
Testing the Efficiency of the Push-and-Pull Strategy during Severe Ips typographus Outbreak and Extreme Drought in Norway Spruce Stands

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study Area
2.2. Dispensers
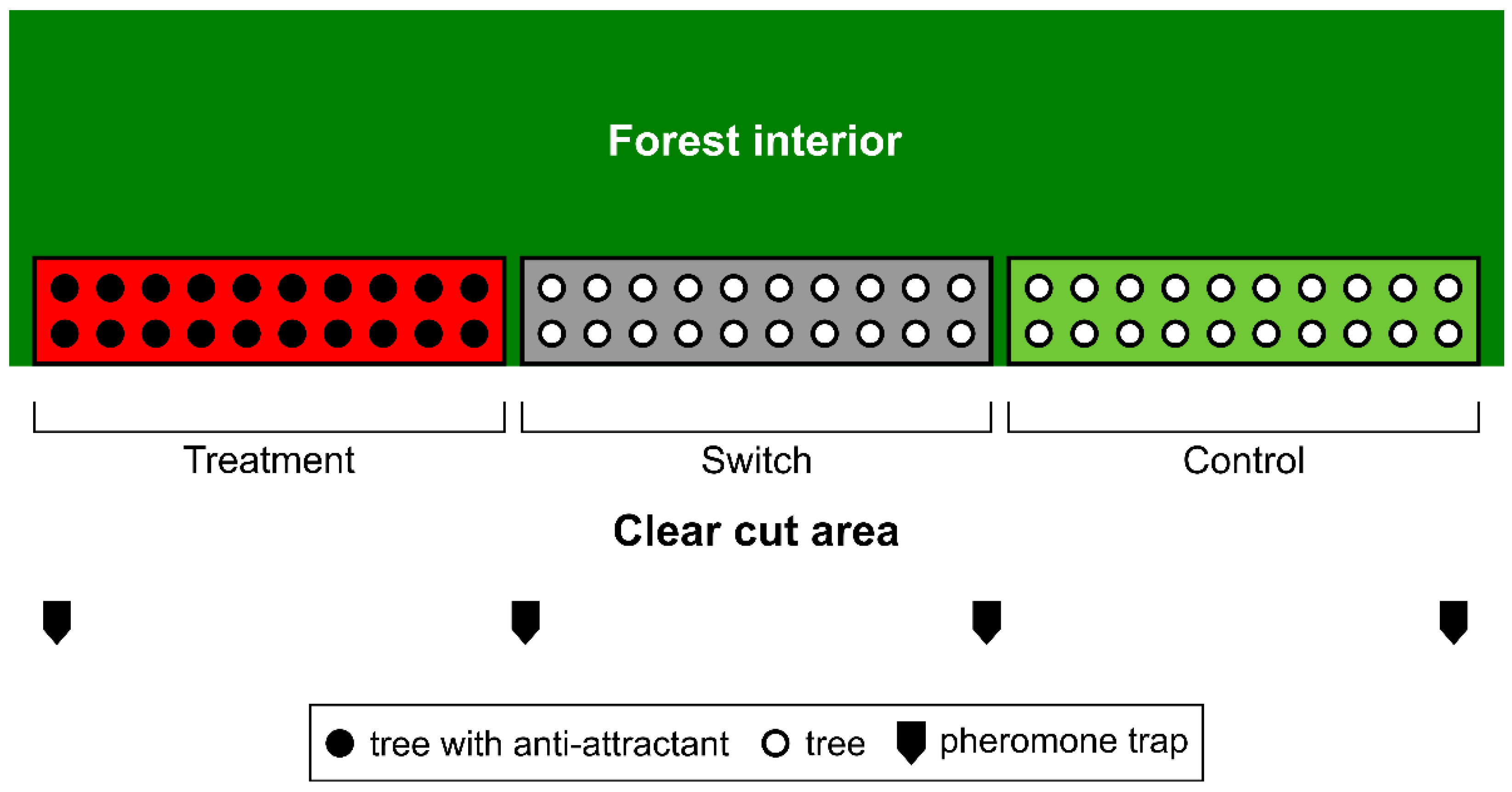
2.3. Experimental Design
2.4. Statistical Analyses
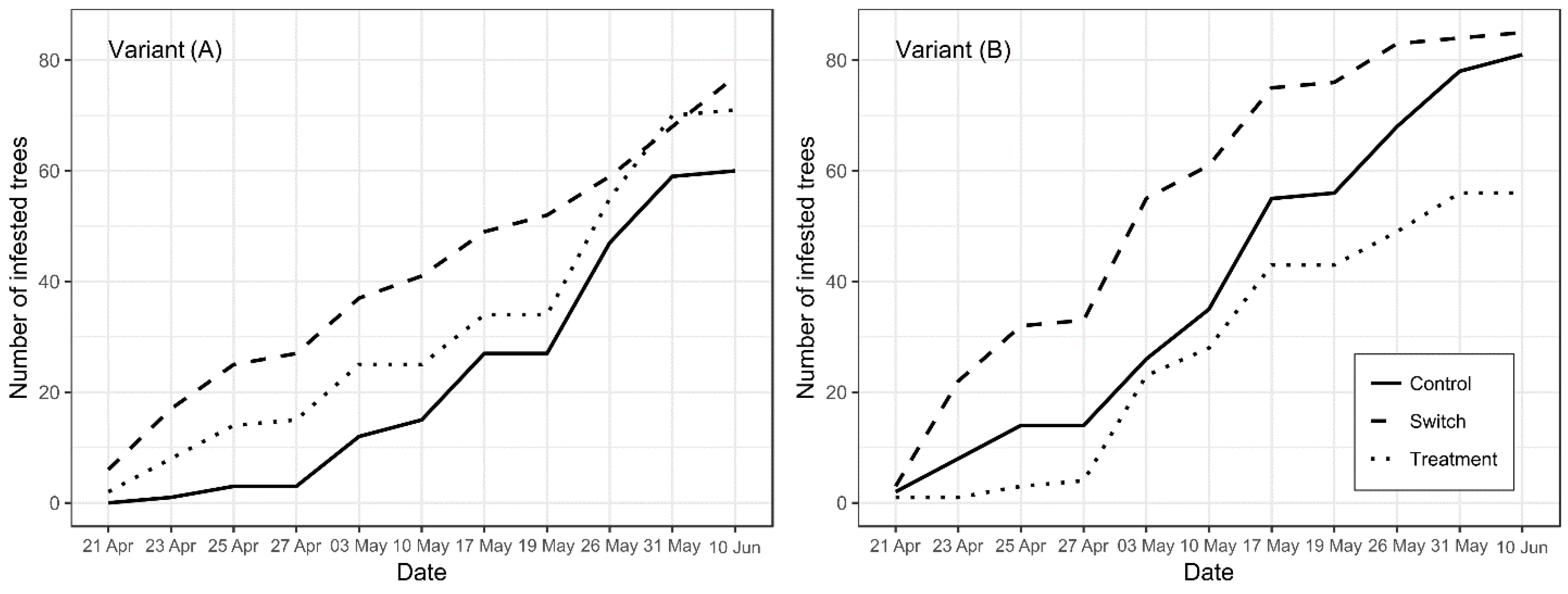
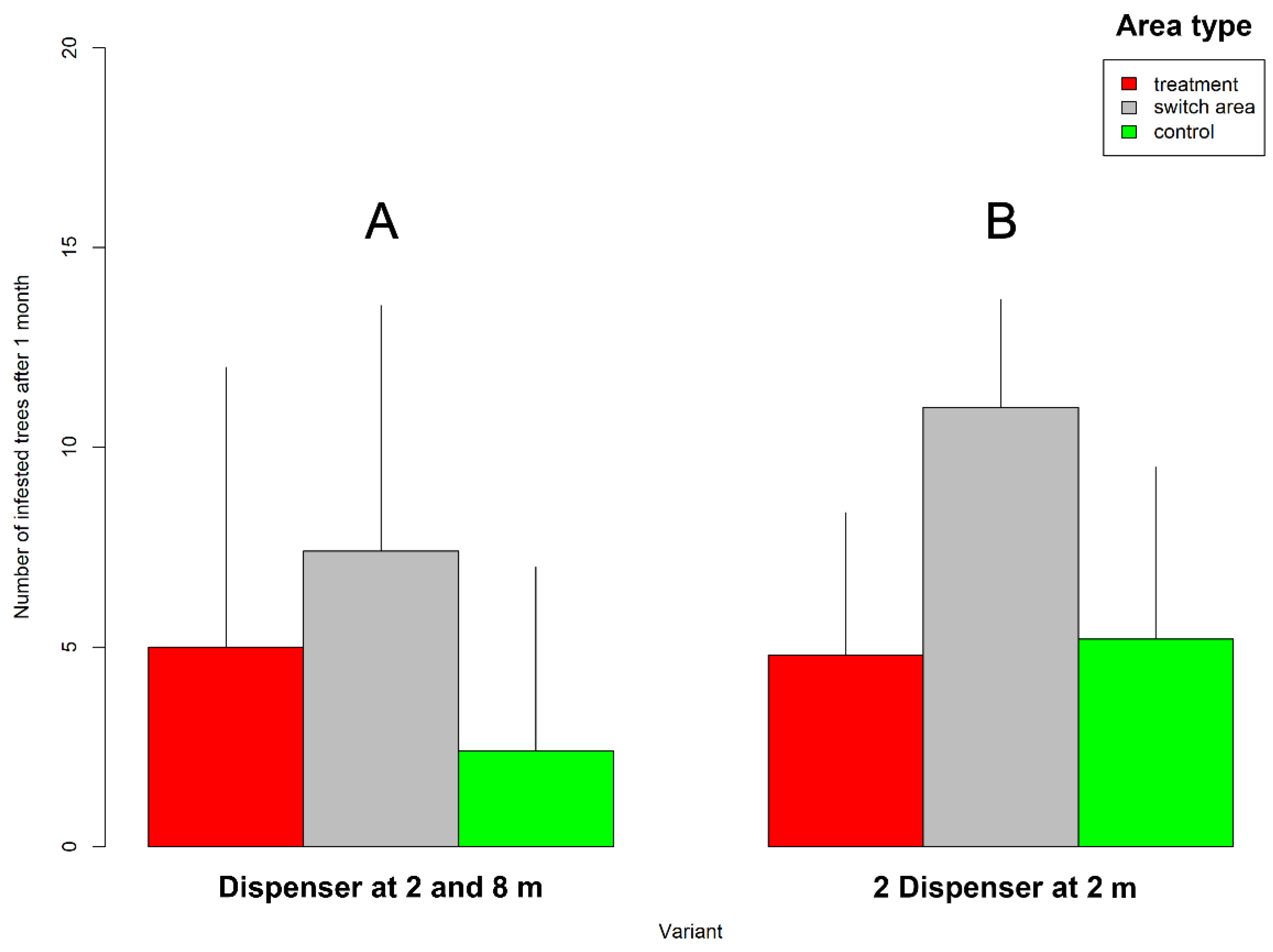
3. Results
4. Discussion
4.1. Experimental Results and “Switch” Effect
4.2. Effect of Drought and High Bark Beetle Population
4.3. Effect of Pheromone Traps and Push-Pull System
4.4. Limitations of the Study
4.5. Research Needs
- Optimize the spatial layout by not placing anti-attractants on all trees and only applying them at lower height;
- Optimize the area of anti-attractant application by using spatial tools (tree health diagnostics, remote sensing, and GIS) and methods on the scale from sniffer dogs [62] to UAVs to satellites;
- Test the possibility of increasing the effectiveness of anti-attractants in summer conditions;
- Test the possibility of using anti-attractants in areas with intensive use of mass trapping or pheromone trap barriers (push-pull effect) and in areas without them (only anti-attractants).
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toth, D.; Maitah, M.; Maitah, K.; Jarolínová, V. The Impacts of Calamity Logging on the Development of Spruce Wood Prices in Czech Forestry. Forests 2020, 11, 283. [Google Scholar] [CrossRef] [Green Version]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating Outbreak of Bark Beetles in the Czech Republic: Drivers, Impacts, and Management Implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Gonzalez, R.; Grégoire, J.-C.; Drumont, A.; De Windt, N. A Sampling Technique to Estimate Within-Tree Populations of Pre-Emergent Ips typographus (Col., Scolytidae). J. Appl. Entomol. 1996, 120, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Kautz, M.; Schopf, R.; Ohser, J. The “Sun-Effect”: Microclimatic Alterations Predispose Forest Edges to Bark Beetle Infestations. Eur. J. For. Res. 2013, 132, 453–465. [Google Scholar] [CrossRef]
