
Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China

Abstract
:1. Introduction
2. Materials and Methods
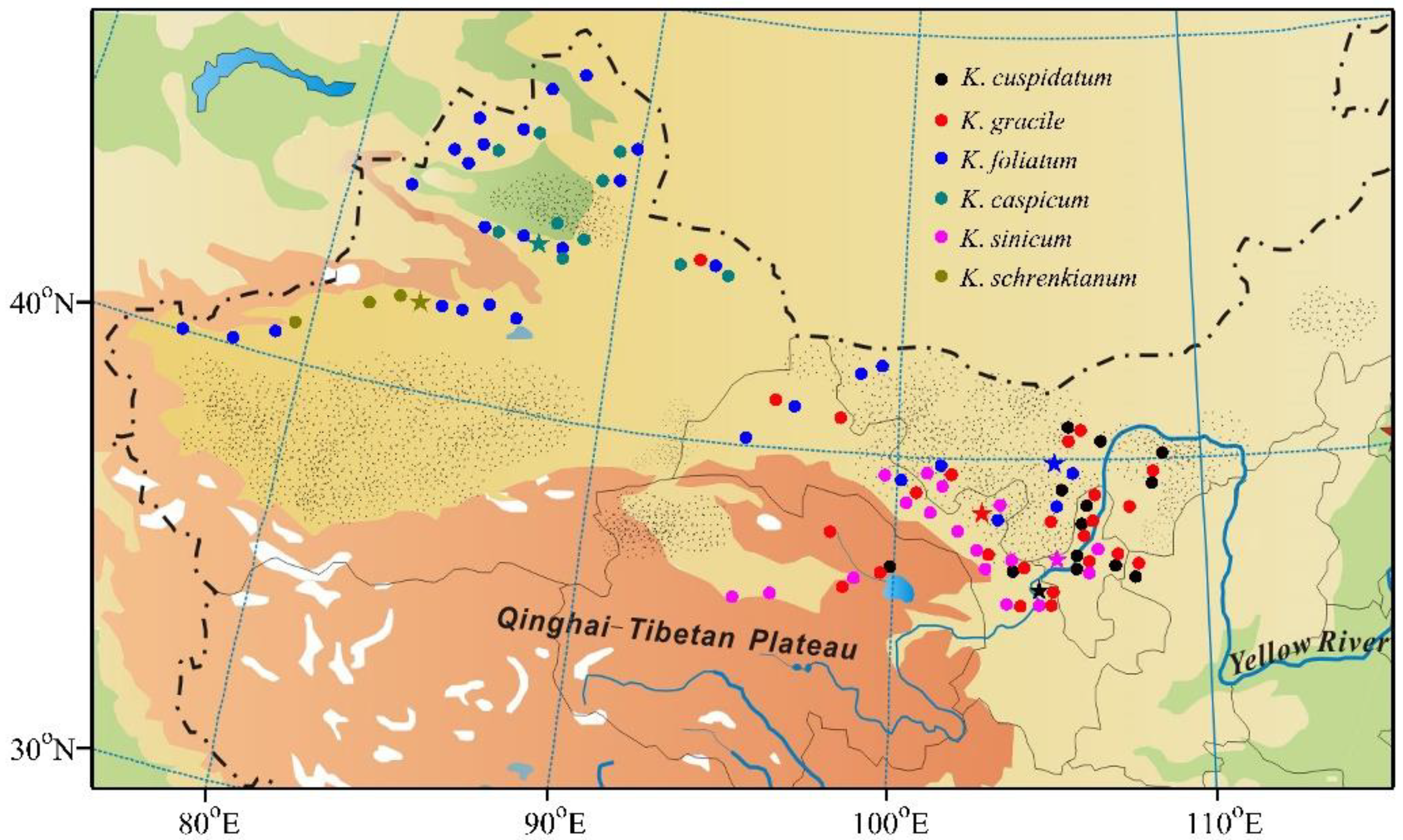
2.1. Field Investigation and Sampling
