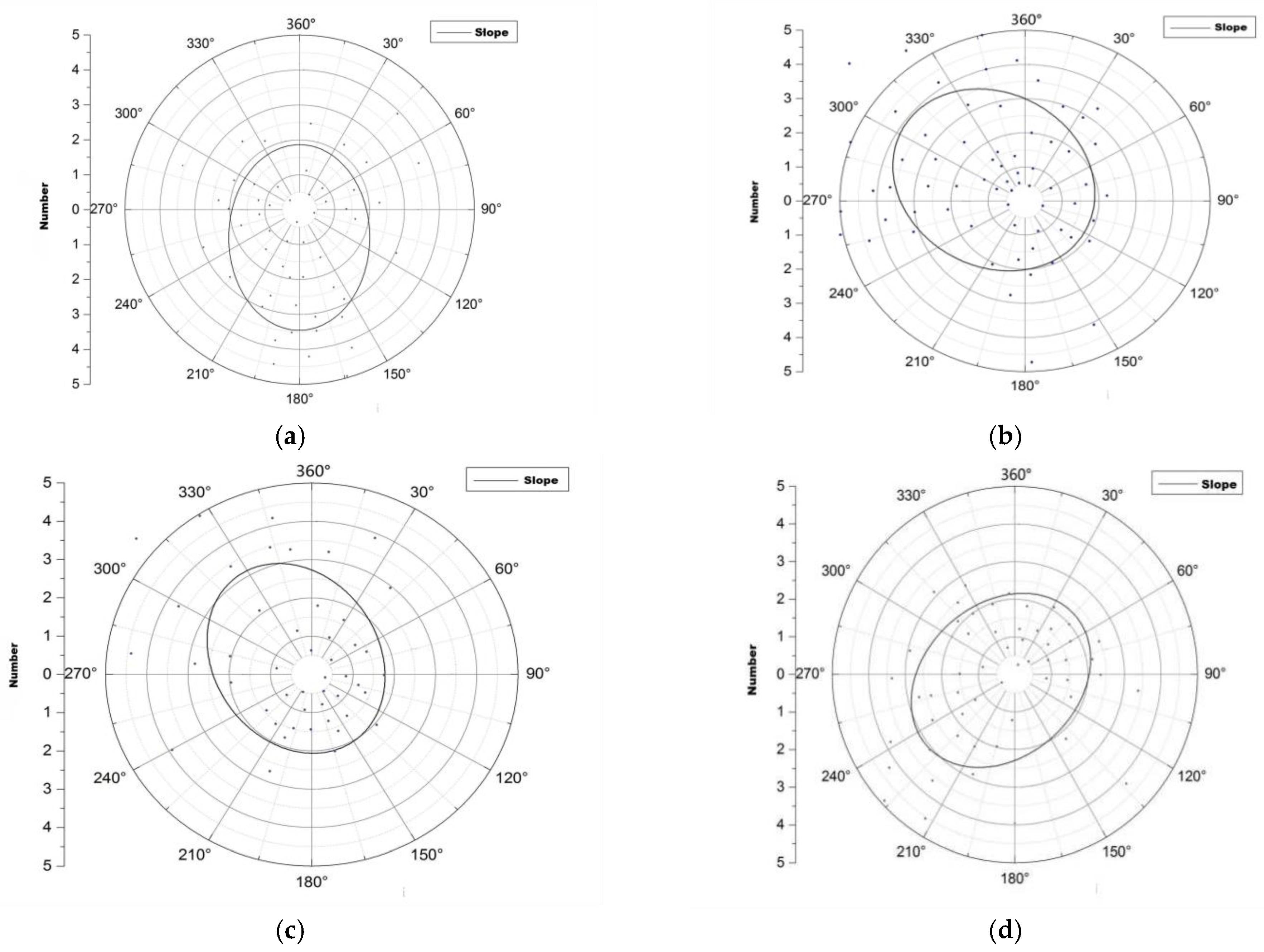
3.1. The Spatial Distribution Characteristics of Root Sprouts
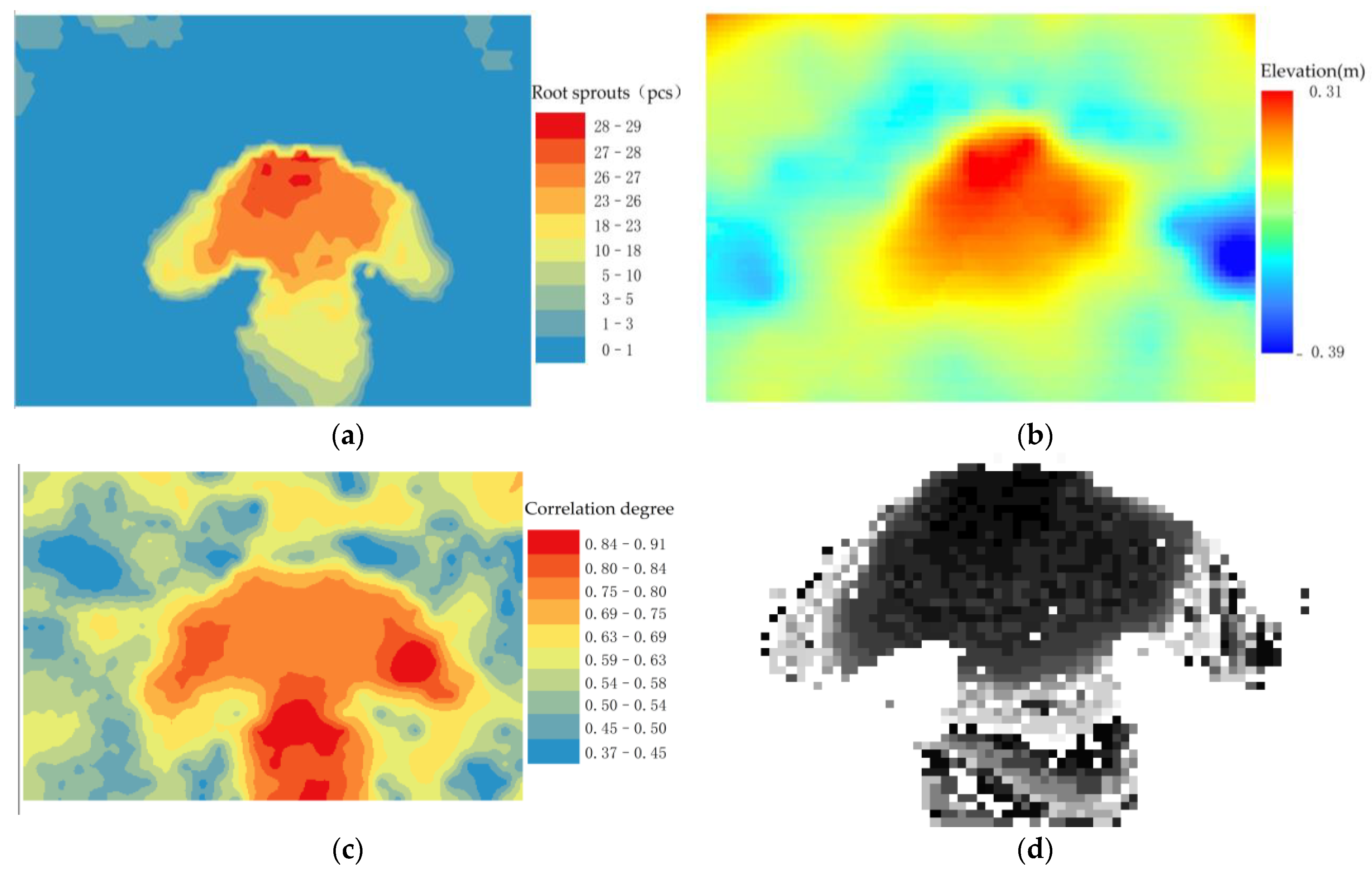
3.2. Impact of Environmental Factors on Root Sprouts Distribution
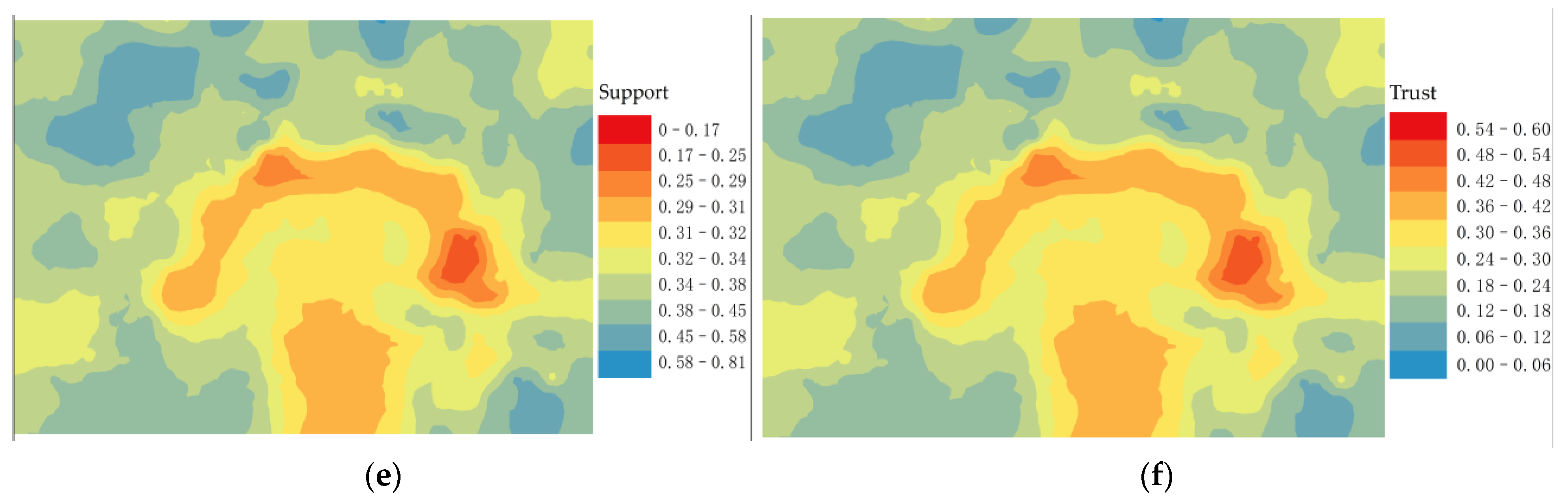
3.3. Spatial Correlation Analysis Study of Root Sprouting under the Tree Canopy
4. Discussion
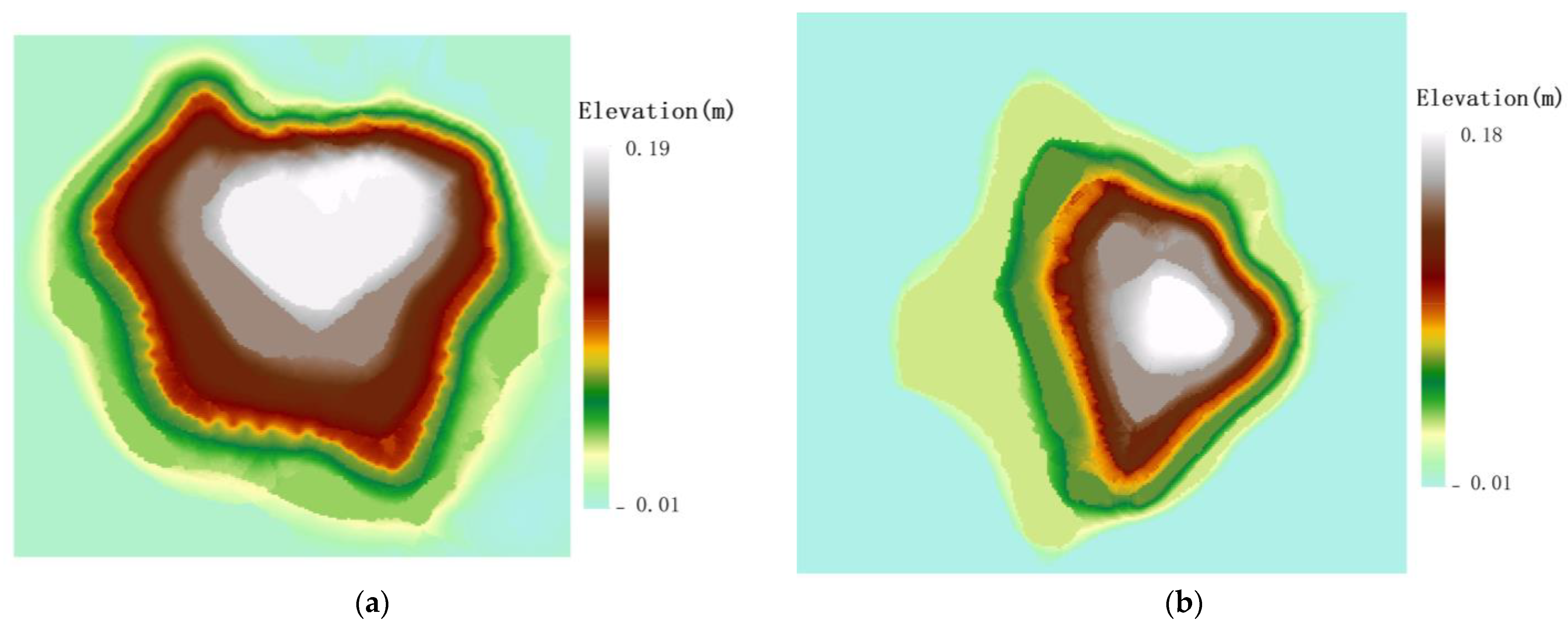
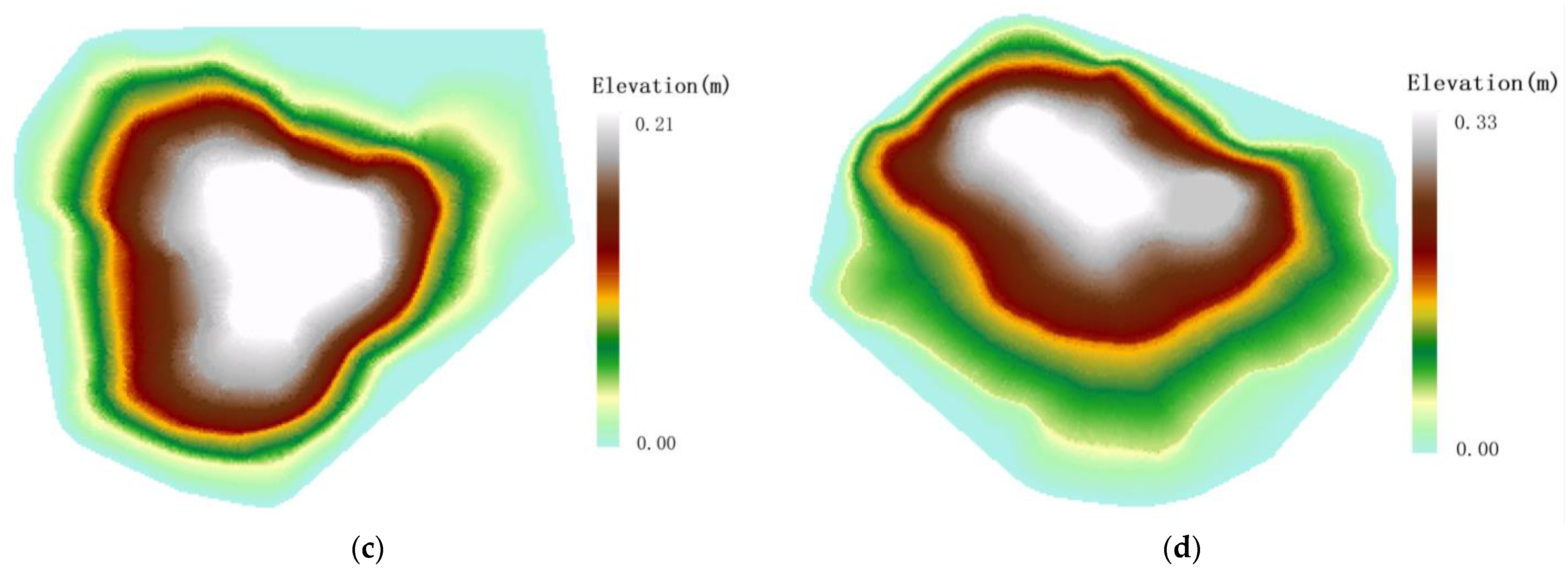
4.1. Spatial Distribution Characteristics of Root Sprouts in Malus sieversii
4.2. Analysis of the Correlation between Root Sprouting Conditions and Topography in Malus sieversii
4.3. Analysis of the Factors Influencing the Spatial Distribution of Root Sprouts in Malus sieversii
4.4. Suggestions for Conservation and Management
5. Conclusions
- (1)
- Spatial orientation and distance from the maternal plant were decisive for root sprouting, explaining 73.686% of the total variance.
- (2)
- The spatial distribution of root sprouts under the canopy of the Malus sieversii is aggregated and random. It has a significant directional distribution (mainly on the downslope), with the most polarized distribution of root sprouts on the western and northern slopes.
- (3)
- The grey correlation was used to spatially correlate the sub-canopy root sprouting point data with relative elevation to verify the scientific validity of the “clover-type” and to illustrate the role of tree platform topography on the root sprouting distribution of Malus sieversii. This study is of great significance for the recovery of natural populations of the Malus sieversii and for predicting the future development of populations. This study fills a gap in vegetative propagation and ecological research on the root sprouting of Malus sieversii.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.-X.; Li, X.-S.; Wang, J.-C.; Zhang, D.-Y. Insights into the aridification history of Central Asian Mountains and international conservation strategy from the endangered wild apple tree. J. Biogeogr. 2021, 48, 332–344. [Google Scholar] [CrossRef]
- Cornille, A.; Gladieux, P.; Smulders, M.J.M.; Roldán-Ruiz, I.; Laurens, F.; Le Cam, B.; Nersesyan, A.; Clavel, J.; Olonova, M.; Feugey, L.; et al. New Insight into the History of Domesticated Apple: Secondary Contribution of the European Wild Apple to the Genome of Cultivated Varieties. PLoS Genet. 2012, 8, e1002703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.J.; Liao, M.K. China’s precious Ili wild fruit forests in urgent need of conservation! Xinjiang For. 1986, 5, 30–31. [Google Scholar]