- Doležal, P.; Sehnal, F. Effects of Photoperiod and Temperature on the Development and Diapause of the Bark Beetle Ips typographus. J. Appl. Entomol. 2007, 131, 165–173. [Google Scholar] [CrossRef]
- Wermelinger, B. Ecology and Management of the Spruce Bark Beetle Ips typographus—A Review of Recent Research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Jakuš, R. A Method for the Protection of Spruce Stands Against Ips typographus by the Use of Barriers of Pheromone Traps in North-Eastern Slovakia. Anz. Für Schädlingskunde Pflanzenschutz Umweltschutz 1998, 71, 152–158. [Google Scholar] [CrossRef]
- Bakke, A.; Sæther, T.; Kvamme, T. Mass Trapping of the Spruce Bark Beetle Ips typographus: Pheromone and Trap Technology; Norsk Institutt for Skogforskning: Ås, Norway, 1983; ISBN 978-82-7169-299-5. [Google Scholar]
- Vité, J.P. The European Struggle to Control Ips typographus—Past, Present and Future. Ecography 1989, 12, 520–525. [Google Scholar] [CrossRef]
- Weslien, J. Effects of Mass Trapping on Ips typographus (L.) Populations. J. Appl. Entomol. 1992, 114, 228–232. [Google Scholar] [CrossRef]
- Niemeyer, H. Integrated Bark Beetle Control: Experiences and Problems in Northern Germany. In Proceedings: Integrating Cultural Tactics into the Management of Bark Beetle and Reforestation Pests; Vallombrosa, Italy, 1–3 September 1996; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1997; pp. 80–86. [Google Scholar]
- Faccoli, M.; Stergulc, F. Damage Reduction and Performance of Mass Trapping Devices for Forest Protection against the Spruce Bark Beetle, Ips typographus (Coleoptera Curculionidae Scolytinae). Ann. For. Sci. 2008, 65, 309. [Google Scholar] [CrossRef]
