
Establishment and Natural Regeneration of Native Trees in Agroforestry Systems in the Paraguayan Atlantic Forest


Abstract
:1. Introduction
2. Materials and Methods
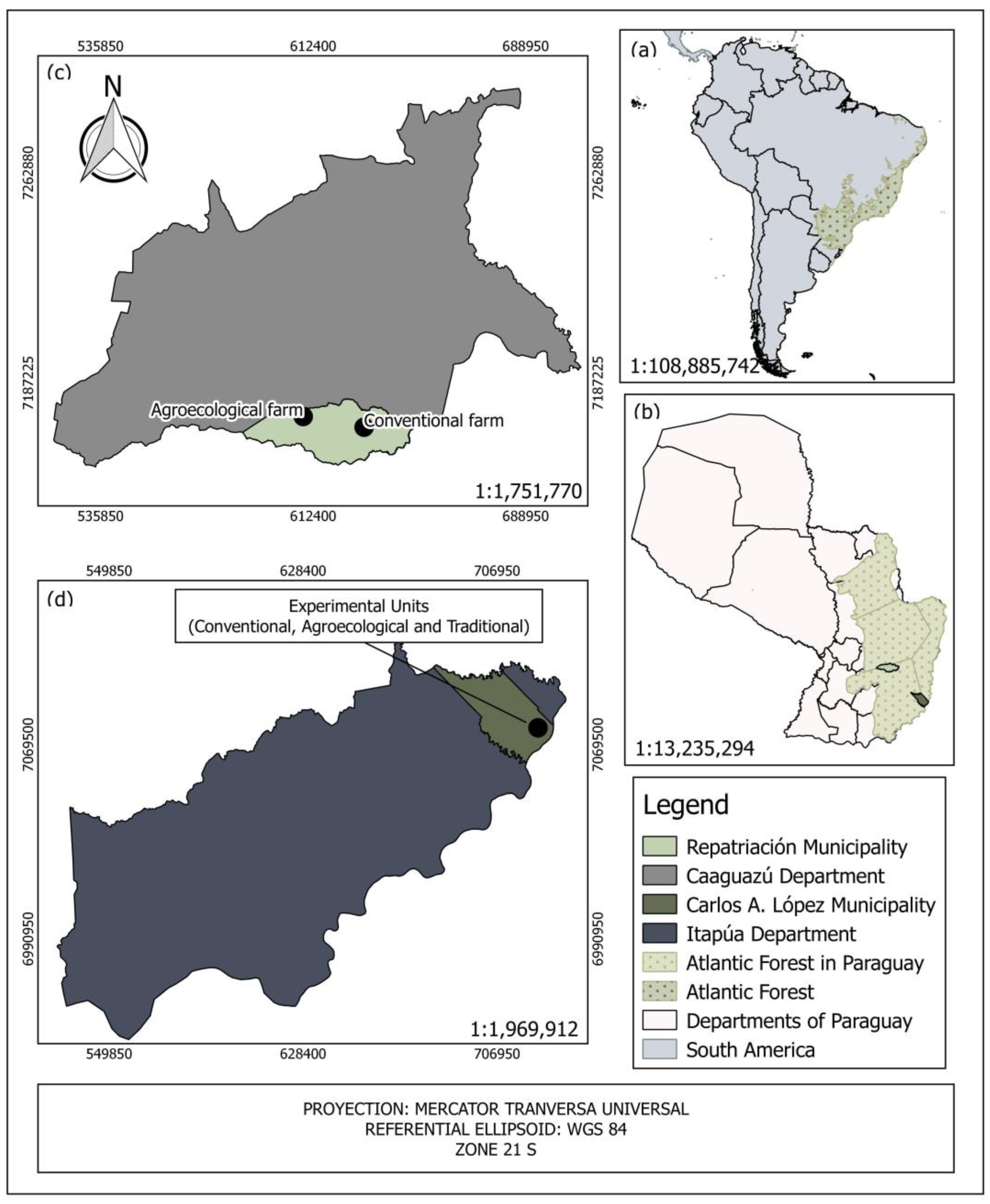
2.1. Study Area
2.2. Experimental Description
2.3. Monitoring the Establishment and Natural Regeneration of Tree Species
2.4. Data Analysis
3. Results
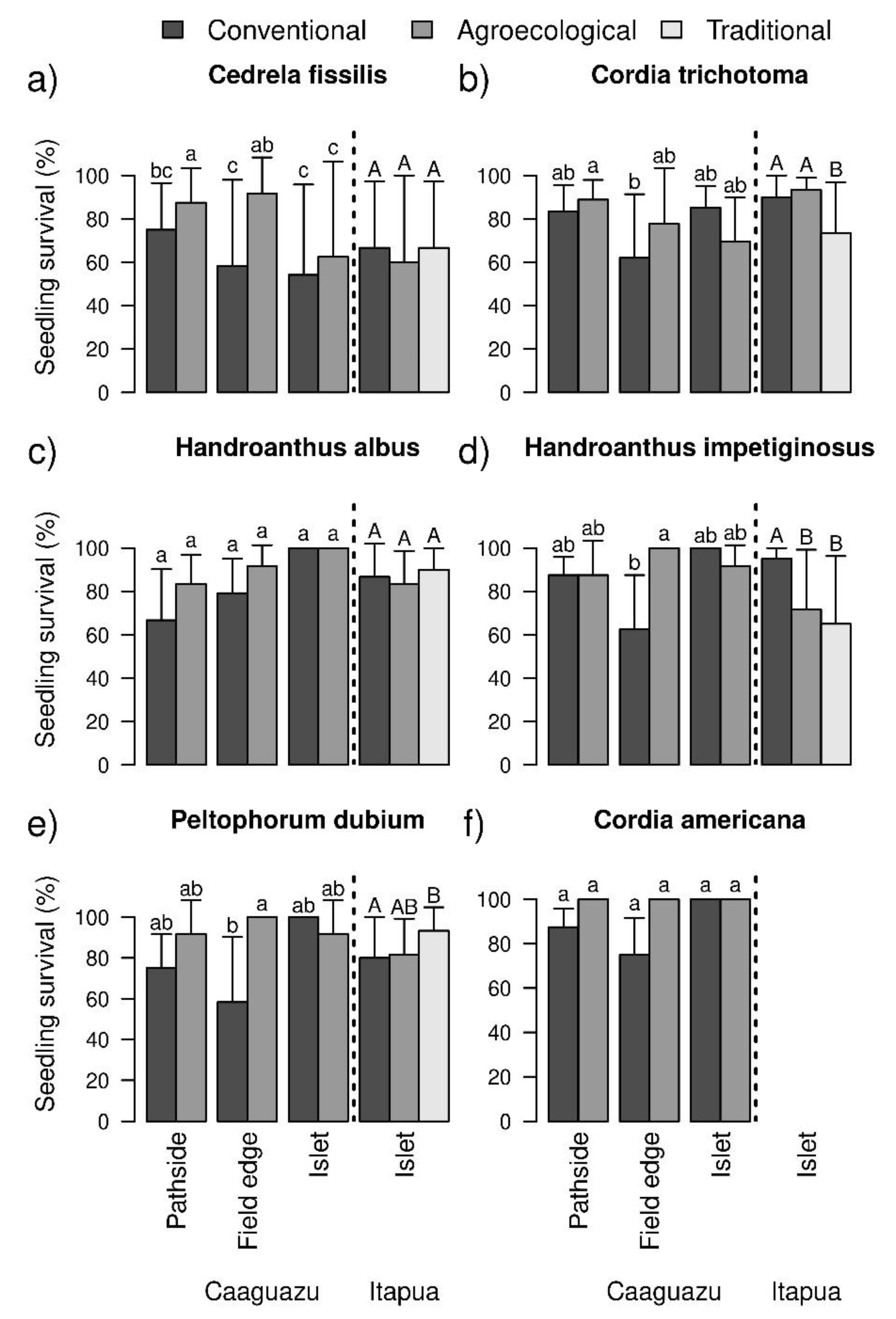
3.1. Survival of Planted Species
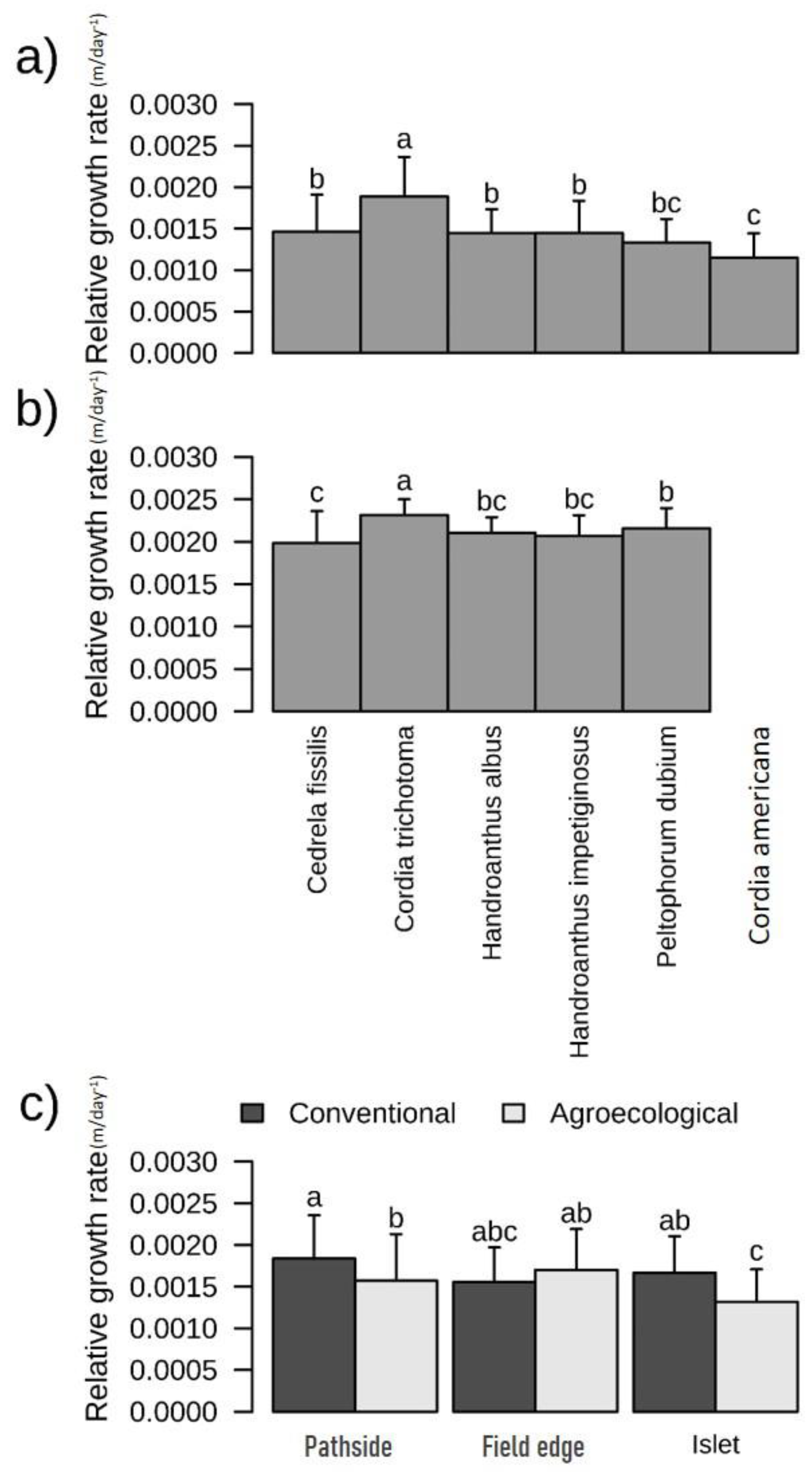
3.2. Growth of Planted Species
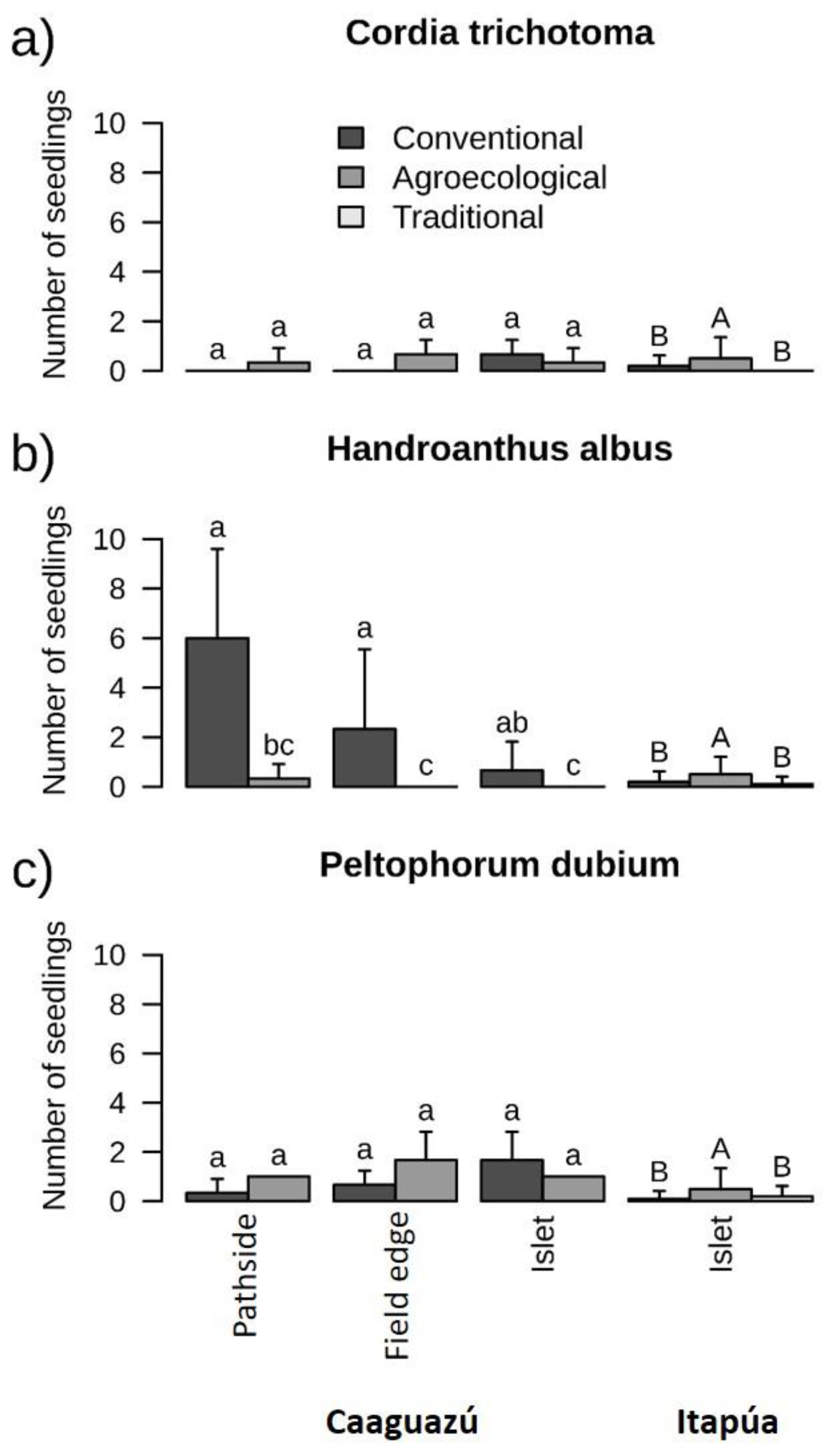
3.3. Natural Regeneration
4. Discussion
4.1. Establishment and Natural Regeneration Depend on the Species