- Shan, Q.J.; Wang, Z.K.; Ling, H.B.; Zhang, G.P.; Yan, J.J.; Han, F.F. Unreasonable human disturbance shifts the positive effect of climate change on tree-ring growth of Malus sieversii in the origin area of world cultivated apples. J. Clean. Prod. 2021, 287, 125008. [Google Scholar] [CrossRef]
- Mierkamili, M.; Liu, Q.Z.; Ma, X.D.; Zhang, H.X.; Tian, Z.P. Suivival status, problems and conservation strategies of Malus sieversii. Guihaia 2021, 41, 2100–2109. [Google Scholar]
- Wang, L.; Sun, Q.W.; Hao, C.Y.; Tian, S.N.; Zhang, S.S.; Chen, Y.K.; Zhang, X.P. Point pattern analysis of different age–class Taxus chinensis var. mairei individuals in mountainous area of southern Anhui Province. Chin. J. Appl. Ecol. 2010, 21, 272–278. [Google Scholar]
- Zhang, Q.J.; Wang, Y.X. The study progress on natural regeneration strategies of Oplopanax elatus Nakai. Seed 2014, 33, 52–55. [Google Scholar]
- Xue, P.P.; Guo, L.J.; Wan, F.; Mao, X.T.; Xiao, Z.P.; Wu, Y.P.; Wang, B.Y.; Zeng, Y.; Fan, H.X. Comparison of the population characteristics of the root sprout and seedling of the endangered plant Emmenopterys henryi. J. Tianjin Norm. Univ. 2022, 42, 38–44. [Google Scholar]
- Zhang, T.; Song, L.N.; Zhu, J.J.; Wang, G.C.; Li, M.C.; Zheng, X.; Zhang, J.X. Spatial distribution of root systems of Pinus sylvestris var. mongolica trees with different ages in a semi-arid sandy region of Northeast China. For. Ecol. Manag. 2021, 483, 118776. [Google Scholar] [CrossRef]
- Hokanson, S.C.; Forsline, P.L.; McFerson, J.R.; Lamboy, W.F.; Aldwinckle, H.S.; Luby, J.J.; Djangaliev, A.D. Ex Situ and In Situ conservation strategies for wild Malus germplasm in Kazakhstan. Eucarpia Symp. Fruit Breed. Genet. 1998, 484, 85–91. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Chen, X.S.; Zhang, Y.M.; Yuan, Z.H.; Liu, Z.C.; Wang, Y.L.; Lin, Q. A method for constructing core collection of Malus sieversii using molecular markers. Agric. Sci. China 2009, 8, 276–284. [Google Scholar] [CrossRef]
- Cornille, A.; Gladieux, P.; Giraud, T. Crop-to-wild gene flow and spatial genetic structure in the closest wild relatives of the cultivated apple. Evol. Appl. 2013, 6, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.Q.; Huang, T.C.; Chen, S.J.; Chen, M.Y.; Jia, X.; Zhu, X.; Lai, F.B.; Wu, H.G.; Zhao, H.X.; Li, C.L.; et al. Quantitative evaluation on pest damage levels of Malus sieversii based on spectral reflectance. Jiangsu J. Agric. Sci. 2019, 35, 798–803. [Google Scholar] [CrossRef] [Green Version]
- Den, X. Study on Cultivation Techniques of Malus sievesii Suckers. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2020. [Google Scholar]