2.2. Determination of Soil Chemical Properties
2.3. Determination of Seeds’ Germinability
2.4. Data Analysis
3. Results
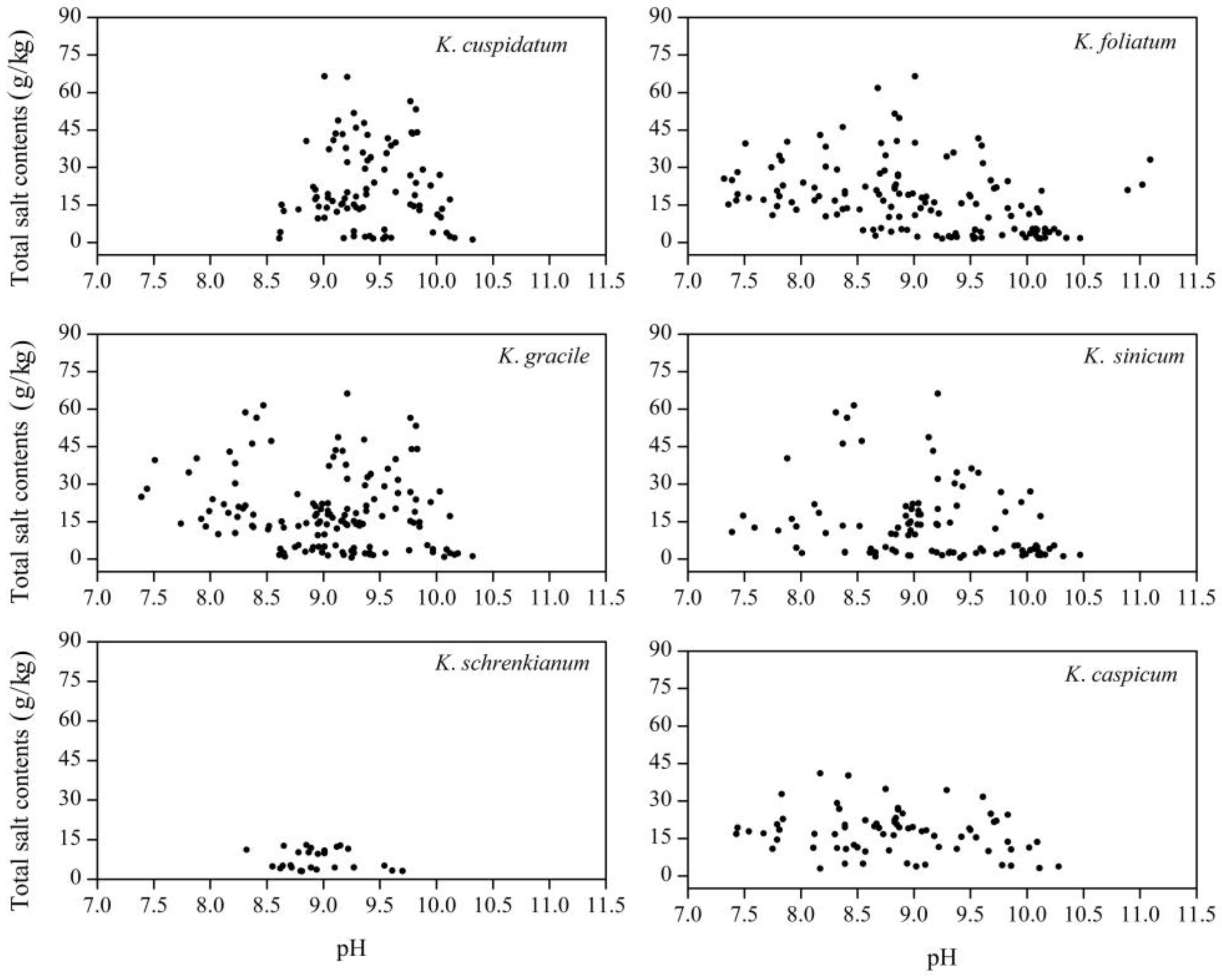
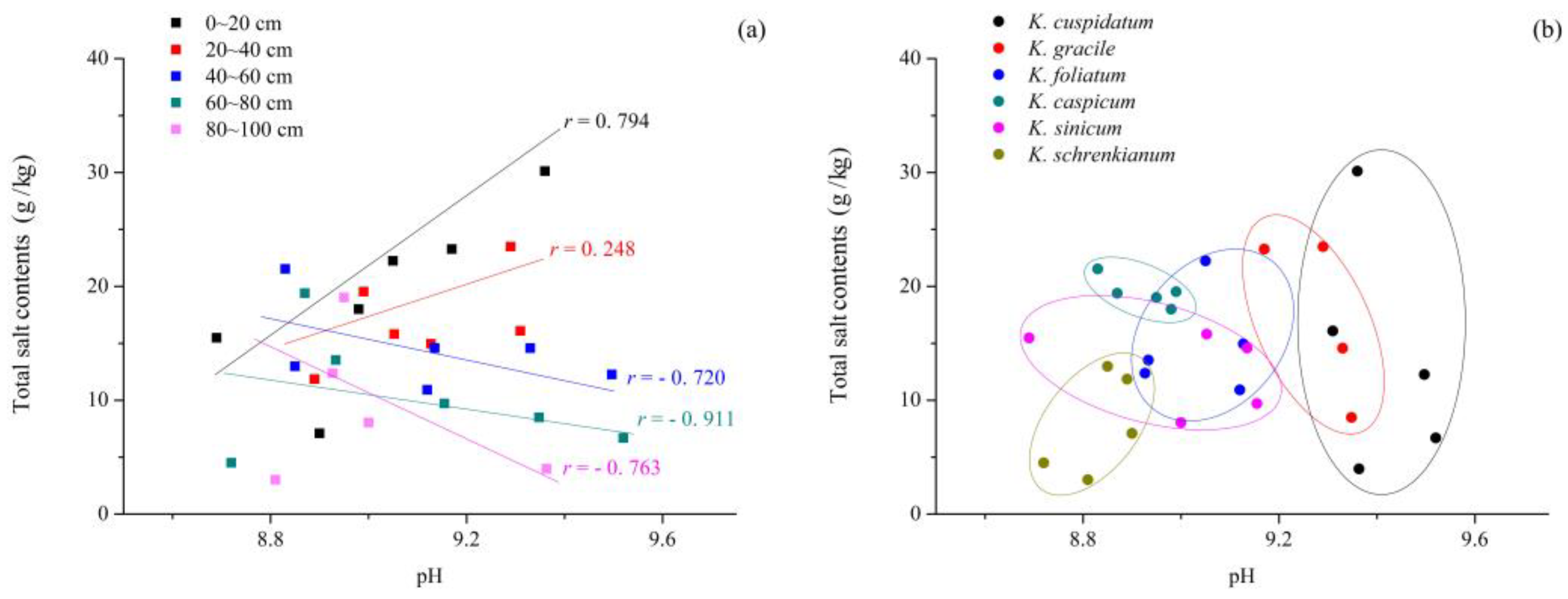