4.2. Effects of the Agronomic System and Plantation Types on Establishment and Natural Regeneration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morantes-Toloza, J.L.; Renjifo, L.M. Cercas vivas en sistemas de producción tropicales: Una revisión mundial de los usos y percepciones. Rev. Biol. Trop. 2018, 66, 739–753. [Google Scholar] [CrossRef] [Green Version]
- World Wildlife Fund. La Degradación de los Bosques: Por Qué Afecta a las Personas y la Vida Silvestre. 2019. Available online: https://www.worldwildlife.org/descubre-wwf/historias/la-degradacion-de-los-bosques-por-que-afecta-a-las-personas-y-la-vida-silvestre (accessed on 20 April 2020).
- Freire Santos, P.Z.; Crouzeilles, R.; Boelsums, J.; Sansevero, B. Can agroforestry systems enhance biodiversity and ecosystem service provision in agricultural landscapes? A meta-analysis for the Brazilian Atlantic Forest. For. Ecol. Manag. 2019, 433, 140–145. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; Brigid, A.; McKenna, A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- FAO; PNUMA. El Estado de los Bosques del Mundo 2020. Los Bosques, la Biodiversidad y las Personas; FAO: Roma, Italy, 2020; 224p. [Google Scholar]
- FAO. Evaluación de los Recursos Forestales Mundiales 2020—Informe Principal; FAO: Roma, Italy, 2021. [Google Scholar] [CrossRef]
- BirdLife International. State of the World’s Birds’—Global Assessment Takes Pulse of the Planet’s Extinction Threat. Available online: https://www.birdlife.org/europe-and-central-asia/pr/ (accessed on 23 April 2018).