- Li, L.; Chen, S.J.; Chen, M.Y.; Zhu, X.; Lai, F.B. Environmental driving force of stool shoot quantitative features of Malus sieversii (Ldb.) Roem in Xiaomohe valley. Ecol. Sci. 2019, 38, 18–24. [Google Scholar]
- Wei, S.G.; Li, L.; Xu, R.; Huang, Z.L.; Cao, H.L. Spatial Pattern and Interspecific Relationship of Dominant Species in Plant Community in Jinggang Mountain. J. Trop. Subtrop. Bot. 2015, 23, 74–80. [Google Scholar]
- Zhang, Z.H.; Lai, F.B.; Chen, S.J.; Sun, H. Tree-Dimensional Shape Characteristics and Spatial Auto-Correlationship of Pop-ulus eu phratica Dune in the Sparse Forest Sandy Land on the West of Hotan River. Res. Soil Water Conserv. 2015, 22, 137–142+149. [Google Scholar]
- Xu, S.W.; Lai, F.B.; Chen, M.Y.; Zhu, X.; Li, H.J.; Chen, S.J.; Huang, T.C.; Jia, X. Morphological characteristics and its clinal variation of the tree platform of Malus sieversii. Arid. Land Geogr. 2019, 42, 305–313. [Google Scholar]
- Tengberg, A.; Chen, D. A comparative analysis of nebkhas in central Tunisia and northern Burkina Faso. Geomorphology 1998, 22, 181–192. [Google Scholar] [CrossRef]
- Fraser, C.S.; Cronk, S. A hybrid measurement approach for close-range photogrammetry. ISPRS J. Photogramm. Remote Sens. 2009, 64, 328–333. [Google Scholar] [CrossRef]
- Gonçalves, J.; Henriques, R.; Alves, P.; Sousa-Silva, R.; Monteiro, A.T.; Lomba, Â.; Marcos, B.; Honrado, J. Evaluating an unmanned aerial vehicle-based approach for assessing habitat extent and condition in fine-scale early successional mountain mosaics. Appl. Veg. Sci. 2016, 19, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Bendig, J.; Yu, K.; Aasen, H.; Bolten, A.; Bennertz, S.; Broscheit, J.; Gnyp, M.L.; Bareth, G. Combining UAV-based plant height from crop surface models, visible, and near infrared vegetation indices for biomass monitoring in barley. Int. J. Appl. Earth Obs. Geoinf. 2015, 39, 79–87. [Google Scholar] [CrossRef]
- Wu, J.; Wang, P.; Wang, J.; Wu, X.H.; Su, X.H. Low Altitude Photogrammetry of Sea Islands and Reefs without Ground Con-trol Points: Research and Application. J. Yangtze River Sci. Res. Inst. 2022, 39, 133–137. [Google Scholar]
- Martens, S.N.; Breshears, D.D.; Meyer, C.W. Spatial distributions of understory light along the grassland/forest continuum: Effects of cover, height, and spatial pattern of tree canopies. Ecol. Model. 2000, 126, 79–93. [Google Scholar] [CrossRef]