3.1. pH and Total Salt Contents
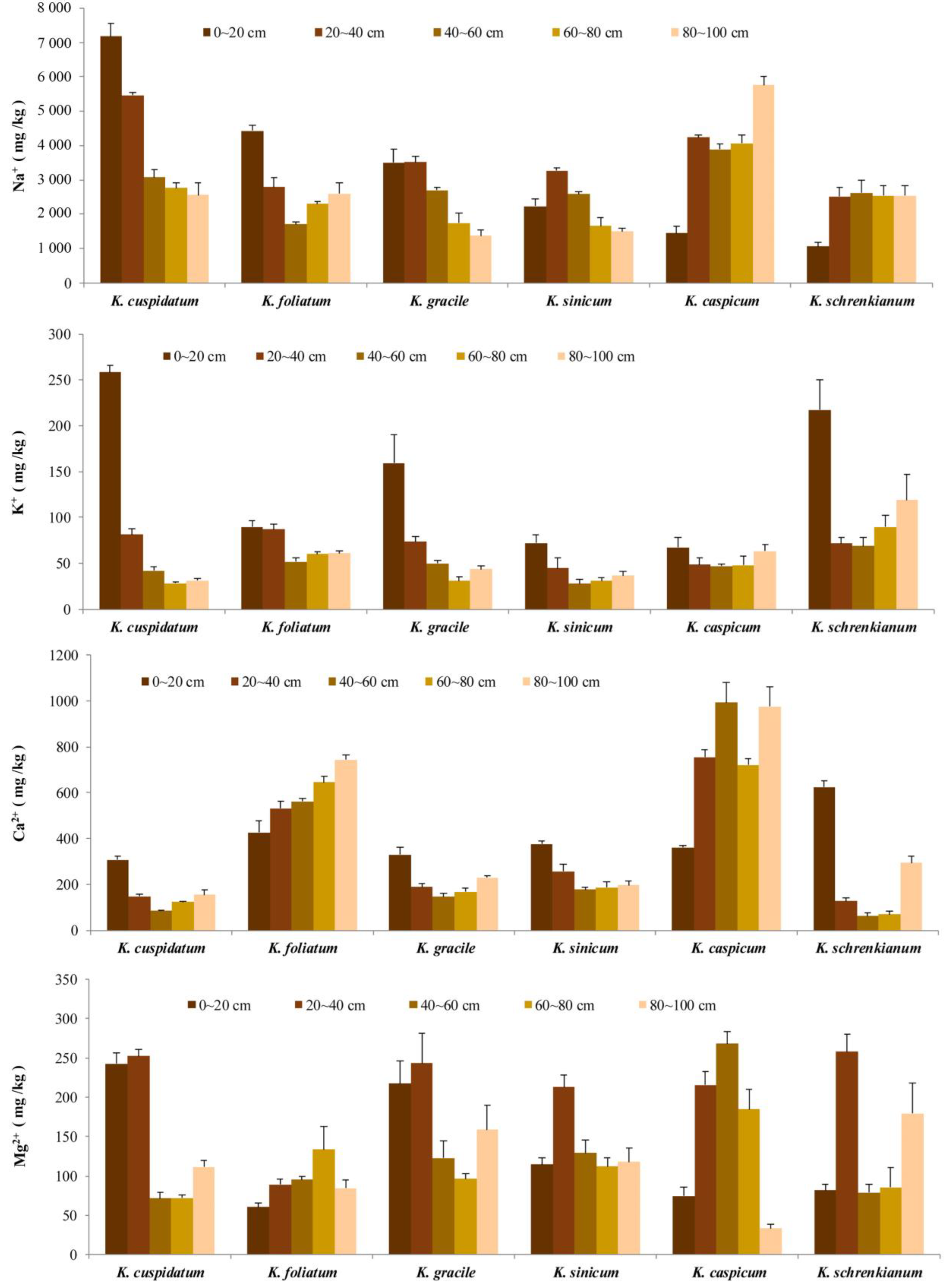
3.2. Na+, Ca2+, K+, and Mg2+ Concentrations
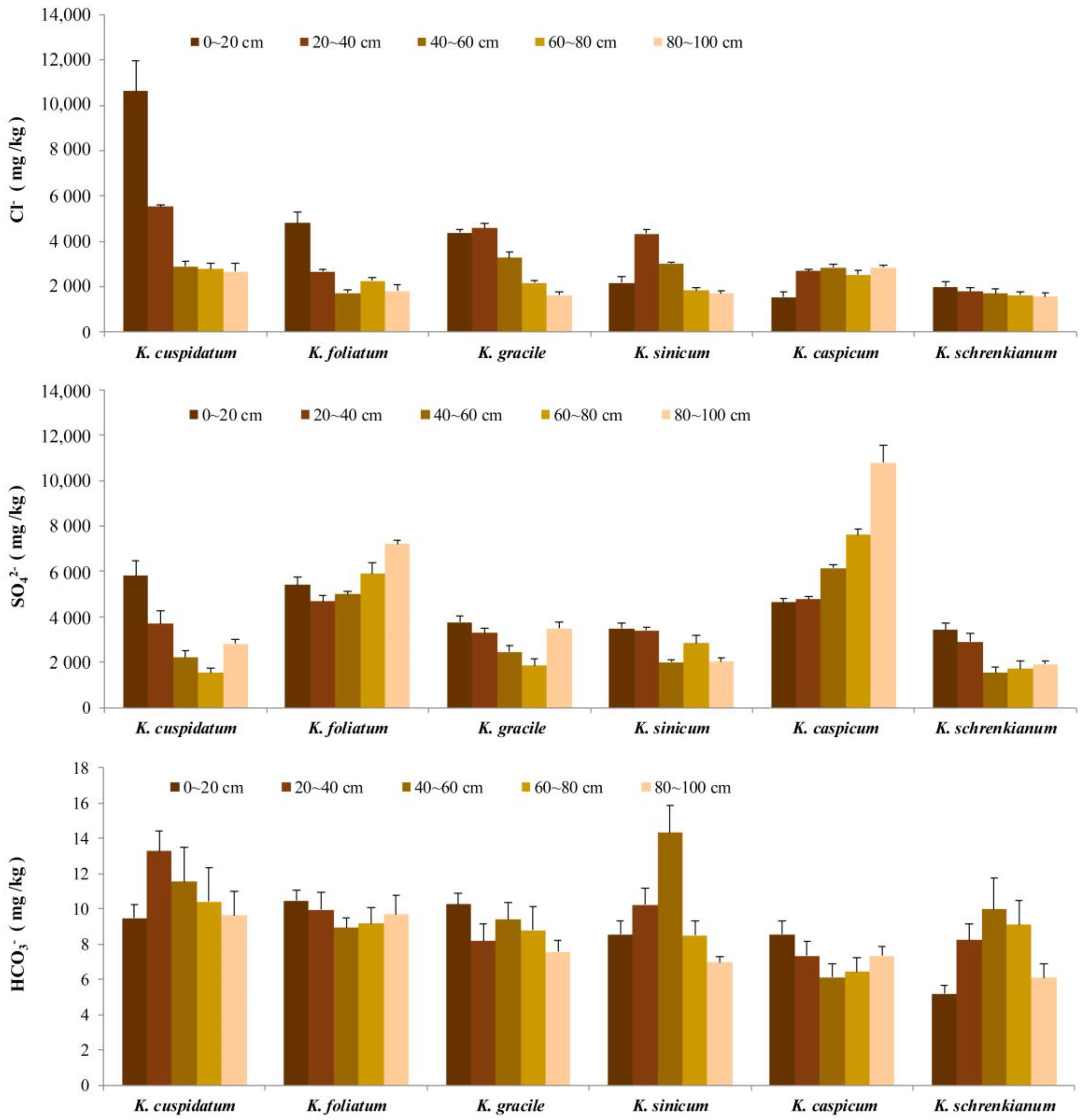
3.3. Cl−, SO42− and HCO3− Concentrations