- Zahradník, P.; Zahradníková, M. The Efficacy of a New Pheromone Trap Setup Design, Aimed for Trapping Ips typographus (Coleoptera, Curculionidae, Scolytinae). Šumar. List 2015, 139, 185–186. [Google Scholar]
- Vité, J.P. Erfahrungen und Erkenntnisse zur akuten Gefahrdung des mitteleuropaischen Fichtenwaldes durch Kaferbefall. Allg. Forstz. 1984, 29, 249–254. [Google Scholar]
- Ross, D.W.; Daterman, G.E. Efficacy of an Antiaggregation Pheromone for Reducing Douglas-Fir Beetle, Dendroctonus pseudotsugae Hopkins (Coleoptera: Scolytidae), Infestation in High Risk Stands. Can. Entomol. 1995, 127, 805–811. [Google Scholar] [CrossRef]
- Schlyter, F. Semiochemical Diversity in Practice: Antiattractant Semiochemicals Reduce Bark Beetle Attacks on Standing Trees—A First Meta-Analysis. Psyche J. Entomol. 2012, 2012, e268621. [Google Scholar] [CrossRef] [Green Version]
- Seybold, S.J.; Bentz, B.J.; Fettig, C.J.; Lundquist, J.E.; Progar, R.A.; Gillette, N.E. Management of Western North American Bark Beetles with Semiochemicals. Annu. Rev. Entomol. 2018, 63, 407–432. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.W.; Daterman, G.E. Reduction of Douglas-Fir Beetle Infestation of High-Risk Stands by Antiaggregation and Aggregation Pheromones. Can. J. For. Res. 1994, 24, 2184–2190. [Google Scholar] [CrossRef]
- Borden, J.H.; Birmingham, A.L.; Burleigh, J.S. Evaluation of the Push-Pull Tactic against the Mountain Pine Beetle Using Verbenone and Non-Host Volatiles in Combination with Pheromone-Baited Trees. For. Chron. 2006, 82, 579–590. [Google Scholar] [CrossRef]
- Gillette, N.E.; Mehmel, C.J.; Mori, S.R.; Webster, J.N.; Wood, D.L.; Erbilgin, N.; Owen, D.R. The Push-Pull Tactic for Mitigation of Mountain Pine Beetle (Coleoptera: Curculionidae) Damage in Lodgepole and Whitebark Pines. Environ. Entomol. 2012, 41, 1575–1586. [Google Scholar] [CrossRef] [Green Version]
- Schiebe, C.; Blaženec, M.; Jakuš, R.; Unelius, C.R.; Schlyter, F. Semiochemical Diversity Diverts Bark Beetle Attacks from Norway Spruce Edges. J. Appl. Entomol. 2011, 135, 726–737. [Google Scholar] [CrossRef]