- Greig-Smith, P. Quantitative Plant Ecology, 3rd ed.; Blackwell Science Publications: Oxford, UK, 1983. [Google Scholar]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Interscience: New York, NY, USA; London, UK, 1969. [Google Scholar]
- Peng, S.L. South Asia Tropical Forest Communit Dynamics; Science Press: Beijing, China, 1996. [Google Scholar]
- Zhang, J.T. Analysis of spatial point patiern for plant species. J. Plant Ecol. 1998, 22, 344–349. [Google Scholar]
- Velázquez, E.; Martínez, I.; Getzin, S.; Moloney, K.A.; Wiegand, T. An evaluation of the state of spatial point pattern analysis in ecology. Ecography 2016, 39, 1042–1055. [Google Scholar] [CrossRef]
- Muvengwi, J.; Mbiba, M.; Chikumbindi, J.; Ndagurwa, H.G.; Mureva, A. Population structure and spatial point-pattern analysis of a mono stand of Acacia polyacantha along a catena in a savanna ecosystem. For. Ecol. Manag. 2018, 409, 499–508. [Google Scholar] [CrossRef]
- Kong, X. Study on Root Sprouting Characteristics and Driving Factors of Populus euphratica in the Birkum. Master’s Thesis, Xinjiang Normal University, Urumqi, China, 2021. [Google Scholar]
- Yang, X.P.; Wang, H.O.; Jiang, L.L.; Zhou, L.; Wang, J. Age Structure Analysis of Different Malus sieversii Population in Yili Areas. Mol. Plant Breed. 2021, 19, 4133–4141. [Google Scholar]
- Yu, S.S.; Zhao, W.X.; Yao, Y.X.; Tan, W.Q.; Yan, W.; Song, W.W. Investigation of habitat conditions of wild apples in Xinjiang. In Proceedings of the 2019 Annual Academic Conference and 90th Anniversary Conference of the Chinese Society of Horti-cultural Science, Zhengzhou, China, 21 October 2019. [Google Scholar]
- Huang, T.C. Withered Branches Grade Monitoring in Malus sieversii Forest Using Hyperspectral Data. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2021. [Google Scholar]
- Wang, P.J.; Zhang, H.F.; Tian, X.; Zhang, J.L.; Zhu, Y.L. Estimation of Forest Stem Volume of Natural Forests based on the Optimization of Active and Passive Remote Sensing Features. Remote Sens. Technol. Appl. 2022, 37, 672–680. [Google Scholar]
- She, D.Y. Formation Conditions, Process and Characteristics of Soil in Ili Wild Fruit Forest. Chin. J. Soil Sci. 1995, 26, 260–263. [Google Scholar]
- Liu, L.C.; Pai, Z.L.; Xu, J.H. The formation characteristics and taxonomic classification of the soil under the wild-frult forests in Yil valley, Xinjiang. Arid. Land Geogr. 1997, 20, 34–40. [Google Scholar]
- Salas, C.; LeMay, V.; Núñez, P.; Pacheco, P.; Espinosa, A. Spatial patterns in an old-growth Nothofagus obliqua forest in south-central Chile. For. Ecol. Manag. 2006, 231, 38–46. [Google Scholar] [CrossRef]