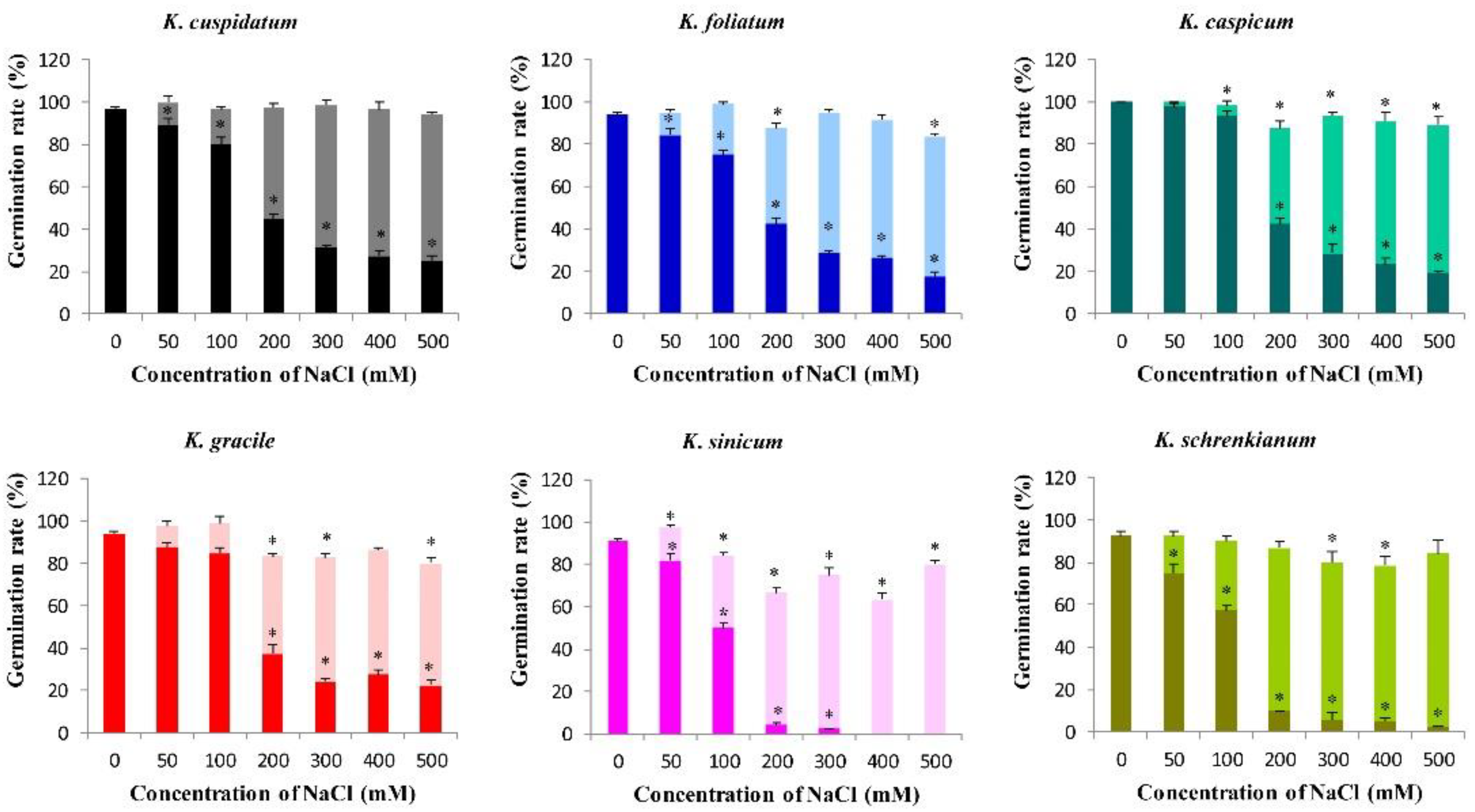
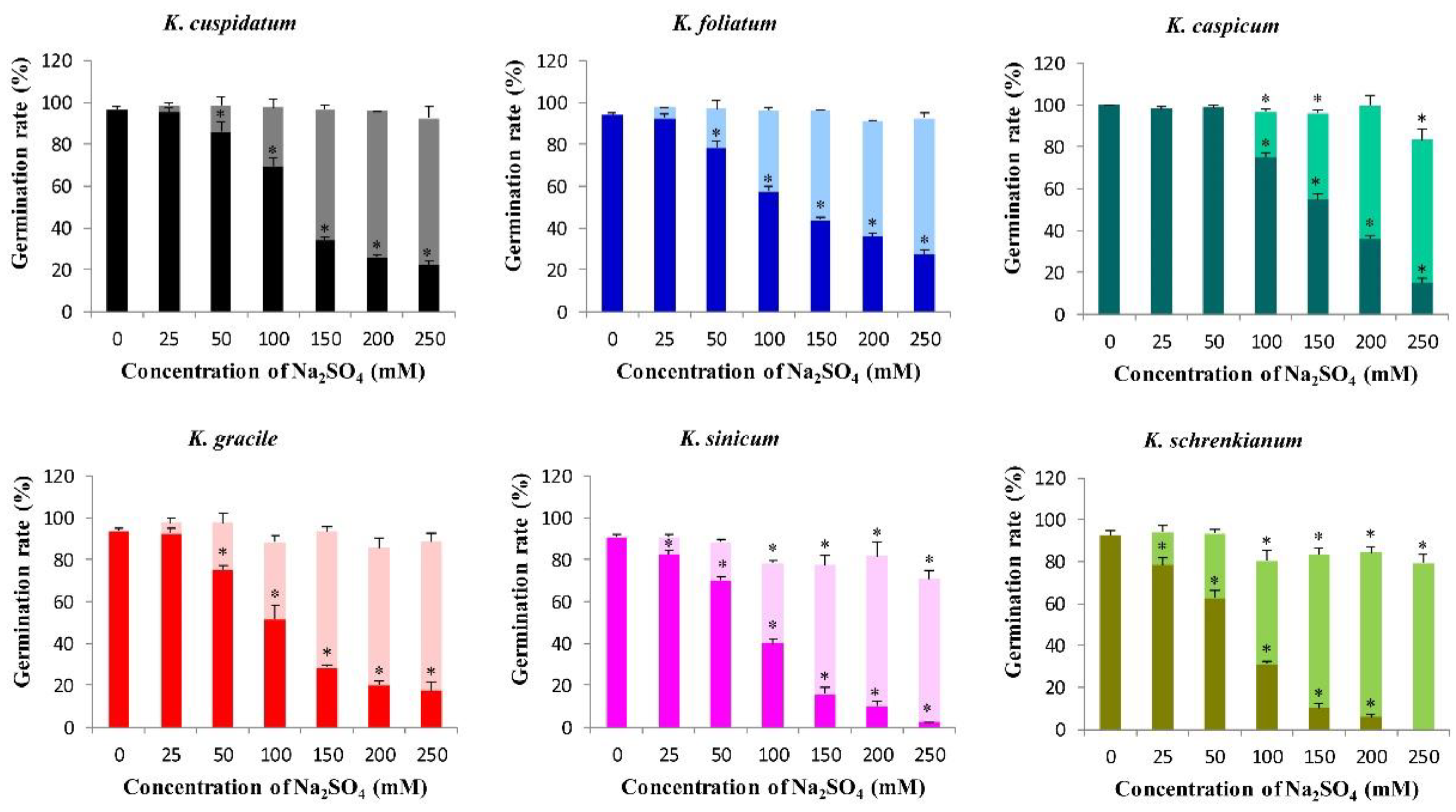
3.4. Germination Rates
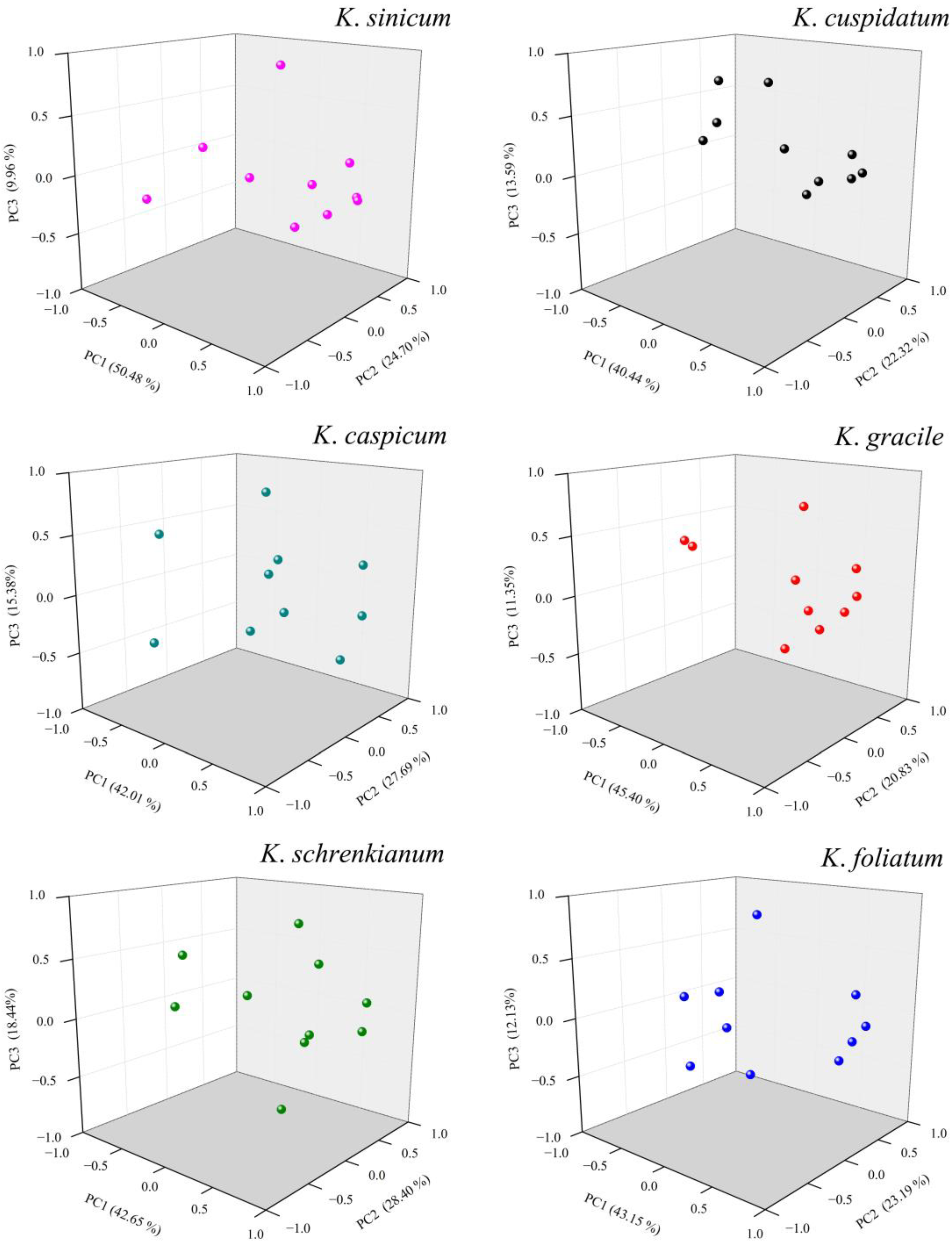
3.5. Principal Component Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheeseman, J.M. The integration of activity in saline environments: Problems and perspectives. Unct. Plant Biol. 2013, 40, 759–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- FAO. FAO Soils Portal. 2016. Available online: http://www.fao.org/soils-portal/soil-management/management-of-some-problem-soils/salt-affected-soils/more-information-on-salt-affected-soils/en (accessed on 9 November 2021).