- Palm, C.; Blanco-Canqu, H.; DeClerck, G.; Gatere, L.; Grace, P. Conservation agriculture and ecosystem services: An overview. Agric. Ecosyst. Environ. 2014, 187, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Diniz, M.F.; Dallmeier, F.; Gregory, T.; Martinez, V.; Saldivar-Bellassai, S.; Benitez-Stanley, M.A.; Sánchez-Cuervo, A.M. Balancing multi-species connectivity and socio-economic factors to connect protected areas in the Paraguayan Atlantic Forest. Landsc. Urban Plan. 2022, 222, 104400. [Google Scholar] [CrossRef]
- Rey Benayas, J.M.; Bullock, J.M. Restoration of Biodiversity and Ecosystem Services on Agricultural Land. Ecosystems 2012, 15, 883–899. [Google Scholar]
- FAO. Agroforestry for landscape restoration. In Exploring the Potential of Agroforestry to Enhance the Sustainability and Resilience of Degraded Landscapes; FAO: Rome, Italy, 2017; 28p. [Google Scholar]
- Viswanath, S.; Lubina, P.A.; Subbanna, S.; Sandhya, M.C. Traditional Agroforestry Systems and Practices: A Review. Adv. Agric. Res. Technol. J. 2018, 2, 2–22. [Google Scholar]
- Jose, S. Agroforestry for conserving and enhancing biodiversity. Agrofor. Syst. 2012, 85, 1–8. [Google Scholar] [CrossRef]
- Haggar, J.; Pons, D.; Saenz, L.; Vides, M. Contribution of agroforestry systems to sustaining biodiversity in fragmented forest landscapes Agriculture. Ecosyst. Environ. 2019, 283, 106567. [Google Scholar] [CrossRef]
- Vihervaara, P.; Marjokorpi, A.; Kumpula, T.; Walls, M.; Kamppinen, M. Ecosystem services of fast-growing tree plantations: A case study on integrating social valuations with land-use changes in Uruguay. For. Policy Econ. 2012, 14, 58–68. [Google Scholar] [CrossRef]
- Oliveira, R.E.; Carvalhaes, M.A. Agroforestry as a tool for restoration in atlantic forest: Can we find multi-purpose species? Oecologia Aust. 2016, 20, 425–435. [Google Scholar] [CrossRef]
- Dhanya, B.; Sathish, B.N.; Viswanath, S.; Purushothaman, S. Ecosystem services of native trees: Experiences from two traditional agroforestry systems in Karnataka, Southern India. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2014, 10, 101–111. [Google Scholar] [CrossRef]
- Rey Benayas, J.M.; Altamirano, A.; Miranda, A.; Catalán, G.P.; Prado, M.; Lisón, F.; Bullock, J.M. Landscape restoration in a mixed agricultural-forest catchment: Planning a buffer strip and hedgerow network in a Chilean biodiversity hotspot. Ambio 2020, 49, 310–323. [Google Scholar] [CrossRef] [Green Version]
- Rey Benayas, J.M.; Bullock, J.M.; Newton, A.C. Creating Woodland islets to reconcile ecological restoration, conservation, and agricultural land use. Front. Ecol. Environ. 2008, 6, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Decocq, G.; Andrieu, E.; Brunet, J.; Chabrerie, O.; De Frenne, P.; De Smedt, P.; Deconchat, M.; Diekmann, M.; Ehrmann, S.; Giffard, B.; et al. Ecosystem Services from Small Forest Patches in Agricultural Landscapes. Curr. For. Rep. 2016, 2, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Salako, V.K.; Kénou, C.; Dainou, K.; Assogbadjo, A.E.; Kakaï, R.G. Impacts of land use types on spatial patterns and neighbourhood distance of the agroforestry palm Borassus aethiopum Mart. in two climatic regions in Benin, West Africa. Agrofor. Syst. 2019, 93, 1057–1071. [Google Scholar] [CrossRef]
- Sun, D.; Yang, H.; Guan, D.; Yang, M.; Wu, J.; Yuan, F.; Jin, C.; Wang, A.; Zhang, Y. The effects of land use change on soil infiltration capacity in China: A meta-analysis. Sci. Total Environ. 2018, 626, 1394–1401. [Google Scholar] [CrossRef]
- Ordonez, J.; Luedeling, E.; Kindt, R.; Lestari Tata, H.; Harja, D.; Jamnadass, R.; Van Noordwijk, M. Tree diversity along the forest transition curve: Drivers, consequences and entry points for multifunctional agriculture. Curr. Opin. Environ. Sustain. 2014, 6, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [Green Version]
- Udawatta, R.; Rankoth, L.; Jose, S. Agroforestry and biodiversity. Sustainability 2019, 9, 2879. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Meacham, M.; Queiroz, C. A plea for multifunctional landscapes. Front. Ecol. Environ. 2017, 13, 59. [Google Scholar] [CrossRef]
- Hermuche, P.M.; Vieira, M.D.L.; Sano, E.E. Modeling tree cover changes in a pasture-dominated landscape by adopting silvopastoral practices in a dry forest region in Central Brazil. Agrofor. Syst. 2013, 87, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Holl, K.; Leighton Reid, J.; Cole, R.J.; Oviedo-Brenes, F.; Rosales, J.A.; Zahawi, R.A. Applied nucleation facilitates tropical forest recovery: Lessons learned from a 15-year study. J. Appl. Ecol. 2020, 57, 2316–2328. [Google Scholar] [CrossRef]
- Murekezi, J.P.; Nduwamungu, J.; Munyanziza, E. Investigation of survival rate of trees planted in agroforestry and forest plantations in Huye District from 2007 to 2011 and underlying factors. Rwanda J. Ser. E: Agric. Sci. 2013, 1, 52–61. [Google Scholar]
- Jaimez, R.E.; Araque, O.; Guzman, D.; Mora, A.; Espinoza, W.; Tezara, W. Agroforestry systems of timber species and cacao: Survival and growth during the early stages. J. Agric. Rural. Dev. Trop. Subtrop. 2013, 114, 1–11. [Google Scholar]
- Zermeño-Hernández, I.; Pingarroni, A.; Martínez-Ramos, M. Agricultural land-use diversity and forest regeneration potential in human- modified tropical landscapes. Agric. Ecosyst. Environ. 2016, 230, 210–220. [Google Scholar] [CrossRef]
- Chazdon1, R.L.; Lindenmayer, D.; Guariguata, M.R.; Crouzeilles, R.; Rey Benayas, J.M.; Chavero, E.L. Fostering natural forest regeneration on former agricultural land through economic and policy interventions. Environ. Res. Lett. 2021, 15, 043002. [Google Scholar] [CrossRef]
- Rey Benayas, J.M.; Bullock, J.M. Vegetation restoration and other actions to enhance wildlife in European agricultural landscapes. In Rewilding European Landscapes; Pereira, H.M., Navarro, L.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 127–142. [Google Scholar]
- Corbin, J.D.; Holl, K.D. Applied nucleation as a forest restoration strategy. For. Ecol. Manag. 2012, 265, 37–46. [Google Scholar] [CrossRef]
- World Wildlife Fund. Bosque Atlántic. 2019. Available online: http://www.wwf.org.py/dondetrabajamos/bosqueatlantico/ (accessed on 13 December 2019).