- Guo, K.; Yuan, Y. Research on Spatial and Temporal Evolution Trends and Driving Factors of Green Residences in China Based on Weighted Standard Deviational Ellipse and Panel Tobit Model. Appl. Sci. 2022, 12, 8788. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, L.; Xu, W.; Ledwith, V. Use of local Moran’s I and GIS to identify pollution hotspots of Pb in urban soils of Galway, Ireland. Sci. Total Environ. 2008, 389, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Min, Z.Q.; Wu, B.G.; Su, X.H.; Chen, Y.L.; Tian, Y.Z. Suitability Evaluation and Dominant Function Model for Multifunctional Forest Management. Forests 2020, 12, 1368. [Google Scholar] [CrossRef]
- Gao, R.; Ai, N.; Liu, G.; Liu, C.; Qiang, F.; Zhang, Z.; Xiang, T.; Zang, K. The Coupling Relationship between Herb Communities and Soil in a Coal Mine Reclamation Area after Different Years of Restoration. Forests 2022, 13, 1481. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Wang, Y.S.; Luo, P.Z.; He, G.X. Coupling relationships between plant diversity and soil characteristics in rocky desertification areas of western Hunan. J. For. Environ. 2021, 41, 471–477. [Google Scholar]
- Yan, Y.; L, Y.G.; Sun, M.H.; Wu, Z.H. Primary Pollutants and Air Quality Analysis for Urban Air in China: Evidence from Shanghai. Sustainability 2019, 11, 2319. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.K.; Jiao, A.Y.; Ling, H.B.; Shan, Q.J.; Wang, W.Q. Characteristics of Populus euphratica root under various irrigation modes. Arid. Zone Res. 2022, 39, 1133–1142. [Google Scholar] [CrossRef]
- Valbuena-Carabaña, M.; Gil, L. Genetic resilience in a historically profited root sprouting oak (Quercus pyrenaica Willd.) at its southern boundary. Tree Genet. Genomes 2013, 9, 1129–1142. [Google Scholar] [CrossRef]
- Guo, L.J.; Shao, X.H.; Xue, P.P.; Tian, Y.Q.; Xiao, Z.P.; Wu, Y.P. Root sprouting ability and growth dynamics of the rootsuckers of Emmenopterys henryi, a rare and endangered plant endemic to China. For. Ecol. Manag. 2017, 389, 35–45. [Google Scholar] [CrossRef]
- Bartušková, A.; Filartiga, A.L.; Herben, T.; Qian, J.; Klimešová, J. Comparative analysis of root sprouting and its vigour in temperate herbs: Anatomical correlates and environmental predictors. Ann. Bot. 2021, 127, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Ye, X.H.; Chen, F.; Yang, Q.; Li, Z.Y.; Fang, K.; Yang, G.Y. Adaptation of bamboo to hetergenerous habital: Pheno-typic plasticity. Acta Ecol. Sin. 2014, 34, 5687–5695. [Google Scholar]
- CHEN, R.; WANG, H.N.; JIANG, Y.M.; WEI, S.H.; CHEN, Q.; GE, S.F. Rhizosphere Soil Microbial Quantity and Enzyme Ac-tivity of Different Apple Rootstocks. Chin. Agric. Sci. 2012, 45, 2099–2106. [Google Scholar]