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Kachout, S.S.; Mansoura, A.B.; Jaffel, K.; Leclerc, J.C.; Rejeb, M.N. The effect of salinity on the growth of the halophyte Atriplex hortensis (Chenopodiaceae). Appl. Ecol. Env. Res. 2009, 7, 319–332. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Waisel, Y. Biology of Halophytes; Academic Press: New York, NY, USA, 1972. [Google Scholar]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Zhao, K.F.; Fan, H.; Jiang, X.Y.; Song, J. Improvement and utilization of saline soil by planting halophytes. Chin. J. Appl. Env. Biol. 2002, 8, 31–35. [Google Scholar]
- Cram, W.J. Negative feedback regulation of transport in cells. The maintenance of turgor, volume and nutrient supply. In Encyclopaedia of Plant Physiology; Luttge, U., Pitman, M.G., Eds.; Springer: Berlin, Germany, 1976; pp. 284–316. [Google Scholar]
- Richau, K.H.; Schat, H. Intraspecific variation of nickel and zinc accumulation and tolerance in the hyperaccumulator Thlaspi caerulescens. Plant and Soil 2009, 314, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.L.; Bourne, E.C.; Von Wettberg, E.J.; Hu, T.T.; Nuzhdin, S.V. Population resequencing reveals local adaptation of Arabidopsis lyrata to serpentine soils. Nat. Genet 2010, 42, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, B.E.; Richards, J.H.; Claassen, V.P.; O’Dell, R.E.; Ferrell, M.A. Species-specific plant-soil interactions influence plant distribution on serpentine soils. Plant and Soil 2011, 342, 327–344. [Google Scholar] [CrossRef] [Green Version]
- Piirainen, M. Kalidium (Chenopodiaceae) the information resource for Euro-Mediterranean plant diversity (Uotila, P., Eds.). Eur. Med. Plantbase 2009, 13, 146. [Google Scholar]
- Liang, X.H.; Wu, Y.X. Identification of Kalidium species (Chenopodiaceae) by DNA barcoding. Sci. Cold. Ari. Regi. 2017, 9, 89–96. [Google Scholar]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Tobe, K.; Li, X.M.; Omasa, K. Seed germination and radicle growth of a halophyte, Kalidium caspicum (Chenopodiaceae). Ann. Bot. 2000, 85, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, G.; Zhang, F.S. Salinity and temperature effects on germination for three salt resistant euhalophytes, Halostachys caspica, Kalidium foliatum and Halocnemum Strobilaceu. Plant and Soil 2006, 279, 201–207. [Google Scholar] [CrossRef]
- Haris, S.; Xia, H.; Elisabeth, B.; Camile, M.; Lindell, B. Predicting species’ tolerance to salinity and alkalinity using distribution data and geochemical modelling: A case study using Australian grasses. Ann. Bot. 2015, 115, 343–351. [Google Scholar]
- Lu, R.K. Analytical Methods for Soil and Agro-Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999. [Google Scholar]
- Li, R.A.; Wang, F.; Qin, F.J.; Lou, F.; Wu, D.Y. Studies on the best curve equation between the total salts and the electrical conductivity of the coastal saline soil. J. Agric. 2015, 5, 59–62. [Google Scholar]
- Wang, W.X.; Li, Z.Z.; Chang, Z.Q. Principal component analysis of geological factors related to landforms-hydrological system of Qilian mountain region. Acta Bot. Boreali-Occident. Sin. 2004, 24, 533–537. (In Chinese) [Google Scholar]
- Tobe, K.; Li, X.M.; Omasa, K. Effects of sodium, magnesium and calcium salts on seed germination and radicle survival of a halophyte, Kalidium caspicum (Chenopodiaceae). Aust. J. Bot. 2002, 50, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.R.; Zhao, R.H.; Du, X.M.; Huang, Z.Y.; Yang, X.J.; Wei, X.Z.; Huang, P.Y. Characteristics of root systems of two halophytes for adaptability to salinity. Sci. Silvae Sin. 2010, 46, 176–182. [Google Scholar]
- Gul, B.; Khan, M.A. Role of calcium in alleviating salinity effects in coastal halophytes. In Ecophysiology of High Salinity Tolerant Plants; Khan, M.A., Weber, D.J., Eds.; Springer: Netherlands, Switzerland, 2006; pp. 107–114. [Google Scholar]