- Da Ponte, E.; Mack, B.; Wohlfart, C.; Rodas, O.; Fleckenstein, M.; Oppelt, N.; Dech, S.; Kuenzer, C. Assessing Forest Cover Dynamics and Forest Perception in the Atlantic Forest of Paraguay, Combining Remote Sensing and Household Level Data. Forests 2017, 8, 389. [Google Scholar] [CrossRef] [Green Version]
- Galeano, P. Calidad de suelo. In Sostenibilidad de la Biomasa Forestal Para Energía y del Etanol de Maíz y Caña de Azúcar en Paraguay; FAO: Rome, Italy, 2018; pp. 70–85. [Google Scholar]
- MAG (Ministerio de Agricultura y Ganadería). Síntesis Estadísticas Producción Agropecuaria, Año agrícola 2015/2016; MAG: San Lorenzo, Paraguay, 2016; p. 51. [Google Scholar]
- Marques, M.C.M.; Trindade, W.; Bohn, A.; Grelle, C.E.V. The Atlantic Forest: An Introduction to the Megadiverse Forest of South America. In The Atlantic Forest; Marques, M.C.M., Grelle, C.E.V., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Olson, D.M.; Dinerstein, E. The Global 200: Priority Ecoregions for Global Conservation. Ann. Mo. Bot. Gard. 2002, 89, 199–224. [Google Scholar] [CrossRef]
- Cartes, J.L. Brief history of conservation in the interior Atlantic forest. In The Atlantic Forest of South America: Biodiversity Status, Threats, and Outlook; Galindo-Leal, C., Camara, I.d.G., Eds.; Island Press: Washington, DC, USA, 2003; pp. 269–287. [Google Scholar]
- Huang, C.; Kim, S.; Altstatt, A.; Townshend, J.R.G.; Davis, P.; Song, K.; Tucker, C.J.; Rodas, O.; Yanosky, A.; Clay, R. Rapid Loss of Paraguay’s Atlantic Forest and status of protected areas—A Landsat assessment. Remote Sens. Environ. 2007, 106, 460–466. [Google Scholar] [CrossRef]
- Da Ponte, E.; Kuenzer, C.; Parker, A.; Rodas, O.; Oppelt, N.; Fleckenstein, M. Forest cover loss in Paraguay and perception of ecosystem services: A case study of the Upper Parana Forest. Ecosyst. Serv. 2017, 24, 200–212. [Google Scholar] [CrossRef] [Green Version]
- Reymondin, L.; Palacios, P.; Caballero, J.; Ortiz, E.; Báez, M.; Bautista, O.; Yanosky, A.; Villalba, A. Indirect Impact Assessment of the road segment San Juan Nepomuceno-Ruta 6 in Paraguay. Paraquaria Nat. 2016, 4, 20–32. [Google Scholar]
- World Wildlife Fund. Informe Cuatrimestral (Mayo a Agosto de 2016). In Monitoreo Satelital de la Deforestación; Desmontes detectados en el Bosque Atlántico: Región Oriental, Paraguay, 2016; Available online: http://d2ouvy59p0dg6k.cloudfront.net/downloads/reporte_deforestacion_wwf_mayago2016.pdf (accessed on 24 June 2020).