- Li, H.B.; Liu, Y.; Taxmamat, M.; Zhao, Y.; Lv, D.K. Natural distribution characteristics of endangered wild cherry plum seed-lings in Xinjiang. Xinjiang Agric. Sci. 2013, 9, 1612–1619. [Google Scholar]
- Lin, Y.-C.; Chang, L.-W.; Yang, K.-C.; Wang, H.-H.; Sun, I.-F. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia 2010, 165, 175–184. [Google Scholar] [CrossRef]
- Ta, F.; Liu, X.-D.; Liu, R.-H.; Zhao, W.-J.; Jing-Zhong, Z.; Wen-Mao, J.; Wu, X.-R.; Zhao, J.-Z.; Ma, X.-E. Spatial distribution patterns and association of Picea crassifolia population in Dayekou Basin of Qilian Mountains, northwestern China. Chin. J. Plant Ecol. 2020, 44, 1172–1183. [Google Scholar] [CrossRef]
- Wang, Q.G.; Punchi-Manage, R.; Lu, Z.J.; Franklin, S.; Wang, Z.H.; Li, Y.Q.; Chi, X.L.; Bao, D.C.; Guo, Y.L.; Lu, J.; et al. Effects of topography on structuring species assemblages in a subtropical forest. J. Plant Ecol. 2017, 10, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.G.; Xu, Y.Z.; Lu, Z.J.; Bao, D.C.; Guo, Y.L.; Lu, J.M.; Zhang, K.H.; Liu, H.B.; Meng, H.J.; Qiao, X.J.; et al. Disentangling the effects of topography and space on the distributions of dominant species in a subtropical forest. Chin. Sci. Bull. 2014, 59, 5113–5122. [Google Scholar] [CrossRef]
- Sun, R.X.; Ma, L.; Zhang, S.H.; Yu, Y.; Shen, M.S.; Zhang, H.B.; Wang, D.D.; Yang, Y.B.; Zhang, J.N.; Zhang, Y.Z.; et al. Study on Landscape Patches Influencing Hillslope Erosion Processes and Flow Hydrodynamics in the Loess Plateau of Western Shanxi Province, China. Water 2020, 12, 3201. [Google Scholar] [CrossRef]
- Rafiee, F.; Ejtehadi, H.; Farzam, M.; Zare, H.; Bashirzadeh, M. Changes in plant biodiversity facets of rocky outcrops and their surrounding rangelands across precipitation and soil gradients. Sci. Rep. 2022, 12, 12518. [Google Scholar] [CrossRef]
- Palacios-Cabrera, T.; Valdes-Abellan, J.; Jodar-Abellan, A.; Rodrigo-Comino, J. Land-use changes and precipitation cycles to understand hydrodynamic responses in semiarid Mediterranean karstic watersheds. Sci. Total. Environ. 2022, 819, 153182. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Zhang, L.P.; Wang, Y. Effect of Slope Length and Rainfall Intensity on Runoff and Erosion Conversion from Laboratory to Field. Water Resour. 2019, 46, 530–541. [Google Scholar] [CrossRef]
- Pan, D.; Gao, X.; Dyck, M.; Song, Y.; Wu, P.; Zhao, X. Dynamics of runoff and sediment trapping performance of vegetative filter strips: Run-on experiments and modeling. Sci. Total Environ. 2017, 593–594, 54–64. [Google Scholar] [CrossRef]