- Birgersson, G.; Leufvén, A. The Influence of Host Tree Response to Ips typographus and Fungal Attack on Production of Semiochemicals. Insect Biochem. 1988, 18, 761–770. [Google Scholar] [CrossRef]
- Zhang, Q.-H.; Schlyter, F. Olfactory Recognition and Behavioural Avoidance of Angiosperm Nonhost Volatiles by Conifer-Inhabiting Bark Beetles. Agric. For. Entomol. 2004, 6, 1–20. [Google Scholar] [CrossRef]
- Zhang, Q.-H.; Birgersson, G.; Zhu, J.; Löfstedt, C.; Löfqvist, J.; Schlyter, F. Leaf Volatiles from Nonhost Deciduous Trees: Variation by Tree Species, Season and Temperature, and Electrophysiological Activity in Ips typographus. J. Chem. Ecol. 1999, 25, 1923–1943. [Google Scholar] [CrossRef]
- Zhang, Q.-H.; Schlyter, F. Redundancy, Synergism, and Active Inhibitory Range of Non-Host Volatiles in Reducing Pheromone Attraction in European Spruce Bark Beetle Ips typographus. Oikos 2003, 101, 299–310. [Google Scholar] [CrossRef]
- Andersson, M.N.; Larsson, M.C.; Blazenec, M.; Jakus, R.; Zhang, Q.-H.; Schlyter, F. Peripheral Modulation of Pheromone Response by Inhibitory Host Compound in a Beetle. J. Exp. Biol. 2010, 213, 3332–3339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binyameen, M.; Jankuvová, J.; Blaženec, M.; Jakuš, R.; Song, L.; Schlyter, F.; Andersson, M.N. Co-Localization of Insect Olfactory Sensory Cells Improves the Discrimination of Closely Separated Odour Sources. Funct. Ecol. 2014, 28, 1216–1223. [Google Scholar] [CrossRef]
- Kalinová, B.; Břízová, R.; Knížek, M.; Turčáni, M.; Hoskovec, M. Volatiles from Spruce Trap-Trees Detected by Ips typographus Bark Beetles: Chemical and Electrophysiological Analyses. Arthropod-Plant Interact. 2014, 8, 305–316. [Google Scholar] [CrossRef]