- Very, A.A.; Sentenac, H. Molecular mechanisms and regulation of K+ transport in higher plants. Annu. Rev. Plant Biol. 2003, 54, 575–603. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2007, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- El-Keblawy, A.; Elnaggar, A.; Tammam, A.; Mosa, K.A. Seed provenance affects salt tolerance and germination response of the habitat-indifferent Salsola drummondii halophyte in the arid Arabian deserts. Flora 2020, 266, 151592. [Google Scholar] [CrossRef]
- Attia, H.; Al-Yasi, H.; Alamer, K.; Esmat, F.; Hassan, F.; Elshazly, S.; Hessini, K. Induced anti-oxidation efficiency and others by salt stress in Rosa damascena Miller. Sci. Hortic. 2020, 274, 109681. [Google Scholar] [CrossRef]
- Yasir, T.A.; Khan, A.; Skalicky, M.; Wasaya, A.; Rehmani, M.; Sarwar, N.; Mubeen, K.; Aziz, M.; Hassan, M.M.; Hassan, F.A.S. Exogenous sodium nitroprusside mitigates salt stress in lentil (Lens culinaris medik.) by affecting the growth, yield, and biochemical properties. Molecules 2021, 26, 2576. [Google Scholar] [CrossRef]
- Hassan, F.; Al-Yasi, H.; Ali, E.F.; Alamer, K.; Hessini, K.; Attia, H.; El-Shazly, S. Mitigation of salt stress effects by moringa leaf extract or salicylic acid through motivating antioxidant machinery in damask rose. Can. J. Plant Sci. 2021, 101, 157–165. [Google Scholar] [CrossRef]
- Hassan, F.; Ali, E.; Gaber, A.; Fetouh, M.I.; Mazrou, R. Chitosan nanoparticles effectively combat salinity stress by enhancing antioxidant activity and alkaloid biosynthesis in Catharanthus roseus (L.) G. Don. Plant Physiol. Biochem. 2021, 162, 291–300. [Google Scholar] [CrossRef]
- Bhatt, A.; Gallacher, D.J.; Jarma-Orozco, A.; Fernandes, D.; Pompelli, M.F. Seed provenance selection of wild halophyte seeds improves coastal rehabilitation efficiency. Estuar. Coast. Shelf Sci. 2021, 265, 107657. [Google Scholar] [CrossRef]
- Vicente, O.; Boscaiu, M.; Naranjo, M.O.; Estrelles, E.; Bellés, J.M.; Soriano, P. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 2004, 58, 463–481. [Google Scholar] [CrossRef]
- Cronin, J.K.; Bundock, P.C.; Henry, R.J.; Nevo, E. Adaptive climatic molecular evolution in wild barley at the Isa defense locus. PNAS 2007, 104, 2773–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcazar, R.; Pecinka, A.; Aarts, M.G.; Fransz, P.F.; Koornneef, M. Signals of speciation within Arabidopsis thaliana in comparison with its relatives. Curr. Opin. Plant Biol. 2012, 15, 205–211. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth | Ca2+ | Mg2+ | K+ | Na+ | HCO3− | Cl− | SO42− |
|---|---|---|---|---|---|---|---|
| 0~20 cm | −0.841 * | 0.725 | 0.018 | 0.896 * | 0.992 ** | 0.856 * | 0.793 |
| 20~40cm | −0.093 | −0.156 | −0.680 | 0.632 | −0.969 ** | 0.689 | 0.031 |
| 40~60 cm | 0.677 | 0.980 ** | −0.140 | 0.986 ** | −0.809 | 0.880 * | 0.784 |
| 60~80 cm | 0.934 * | 0.991 ** | 0.567 | 0.972 ** | −0.873 | 0.933 * | 0.966 ** |
| 80~100 cm | 0.965 ** | −0.958 * | 0.660 | 0.973 ** | −0.502 | 0.167 | 0.344 |
| Species | Main Factors and Loadings (Correlation Coefficients) | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| K. schrenkianum | bio5 (0.971) | pH (0.797) | TS (0.719) |
| K. sinicum | bio9 (0.975) | bio14 (0.812) | TS (0.906) |
| K.cuspidatum | bio5 (0.892) | bio14 (0.707) | TS (0.766) |
| K. capsicum | bio1 (0.949) | bio14 (0.870) | pH (0.833) |
| K. gracile | bio1 (0.993) | bio12 (0.832) | bio14 (0.658) |
| K. foliatum | bio9 (0.924) | bio5 (0.713) | TS (0.840) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Chang, Z.; Liang, X.; Wu, Y. Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China. Forests 2022, 13, 2178. https://doi.org/10.3390/f13122178
Liu D, Chang Z, Liang X, Wu Y. Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China. Forests. 2022; 13(12):2178. https://doi.org/10.3390/f13122178
Chicago/Turabian StyleLiu, Decheng, Zongqiang Chang, Xiaohui Liang, and Yuxia Wu. 2022. "Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China" Forests 13, no. 12: 2178. https://doi.org/10.3390/f13122178
APA StyleLiu, D., Chang, Z., Liang, X., & Wu, Y. (2022). Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China. Forests, 13(12), 2178. https://doi.org/10.3390/f13122178