- Zhao, W.Y.; Chen, Y.N.; Zhou, H.H.; Zhou, X.M.; Wang, X.P. Reproductive ability and relative environment factors of de-graded Populus euphratica forest in ecological water delivery project ai tlwer reaches of Tarim River. J. Desert Res. 2009, 29, 108–113. [Google Scholar]
- Nagamatsu, D.; Miura, O. Soil disturbance regime in relation to micro-scale landforms and its effects on vegetation structure in a hilly area in Japan. Plant Ecol. 1997, 133, 191–200. [Google Scholar] [CrossRef]
- McEwan, R.W.; Lin, Y.-C.; Sun, I.-F.; Hsieh, C.-F.; Su, S.-H.; Chang, L.-W.; Song, G.-Z.M.; Wang, H.-H.; Hwong, J.-L.; Lin, K.-C.; et al. Topographic and biotic regulation of aboveground carbon storage in subtropical broad-leaved forests of Taiwan. For. Ecol. Manag. 2011, 262, 1817–1825. [Google Scholar] [CrossRef]
- Hu, X.H.; Ren, Y.L.; Zhang, Y.X. Morphological variation of lithocarpus glaber seeds in evergreen broadleaved forest at tis different succession stages in Gutian Mountain. Chin. J. Ecol. 2012, 31, 2449–2454. [Google Scholar]
- Hoyle, G.L.; Venn, S.; Steadman, K.; Good, R.B.; McAuliffe, E.J.; Williams, E.R.; Nicotra, A. Soil warming increases plant species richness but decreases germination from the alpine soil seed bank. Glob. Chang. Biol. 2013, 19, 1549–1561. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Multi-stemmed trees in montane rain forests: Their frequency and demography in relation to elevation, soil nutrients and disturbance. J. Ecol. 2009, 97, 472–483. [Google Scholar] [CrossRef]
- Shi, L.L.; Luo, Z.R.; Xia, J.T.; Zhao, W.J.; Wu, G.Y.; Ding, B.Y. Woody seedling dynamics and the correlation between habitat and regeneration / mortality in a subtropical evergreen broad-leaved forest in China. Acta Ecol. Sin. 2014, 34, 6510–6518. [Google Scholar]
- Zhang, C.Q.; Zhang, W.H. A study on asexual reproduction and regeneration of Robinia pseudoacacia plantations in different habitats in Hilly Area of the Loess Plateau. J. Northwest A F Univ. 2009, 37, 135–144. [Google Scholar]
- Ye, J.; Hao, Z.Q.; Wang, X.G.; Bai, X.J.; Xing, D.L.; Yuan, Z.Q. Local-scale drivers of multi-stemmed tree formation in Acer, in a temperate forest of Northeast China. Chin. Sci. Bull. 2014, 59, 320–325. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aspect | Eastern Slope | Western Slope | Southern Slope | North Slope |
|---|---|---|---|---|
| Rotation θ | 130.64° | 1.21° | 152.86° | 51.80° |
| Standard deviation along the Y-axis/m | 7.61° | 8.09° | 6.06° | 7.86° |