- Blažytė-Čereškienė, L.; Apšegaitė, V.; Radžiutė, S.; Mozūraitis, R.; Būda, V.; Pečiulytė, D. Electrophysiological and Behavioural Responses of Ips typographus (L.) to Trans-4-Thujanol—A Host Tree Volatile Compound. Ann. For. Sci. 2016, 73, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Schiebe, C.; Unelius, C.R.; Ganji, S.; Binyameen, M.; Birgersson, G.; Schlyter, F. Styrene, (+)-Trans-(1R,4S,5S)-4-Thujanol and Oxygenated Monoterpenes Related to Host Stress Elicit Strong Electrophysiological Responses in the Bark Beetle Ips typographus. J. Chem. Ecol. 2019, 45, 474–489. [Google Scholar] [CrossRef] [Green Version]
- Jirošová, A.; Kalinová, B.; Modlinger, R.; Jakuš, R.; Unelius, C.R.; Blaženec, M.; Schlyter, F. Anti-Attractant Activity of (+)-Trans-4-Thujanol for Eurasian Spruce Bark Beetle Ips typographus: Novel Potency for Females. Pest Manag. Sci. 2022, 78, 1992–1999. [Google Scholar] [CrossRef]
- Miller, J.; Siegert, P.; Amimo, F.; Walker, E. Designation of chemicals in terms of the locomotor responses they elicit from insects: An update of Dethier et al. (1960). J. Econ. Entomol. 2009, 102, 2056–2060. [Google Scholar] [CrossRef] [Green Version]
- Jakuš, R.; Dudová, A. Experimental Use of Aggregation and Anti-Aggregation Pheromones against the Spruce Bark Beetle (Ips typographus) in Decaying Spruce Stands with a Lower Level of Canopy Closure. J. For. Sci. 1999, 45, 525–531. [Google Scholar]
- Jakuš, R.; Schlyter, F.; Zhang, Q.-H.; Blaženec, M.; Vaverčák, R.; Grodzki, W.; Brutovský, D.; Lajzová, E.; Turčáni, M.; Bengtsson, M.; et al. Overview of Development of an Anti-Attractant Based Technology for Spruce Protection against Ips typographus: From Past Failures to Future Success. Anz. Für Schädlingskunde 2003, 76, 89–99. [Google Scholar] [CrossRef]
- Jakuš, R.; Blaženec, M.; Vojtěch, O. Use of anti-attractants in specific conditions of protected areas. Folia Oecol. 2011, 38, 46–51. [Google Scholar]
- Vojenský Újezd Libavá. Available online: https://www.vojujezd-libava.cz/vismo/dokumenty2.asp?u=9342&id_org=9342&id=3381 (accessed on 11 December 2022).