- Cartes, J.L.; Yanosky, A. Dinámicas de la pérdida de la biodiversidad en el Bosque Atlántico del Paraguay: Una Introducción. In El Bosque Atlántico en Paraguay, Biodiversidad, Amenazas y Perspectivas; Cartes, J.L., Ed.; State of Hotspost Serie: Asunción, Paraguay; Conservation International—Center for Applied Biodiversity Sciencie: Guyra, Paraguay, 2006; pp. 31–57. [Google Scholar]
- Montoya, D.; Haegeman, B.; Gaba, S.; Mazancourt, C.; Bretagnolle, V.; Loreau, M. Trade-offs in the provisioning and stability of ecosystem services in agroecosystems. Ecol. Appl. 2019, 29, 1853. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. International Society for Microbial Ecology. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tal, A. Making Conventional Agriculture Environmentally Friendly: Moving beyond the Glorification of Organic Agriculture and the Demonization of Conventional Agriculture. Sustainability 2018, 10, 1078. [Google Scholar] [CrossRef] [Green Version]
- Correia, J.E. Soy states: Resource politics, violent environments and soybean territorialization in Paraguay. J. Peasant. Stud. 2017, 46, 316–336. [Google Scholar] [CrossRef]
- Insfrán Ortiz, A.; Rey Benayas, J.M. La cultura de la restauración de los ecosistemas: Una tarea pendiente en sistemas tropicales y en el BAAPA en Paraguay. In Ecología Humana Contemporánea: Apuntes y Visiones en la Complejidad del Desarrollo; FCA-Universidad Nacional de Asunción: San Lorenzo, Paraguay, 2017; pp. 17–57. [Google Scholar]
- Altieri, M. Linking ecologists and traditional farmers in the search for sustainable agriculture. Front. Ecol. Environ. 2004, 2, 35–42. [Google Scholar] [CrossRef]
- Sims, B.; Kienzle, J. Sustainable agricultural mechanization for smallholders: What is it and how can we implement it? Agriculture 2017, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Mier Terán, M.; Giraldo, O.F.; Aldasoro, M.; Morales, H.; Ferguson, B.; Rosset, P.; Khadse, M.; Campos, A. Bringing agroecology to scale: Key drivers and emblematic cases. J. Agroecol. Sustain. Food Syst. 2018, 42, 637–665. [Google Scholar] [CrossRef]
- Rosset, P.M.; Altieri, M.A. Agroecology: Science and Politics; Fernwood Publishing: Winnipeg, MB, Canada, 2017; p. 160. [Google Scholar]
- Altieri, M.A.; Funes-Monzote, F.R.; Petersen, P. Agroecologically efficient agricultural systems for smallholder farmers: Contributions to food sovereignty. Agron. Sustain. Dev. 2012, 32, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Campanhola, C.; Pandey, S. (Eds.) Agroecological Approaches to Sustainable Intensification. In Sustainable Food and Agriculture, An Integrated Approach; Academic Press: Cambridge, MA, USA, 2019; pp. 179–184. [Google Scholar]
- Wezel, A.; Herren, B.G.; Kerr, R.B.; Barrios, E.; Rodrigues Gonçalves, A.L.; Sinclair, F. Agroecological principles and elements and their implications for transitioning to sustainable food systems. A review. Agron. Sustain. Dev. 2020, 40, 40. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Vieweg Verlag: New York, NY, USA, 2002. [Google Scholar]
- Dunn, K.P.; Smyth, G.K. Randomized quantile residuals. J. Comput. Graph. Stat. 1996, 5, 236–244. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, R version 4.1.0; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 18 December 2019).
- Bates, D.; Machler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2015, arXiv:1406.5823. Available online: http://arxiv.org/abs/1406.5823 (accessed on 20 December 2019).
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.43.6. 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 20 December 2019).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models; Technical Report Number 019, 2008; Department of Statistics University of Munich: München, Germany, 15 February 2008; p. 25. Available online: https://epub.ub.uni-muenchen.de/2120/1/tr019.pdf (accessed on 20 December 2019).
- Hartig, F. Package ‘DHARMa’. Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2019. Available online: http://florianhartig.github.io/DHARMa/ (accessed on 20 December 2019).
- Somarriba, E.; Beer, J. Productivity of Theobroma cacao agroforestry systems with timber of legume service shade trees. Agrofor. Syst. 2011, 81, 109–121. [Google Scholar] [CrossRef]
- Plath, M.; Mody, K.; Potvin, C.; Dorn, S. Establishment of native tropical timber trees in monoculture and mixed-species plantations: Small-scale effects on tree performance and insect herbivory. For. Ecol. Manag. 2011, 261, 741–750. [Google Scholar] [CrossRef]
- Salvadori, S.L.; Neves, C.U.; Duarte, B.D.; Gomes da Silva, A.F.; Lopes Klein, W. Análise de sobrevivência e crescimento de Cordia trichotoma, boraginaceae, lamiales, no sul de Mato Grosso Do Sul—Brasil. Ciência Florest. 2013, 23, 735–742. Available online: https://www.scielo.br/pdf/cflo/v23n4/1980-5098-cflo-23-04-00735.pdf (accessed on 24 January 2021). [CrossRef] [Green Version]
- Huntington, H.; Haflett, A.; Ewing, B. The impact of interventions to promote climate change adaptation: Does stronger tenure security increase farmer investment in sustainable agroforestry? In Proceedings of the 2018 World Bank Conference on Land and Poverty, Washington, DC, USA, 19–23 March 2018; pp. 19–23. [Google Scholar]
- Harmer, A. The effect of plant competition and simulated summer browsing by deer on tree regeneration. J. Appl. Ecol. 2001, 38, 1094–1103. [Google Scholar] [CrossRef]
- Bardgett, R. The Biology of Soil. In A Community and Ecosystem Approach; Oxford University Press: New York, NY, USA, 2005; p. 242. [Google Scholar]
- Chanatásig Vaca, C.I. Estudio Comparativo de la Macrofauna Edáfica en Huertos Caseros Y Plantaciones Monoespecíficas de Cedro (Cedrela odorata L.) en Tikinmul, Campeche; Tesis doctoral; ECOSUR: Campeche, Mexico, 2014; p. 158. [Google Scholar]
- Borda, A.; Nilda, E.; Vargas García, C. Evaluación de una Estrategia para el Control del Ataque de Hypsipyla grandella Zellar en una Plantación de Cedrela odorata L. (cedro rojo) y Swietenia macrophylla King (Caoba); en la provincia de Tambopata—Madre de Dios: Tambopata, Perú, 2019; Available online: http://repositorio.unamad.edu.pe/handle/UNAMAD/412 (accessed on 30 January 2021).