| Standard deviation along the X-axis/m | 6.51° | 6.15° | 4.77° | 6.07° |
| Semi-axes/m | 1.10° | 1.94° | 1.29° | 1.79° |
| Composition | Spatial Orientation | Distance | Relative Elevation | Buried Thickness | Root Sprout Thickness | Root Sprout Height |
|---|---|---|---|---|---|---|
| Spatial Orientation | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Distance | −0.67 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Relative Elevation | 0.56 | −0.21 | 1.00 | 0.00 | 0.00 | 0.00 |
| Buried Thickness | 0.83 | −0.52 | 0.64 | 1.00 | 0.00 | 0.00 |
| Root Sprout Thickness | 0.18 | −0.10 | 0.10 | 0.20 | 1.00 | 0.00 |
| Root Sprout Height | 0.56 | −0.18 | 0.65 | 0.81 | 0.30 | 1.00 |
| Principal Component | Eigenvalues | Contribution Rate (%) | Accumulative Contribution Rate (%) |
|---|---|---|---|
| Z1 | 3.38 | 56.35 | 56.35 |
| Z2 | 1.04 | 17.34 | 73.69 |
| Z3 | 0.92 | 15.39 | 89.08 |
| Z4 | 0.38 | 6.25 | 95.33 |
| Z5 | 0.20 | 3.39 | 98.72 |
| Z6 | 0.08 | 1.28 | 100.00 |
| Composition | Z1 | Z2 |
|---|---|---|
| Spatial Orientation | 0.89 | −0.29 |
| Distance | −0.60 | 0.67 |
| Relative Elevation | 0.95 | 0.22 |
| Buried Thickness | 0.75 | −0.02 |
| Root Sprout Thickness | 0.31 | 0.54 |
| Root Sprout Height | 0.82 | 0.41 |
| Slope | Eastern | Western | Southern | Northern |
|---|---|---|---|---|
| Upslope | 180°–359° | 0°–179° | 0°–89°, 270°–359° | 90°–269° |
| Downslope | 0°–179° | 180°–359° | 90°–269° | 270°–359°, 0°–89° |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Chen, M.; Zhang, X.; Jia, X. Spatial Distribution Pattern of Root Sprouts under the Canopy of Malus sieversii in a Typical River Valley on the Northern Slopes of the Tianshan Mountain. Forests 2022, 13, 2044. https://doi.org/10.3390/f13122044
Li L, Chen M, Zhang X, Jia X. Spatial Distribution Pattern of Root Sprouts under the Canopy of Malus sieversii in a Typical River Valley on the Northern Slopes of the Tianshan Mountain. Forests. 2022; 13(12):2044. https://doi.org/10.3390/f13122044
Chicago/Turabian StyleLi, Li, Mengyu Chen, Xiaoli Zhang, and Xiang Jia. 2022. "Spatial Distribution Pattern of Root Sprouts under the Canopy of Malus sieversii in a Typical River Valley on the Northern Slopes of the Tianshan Mountain" Forests 13, no. 12: 2044. https://doi.org/10.3390/f13122044
APA StyleLi, L., Chen, M., Zhang, X., & Jia, X. (2022). Spatial Distribution Pattern of Root Sprouts under the Canopy of Malus sieversii in a Typical River Valley on the Northern Slopes of the Tianshan Mountain. Forests, 13(12), 2044. https://doi.org/10.3390/f13122044