- Tolasz, R.; Míková, T.; Valerıánová, A.; Voženílek, V. Climate Atlas of Czechia, 1st ed.; Czecia Český Hydrometeoro-Logický Ústav, Univerzita Palackého Praha: Olomouc, Czech Republic, 2007; p. 255. ISBN 978-80-86690-26-1. [Google Scholar]
- Buras, A.; Rammig, A.; Zang, C.S. Quantifying Impacts of the 2018 Drought on European Ecosystems in Comparison to 2003. Biogeosciences 2020, 17, 1655–1672. [Google Scholar] [CrossRef]
- Wichmann, L.; Ravn, H.P. The Spread of Ips typographus (L.) (Coleoptera, Scolytidae) Attacks Following Heavy Windthrow in Denmark, Analysed Using GIS. For. Ecol. Manag. 2001, 148, 31–39. [Google Scholar] [CrossRef]
- Pekár, S.; Brabec, M. Moderní Analýza Biologických Dat; MUNI Press: Brno, Czech Republic, 2012; ISBN 978-80-210-5812-5. [Google Scholar]
- Schielzeth, H.; Nakagawa, S. Nested by Design: Model Fitting and Interpretation in a Mixed Model Era. Methods Ecol. Evol. 2013, 4, 14–24. [Google Scholar] [CrossRef]
- Pekár, S.; Brabec, M. Modern Analysis of Biological Data; MUNI Press: Brno, Czech Republic, 2016; ISBN 978-80-210-8019-5. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2022. Available online: https://www.r-project.org/ (accessed on 11 December 2022).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; Van Willigen, B.; Ranke, J.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. 2022. Available online: https://cran.r-project.org/web/packages/nlme/nlme.pdf (accessed on 11 December 2022).
- Maher, J.M.; Markey, J.C.; Ebert-May, D. The Other Half of the Story: Effect Size Analysis in Quantitative Research. CBE—Life Sci. Educ. 2013, 12, 345–351. [Google Scholar] [CrossRef]
- Nakagawa, S.; Cuthill, I.C. Effect Size, Confidence Interval and Statistical Significance: A Practical Guide for Biologists. Biol. Rev. 2007, 82, 591–605. [Google Scholar] [CrossRef]
- Fettig, C.J.; Munson, A.S.; Reinke, M.; Mafra-Neto, A. A Novel Semiochemical Tool for Protecting Pinus contorta from Mortality Attributed to Dendroctonus ponderosae (Coleoptera: Curculionidae). J. Econ. Entomol. 2015, 108, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Hansen, E.M.; Munson, A.S.; Blackford, D.C.; Graves, A.D.; Coleman, T.W.; Baggett, L.S. 3-Methylcyclohex-2-En-1-One for Area and Individual Tree Protection against Spruce Beetle (Coleoptera: Curculionidae: Scolytinae) Attack in the Southern Rocky Mountains. J. Econ. Entomol. 2017, 110, 2140–2148. [Google Scholar] [CrossRef] [Green Version]
- Foote, G.G.; Fettig, C.J.; Ross, D.W.; Runyon, J.B.; Coleman, T.W.; Gaylord, M.L.; Graves, A.D.; McMillin, J.D.; Mortenson, L.A.; Mafra-Neto, A. A Biodegradable Formulation of MCH (3-Methylcyclohex-2-En-1-One) for Protecting Pseudotsuga menziesii from Dendroctonus pseudotsugae (Coleoptera: Curculionidae) Colonization. J. Econ. Entomol. 2020, 113, 1858–1863. [Google Scholar] [CrossRef] [PubMed]
- Gillette, N.E.; Mehmel, C.J.; Webster, J.N.; Mori, S.R.; Erbilgin, N.; Wood, D.L.; Stein, J.D. Aerially Applied Methylcyclohexenone-Releasing Flakes Protect Pseudotsuga Menziesii Stands from Attack by Dendroctonus pseudotsugae. For. Ecol. Manag. 2009, 257, 1231–1236. [Google Scholar] [CrossRef]
- Gillette, N.E.; Erbilgin, N.; Webster, J.N.; Pederson, L.; Mori, S.R.; Stein, J.D.; Owen, D.R.; Bischel, K.M.; Wood, D.L. Aerially Applied Verbenone-Releasing Laminated Flakes Protect Pinus contorta Stands from Attack by Dendroctonus ponderosae in California and Idaho. For. Ecol. Manag. 2009, 257, 1405–1412. [Google Scholar] [CrossRef]