- Montero-Solís, F.M.; López-López, M.A.; Jiménez-Casas, M.; Cetina-Alcalá, V.M.; Platas-Rosado, D.E.; López-Collado, C.J. Gliricidia sepium and fertilization affect growth, nutrient status, and incidence of Hypsipyla grandella in a Cedrela odorata plantation. Agrofor. Syst. 2019, 93, 813–823. [Google Scholar] [CrossRef]
- Lisboa, D.O.; Silva, M.A.; Machado, A.R.; Pinho, D.B.; Borges, L.S.; Pereira, O.L.; Furtado, G.Q. First report of botryosphaeriaceous fungi causing canker on Cedrela fissilis and leaf spots on Cariniana estrellensis in forest nursery in Brazil. For. Pathol. 2016, 46, 362–365. [Google Scholar] [CrossRef]
- Pumariño, L.; Sileshi, G.W.; Gripenberg, S.; Kaartinen, R.; Barrios, E.; Muchane, M.N.; Midega, C.; Jonsson, M. Efectos de la agrosilvicultura en el control de plagas, enfermedades y malezas: Un metanálisis. Ecol. Básica Apl. 2015, 16, 573–582. [Google Scholar]
- Iverson, A.; Marin, L.E.; Ennis, K.K.; Gonthier, D.J.; Connor-Barrie, B.T.; Remfert, J.L. Review: Do polycultures promote win-wins or trade-offs in agricultural ecosystem services? A meta-analysis. J. Appl. Ecol. 2014, 51, 1593–1602. [Google Scholar] [CrossRef]
- Baggio, A.J.; Felizari, S.R.; Ruffato, A.; Soarez, A.O. Produção do Componente Arbóreo no Sistema Agroflorestal da Erva-Mate (Ilex Paraguariensis) em Machadinho, RS. Actas; V Congreso Sudamericano de la Yerba Mate: Posadas, Argentina, 2011; pp. 105–110. [Google Scholar]
- Pringle, E.; Dirzo, R.; Gordon, D. Plant defense, herbivory, and the growth of Cordia alliodora trees and their symbiotic Azteca ant colonies. Oecologia 2012, 170, 677–685. [Google Scholar] [CrossRef] [PubMed]
- De Castro, M.T.; Linhares Montalvão, S.C.; Gomes Monnerat, R. Breeding and biology of Hypsipyla grandella Zeller (Lepidoptera: Pyralidae) fed with mahogany seeds (Swietenia macrophylla King). J. Asia-Pac. Entomol. 2016, 19, 217–221. [Google Scholar] [CrossRef]
- Borges, R.; Boff, M.I.C.; Mantovani, A.; Biscaro-Borges, C.; Blassioli-Moraes, M.C. Effect of canopy cover on development of cedar (Cedrela fissilis) and aspects of damage caused by Hypsipyla grandella in agroforestry system. Ciência Florest. 2019, 29, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Pontes, D.M.F.; Vera Lex Engel, V.L.; Parrotta, J.A. Forest Structure, Wood Standing Stock, and Tree Biomass in Different Restoration Systems in the Brazilian Atlantic Forest. Forests 2019, 10, 588. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Shao, M.; Zhu, Y.; Luo, Y. Soil moisture decline due to afforestation across the Loess Plateau, China. J. Hydrol. 2017, 546, 113–122. [Google Scholar] [CrossRef]
- Badari, C.G.; Bernardini, L.E.; de Almeida, D.R.A.; Brancalion, P.H.S.; Ricardo Gomes César, R.; Gutierrez, V.; Chazdon, R.L.; Haroldo Borges Gomes, H.; Viani, R.A.G. Ecological outcomes of agroforests and restoration 15 years after planting. Restor. Ecol. 2020, 28, 1135–1144. [Google Scholar] [CrossRef]
- Lonrenzi, H. Brazilian Trees; Instituto Plantarum de Estudos da Flora: Nova Odessa, Brazil, 2002; Volume 1, p. 4. [Google Scholar]
- Bertolini, I.C.; Brun, E.J.; Debastiani, A.B. Caracterização silvicultural da canafístula (Peltophorum dubium (Sprengel) Taubert). Sci. Agrar. Parana. 2015, 14, 67–76. Available online: http://e-revista.unioeste.br/index.php/scientiaagraria/article/view/9842/8547 (accessed on 3 July 2019). [CrossRef]
- Gao, R.; Yang, X.; Yang, F.; Wei, L.; Huang, Z.; Walck, J.L. Aerial and soil seed banks enable populations of an anual species to cope with an unpredictable dune ecosystem. Ann. Bot. 2014, 114, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Silva, H.G.; José, A.C.; Rocha Faria, J.M.; Souza Pereira, W.V. Aspects of Peltophorum dubium Sprengel (Taubert) sedes in an aerial seed bank. J. Seed Sci. 2017, 39, 32–40. [Google Scholar] [CrossRef] [Green Version]
- CITES; UNEP. Apéndices I, II y III, Interpretación. CITES. 2017. Available online: https://cites.org/sites/default/files/esp/app/2017/S-Appendices-2017-10-04.pdf (accessed on 2 July 2019).