- Weslien, J.; Annila, E.; Bakke, A.; Bejer, B.; Eidmann, H.H.; Narvestad, K.; Nikula, A.; Ravn, H.P. Estimating Risks for Spruce Bark Beetle (Ips typographus (L.)) Damage Using Pheromone-baited Traps and Trees. Scand. J. For. Res. 1989, 4, 87–98. [Google Scholar] [CrossRef]
- Faccoli, M.; Stergulc, F. Ips typographus (L.) Pheromone Trapping in South Alps: Spring Catches Determine Damage Thresholds. J. Appl. Entomol. 2004, 128, 307–311. [Google Scholar] [CrossRef]
- Weslien, J. Monitoring Ips typographus (L.) Populations and Forecasting Damage. J. Appl. Entomol. 1992, 114, 338–340. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The Use of Push-Pull Strategies in Integrated Pest Management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, B.S.; Borden, J.H. Displacement and Aggregation of Mountain Pine Beetles, Dendroctonus ponderosae (Coleoptera: Scolytidae), in Response to Their Antiaggregation and Aggregation Pheromones. Can. J. For. Res. 1993, 23, 286–290. [Google Scholar] [CrossRef]
- Vandygriff, J.C.; Rasmussen, L.A.; Rineholt, J.F. A Novel Approach to Managing Fuelwood Harvest Using Bark Beetle Pheromones. West. J. Appl. For. 2000, 15, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Borden, J.H.; Chong, L.J.; Earle, T.J.; Huber, D.P.W. Protection of Lodgepole Pine from Attack by the Mountain Pine Beetle, Dendroctonus ponderosae (Coleoptera: Scolytidae) Using High Doses of Verbenone in Combination with Nonhost Bark Volatiles. For. Chron. 2003, 79, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Perkins, D.L.; Jorgensen, C.L.; Rinella, M.J. Verbenone Decreases Whitebark Pine Mortality Throughout a Mountain Pine Beetle Outbreak. For. Sci. 2015, 61, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Raffa, K.F.; Andersson, M.N.; Schlyter, F. Chapter One—Host Selection by Bark Beetles: Playing the Odds in a High-Stakes Game. In Advances in Insect Physiology; Beetles, P.B., Tittiger, C., Blomquist, G.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 50, pp. 1–74. [Google Scholar]
- Fettig, C. Efficacy of SPLAT® Verb for Protecting Individual Pinus contorta, Pinus ponderosa, and Pinus lambertiana from Mortality Attributed to Dendroctonus ponderosae. J. Entomol. Soc. Br. Columbia 2016, 113, 11–20. [Google Scholar]
- Vošvrdová, N.; Johansson, A.; Turčáni, M.; Jakuš, R.; Tyšer, D.; Schlyter, F.; Modlinger, R. Dogs trained to recognise a bark beetle pheromone locate recently attacked spruces better than human experts. For. Ecol. Manag. 2023, 528, 120626. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Block | Age | % of Spruce | Altitude (m) | Aspect | Volume of Bark-Beetle-Attacked Trees (m3/ha) in Surrounding of Experimental Plots | |||
|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 * | 2019 | |||||
| 1 | 59 | 95 | 640 | E | 11 | 60 | 313 | 2 |
| 2 | 83 | 98 | 620 | S | 38 | 73 | 213 | 0 |
| 3 | 79 | 92 | 640 | SE | 28 | 130 | 103 | 26 |
| 4 | 75 | 95 | 640 | W | 33 | 49 | 215 | 27 |
| 5 | 71 | 100 | 610 | S | 20 | 24 | 78 | 1 |
| 6 | 85 | 100 | 605 | S | 62 | 61 | 106 | 104 |
| 7 | 74 | 100 | 605 | S | 67 | 44 | 88 | 0 |
| 8 | 68 | 50 | 620 | S | 38 | 24 | 136 | 0 |
| 9 | 68 | 50 | 620 | S | 34 | 30 | 255 | 25 |
| 10 | 83 | 98 | 620 | SE | 38 | 73 | 213 | 0 |
| Mean | 74.5 | 87.8 | 622 | - | 37 | 57 | 172 | 19 |
| Factors | Number of df | Denominator df | F-Value | p-Value | |
|---|---|---|---|---|---|
| Intercept | 1 | 16 | 36.3 | <0.0001 | *** |
| Variant 1 | 1 | 8 | 1.01 | 0.328 | |
| Expt Area 2 | 2 | 16 | 4.20 | 0.034 | * |
| Variant × Expt Area 3 | 2 | 16 | 0.52 | 0.6086 |
| Trmt Nr | Test Data (Treatments) 2 | Test Mean | Test SD | Test n | Control Mean | Control SD | Control n | ∆ Mean | ES * (d) | Species 1 (Plot Area, ha) | Study |
|---|---|---|---|---|---|---|---|---|---|---|---|
| “A” = ‘1 + 8 m’ | |||||||||||
| 1 | A Trmt vs. Ctrl | 5.0 | 7.0 | 5 | 2.4 | 3.3 | 5 | 2.6 | 0.5 | ITYP | This study |
| 2 | A Switch vs. Ctrl | 7.4 | 6.2 | 5 | 2.4 | 3.3 | 5 | 5.0 | 1.1 | ITYP | This study |
| “B” = ‘2 × 1 m’ | |||||||||||
| 3 | B Trmt vs. Ctrl | 4.8 | 3.6 | 5 | 5.2 | 4.3 | 5 | −0.4 | −0.1 | ITYP | This study |
| 4 | B Switch vs. Ctrl | 11.0 | 2.7 | 5 | 5.2 | 4.3 | 5 | 5.8 | 1.6 | ITYP | This study |
| 5 | Mean of 32 studies (Trmt vs. Ctrl) | −1.0 | ITYP and MPB | [16] | |||||||
| 6 | SPLAT Vn | 1.5 | 2.1 | 5 | 7.9 | 5.5 | 5 | −6.4 | −1.7 | MPB (0.4) | [47] |
| 7 | Vn 7g pouch | 1.9 | 2.0 | 5 | 7.9 | 5.5 | 5 | −6.0 | −1.6 | MPB (0.4) | [47] |
| 8 | MCH alone | 2.2 | 0.8 | 3 | −0.4 | SB (0.6) | [48] | ||||
| 9 | MCH | 53% | 31% | 3 | 97% | 5.2% | 3 | −44% | −2.4 | SB | [48] |
| 10 | AKB | 47% | 31% | 3 | 97% | 5.2% | 3 | −50% | −2.8 | SB | [48] |
| 11 | MCH&AKB | 0% | 0% | 3 | 97% | 5.2% | 3 | −97% | −37 | SB | [48] |
| 12 | SPLAT MCH | 0.09 | 0.25 | 10 | 0.59 | 0.35 | 10 | −0.50 | −1.7 | DFB (0.04) | [49] |
| 13 | MCH Bubble Cap | 0.16 | 0.29 | 10 | 0.59 | 0.35 | 10 | −0.43 | −1.3 | DFB (0.04) | [49] |
| 14 | SPLAT MCH | 29.3% | 7.1% | 6 | 45.6% | 19.1% | 6 | −16% | −1.2 | DFB (0.4) | [49] |
| 15 | MCH Bubble Cap | 18.6% | 9.3% | 6 | 45.6% | 19.1% | 6 | −27% | −1.9 | DFB (0.4) | [49] |
| 16 | Vn Plus | 2.8 | 2.0 | 6 | 12.7 | 8.7 | 6 | −9.9 | −1.9 | WPB | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakuš, R.; Modlinger, R.; Kašpar, J.; Majdák, A.; Blaženec, M.; Korolyova, N.; Jirošová, A.; Schlyter, F. Testing the Efficiency of the Push-and-Pull Strategy during Severe Ips typographus Outbreak and Extreme Drought in Norway Spruce Stands. Forests 2022, 13, 2175. https://doi.org/10.3390/f13122175
Jakuš R, Modlinger R, Kašpar J, Majdák A, Blaženec M, Korolyova N, Jirošová A, Schlyter F. Testing the Efficiency of the Push-and-Pull Strategy during Severe Ips typographus Outbreak and Extreme Drought in Norway Spruce Stands. Forests. 2022; 13(12):2175. https://doi.org/10.3390/f13122175
Chicago/Turabian StyleJakuš, Rastislav, Roman Modlinger, Jaroslav Kašpar, Andrej Majdák, Miroslav Blaženec, Nataliya Korolyova, Anna Jirošová, and Fredrik Schlyter. 2022. "Testing the Efficiency of the Push-and-Pull Strategy during Severe Ips typographus Outbreak and Extreme Drought in Norway Spruce Stands" Forests 13, no. 12: 2175. https://doi.org/10.3390/f13122175
APA StyleJakuš, R., Modlinger, R., Kašpar, J., Majdák, A., Blaženec, M., Korolyova, N., Jirošová, A., & Schlyter, F. (2022). Testing the Efficiency of the Push-and-Pull Strategy during Severe Ips typographus Outbreak and Extreme Drought in Norway Spruce Stands. Forests, 13(12), 2175. https://doi.org/10.3390/f13122175