- Corea-Arias, E.; Arnáez-Serrano, E.; Moreira-González, E.; Castillo-Ugalde, M. Situación de nueve especies forestales en peligro crítico de extinción en Costa Rica. Rev. For. Mesoam. Kurú 2016, 13, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Rey Benayas, J.M. Restauración de campos agrícolas sin competir por el uso de la tierra para aumentar su biodiversidad y servicios ecosistémicos. Investig. Ambient. 2012, 4, 101–110. [Google Scholar]
- Bayala, J.; Sanou, J.; Bazie, H.R.; Coe, R.; Kalinganire, A.; Sinclair, F.L. Regenerated trees in farmers’ fields increase soil carbon across the Sahel. Agrofor. Syst. 2019, 94, 401–415. [Google Scholar] [CrossRef] [Green Version]
- Häger, A.; Fernández Otárola, M.; Stuhlmacher, M.F.; Acuña Castillo, R.; Contreras Arias, A. Effects of management and landscape composition on the diversity and structure of tree species assemblages in coffee agro forests. Agric. Ecosyst. Environ. 2015, 199, 43–51. [Google Scholar] [CrossRef]
- Bukomeko, H.; Jassogne, L.; Tumwebaze, S.B.; Eilu, G.; Vaast, P. Integrating local knowledge with tree diversity analyses to optimize on-farm tree species composition for ecosystem service delivery in coffee agroforestry systems of Uganda. Agrofor. Syst. 2019, 93, 755–777. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.L. Landscape Restoration, Natural Regeneration, and the Forests of the Future. Ann. Mo. Bot. Gard. 2017, 102, 251–257. [Google Scholar] [CrossRef]
- Holl, K.D.; Reid, L.; Chaves-Fallas, J.M.; Oviedo-Brenes, F.; Zahawi, R.A. Local Tropical Forest Restoration Strategies Affect Tree Recruitment More Strongly than Does Landscape Forest Cover. J. Appl. Ecol. 2017, 54, 1091–1099. [Google Scholar] [CrossRef]
- Reilly, K.; Cullen, E.; Lola-Luz, T.; Stone, D.; Valverde, J.; Gaffney, M.; Brunton, N.; Grant, J.; Griffiths, B.S. Effect of organic, conventional and mixed cultivation practices on soil microbial community structure and nematode abundance in a cultivated onion crop. J. Sci. Food Agric. 2013, 93, 3700–3709. [Google Scholar] [CrossRef] [Green Version]
- Lupatini, M.; Korthals, G.W.; Hollander, M.; Janssens, T.K.S.; Kuramae, E.E. Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 2017, 7, 2064. [Google Scholar] [CrossRef] [Green Version]
- Clasen, C.; Heurich, M.; Glaesener, L.; Kennel, E.; Knoke, T. What factors affect the survival of tree saplings under browsing, and how can a loss of admixed tree species be forecast? Ecol. Model. 2015, 305, 1–9. [Google Scholar] [CrossRef]
- Schapovaloff, C.; Daría, A. Efecto del Glifosato en el Suelo y Plantas Nativas—Exóticas. Master’s Thesis, Universidad Nacional de Itapúa, Encarnación, Paraguay, 2013. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Agronomic System | Plantation Type | No. Plots | Plot Size (m2) | Tree Density (Individuals ha−1) | Monitoring Period (Years) * |
|---|---|---|---|---|---|---|
| Caaguazú | Conventional | Pathside | 3 | 100 | 1800 | 2010–2016 |
| Field edge | 3 | 100 | 1800 | 2010–2016 | ||
| Islet | 3 | 100 | 1800 | 2010–2016 | ||
| Agroecological | Pathside | 3 | 100 | 1800 | 2010–2016 | |
| Field edge | 3 | 100 | 1800 | 2010–2016 | ||
| Islet | 3 | 100 | 1800 | 2010–2016 | ||
| Total | 18 | |||||
| Itapúa | Conventional | Islet | 10 | 50 | 1600 | 2012–2016 |
| Traditional | Islet | 10 | 50 | 1600 | 2012–2016 | |
| Agroecological | Islet | 10 | 50 | 1600 | 2012–2016 | |
| Total | 30 |
| Site | Model | (Intercept) | AICc | Delta | Weight | R2m | R2c |
|---|---|---|---|---|---|---|---|
| Caaguazú | 512 | 3,590,128 | 959.8110 | 0.0000000 | 269,608.5 | 0.8687040 | 0.8748712 |
| 1024 | 3,750,841 | 960.4830 | 0.6720045 | 192,668.1 | 0.8671280 | 0.8733760 | |
| 384 | 3,656,629 | 961.1081 | 1.2971396 | 140,949.4 | 0.8633484 | 0.8693383 | |
| Itapúa | 24 | 0.5699077 | 556.5128 | 0.000000 | 863,392.6 | 0.2894882 | 0.2924227 |
| Site | Model | (Intercept) | AICc | Delta | Weight | R2m | R2c |
|---|---|---|---|---|---|---|---|
| Caaguazú | 40 | 6,978,656 | 298.5042 | 0.000000 | 946,708.8 | 0.8671280 | 0.8733760 |
| Itapúa | 2 | 0.2708572 | 638.4951 | 0.000000 | 888,152.8 | 0.2894882 | 0.2924227 |
| Site | Model | (Intercept) | AICc | Delta | Weight |
|---|---|---|---|---|---|
| Caaguazú | 22 | −193,025,850 | 159.8007 | 0.000000 | 676,183.7 |
| 24 | −200,038,011 | 161.6283 | 1.827547 | 271,155.1 | |
| Itapúa | 4 | −18.560 | 130.8007 | 0.000000 | 676,183.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Insfrán Ortiz, A.; Rey Benayas, J.M.; Cayuela, L. Establishment and Natural Regeneration of Native Trees in Agroforestry Systems in the Paraguayan Atlantic Forest. Forests 2022, 13, 2045. https://doi.org/10.3390/f13122045
Insfrán Ortiz A, Rey Benayas JM, Cayuela L. Establishment and Natural Regeneration of Native Trees in Agroforestry Systems in the Paraguayan Atlantic Forest. Forests. 2022; 13(12):2045. https://doi.org/10.3390/f13122045
Chicago/Turabian StyleInsfrán Ortiz, Amado, José María Rey Benayas, and Luis Cayuela. 2022. "Establishment and Natural Regeneration of Native Trees in Agroforestry Systems in the Paraguayan Atlantic Forest" Forests 13, no. 12: 2045. https://doi.org/10.3390/f13122045
APA StyleInsfrán Ortiz, A., Rey Benayas, J. M., & Cayuela, L. (2022). Establishment and Natural Regeneration of Native Trees in Agroforestry Systems in the Paraguayan Atlantic Forest. Forests, 13(12), 2045. https://doi.org/10.3390/f13122045